
Biosynthesis of Silver Chloride Nanoparticles by Rhizospheric Bacteria and Their Antibacterial Activity against Phytopathogenic Bacterium Ralstonia solanacearum




Abstract
:1. Introduction
2. Results
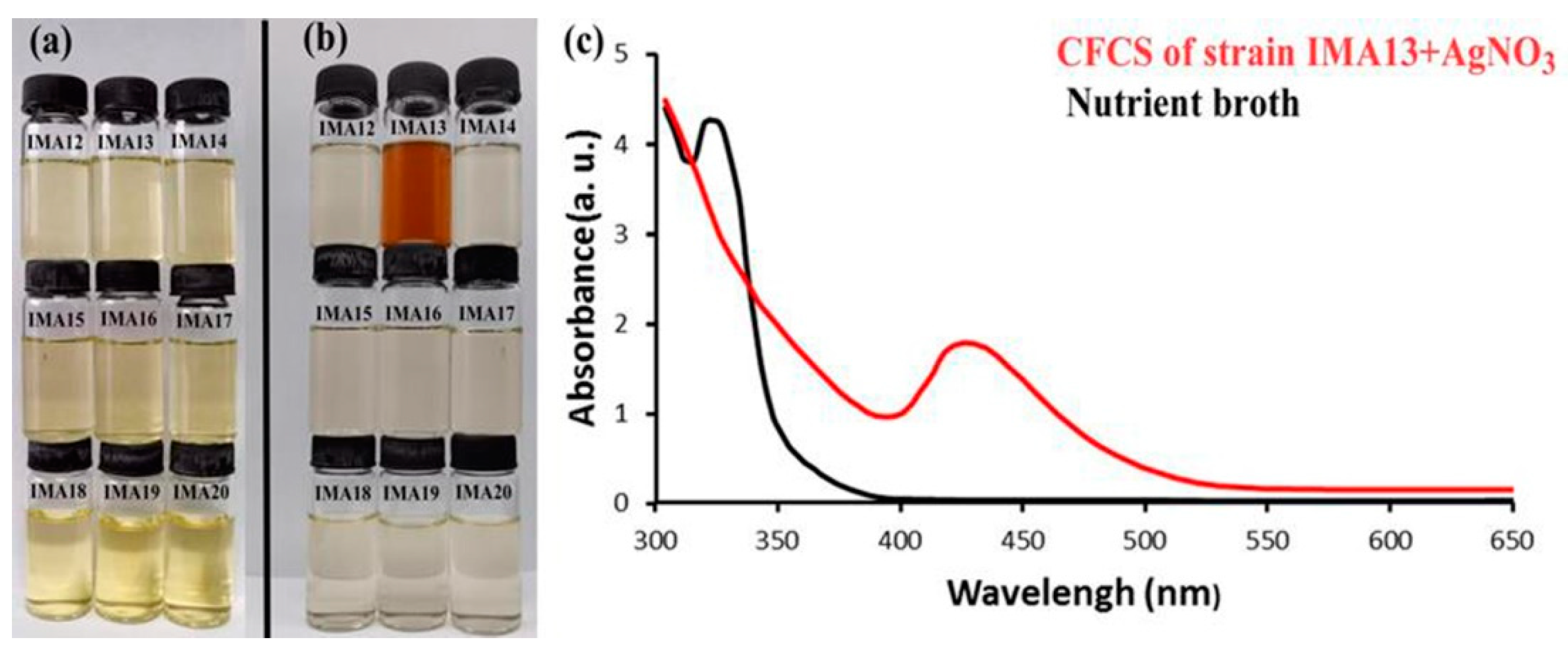
2.1. Bacteria Strains and Biosynthesis of AgCl-NPs
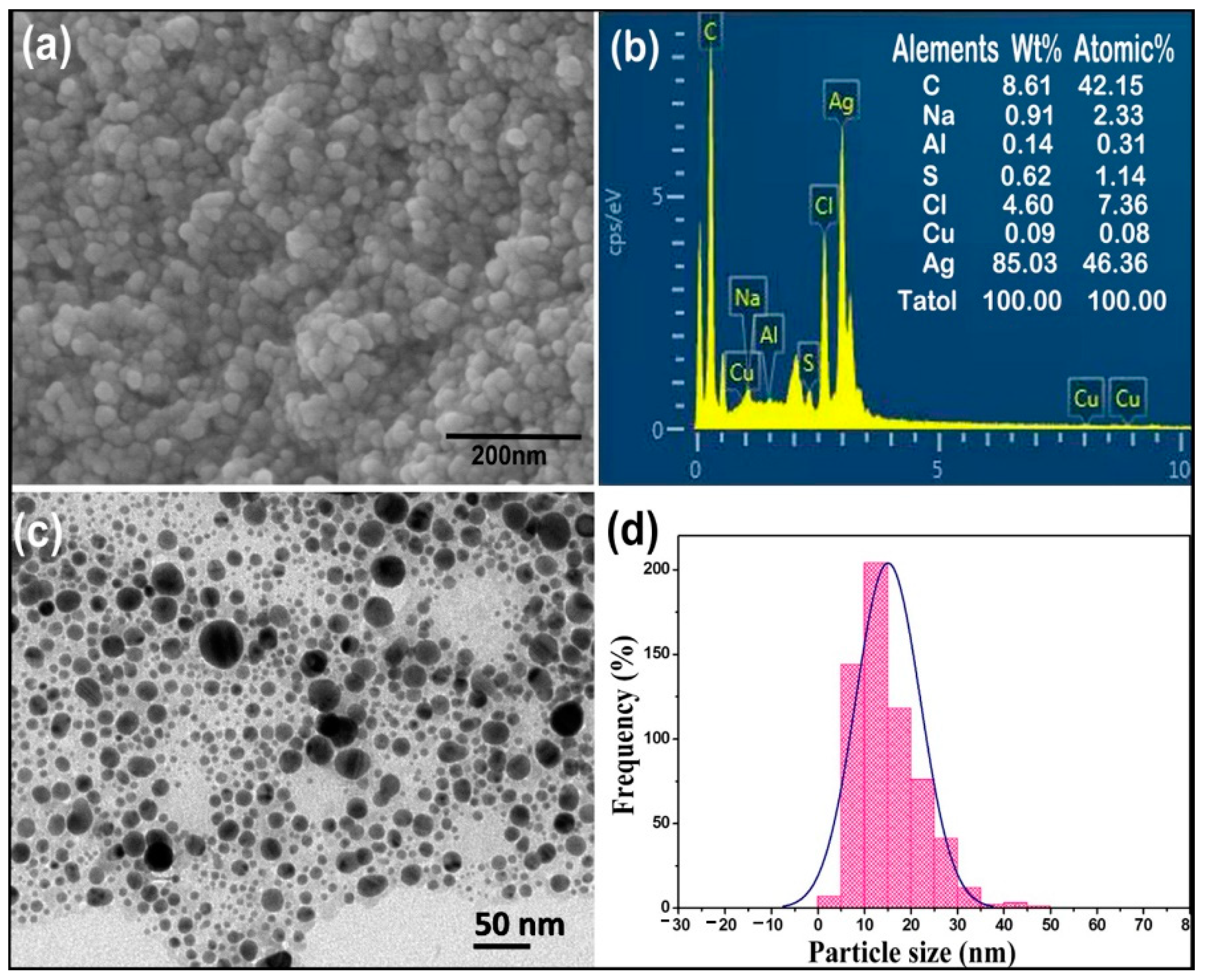
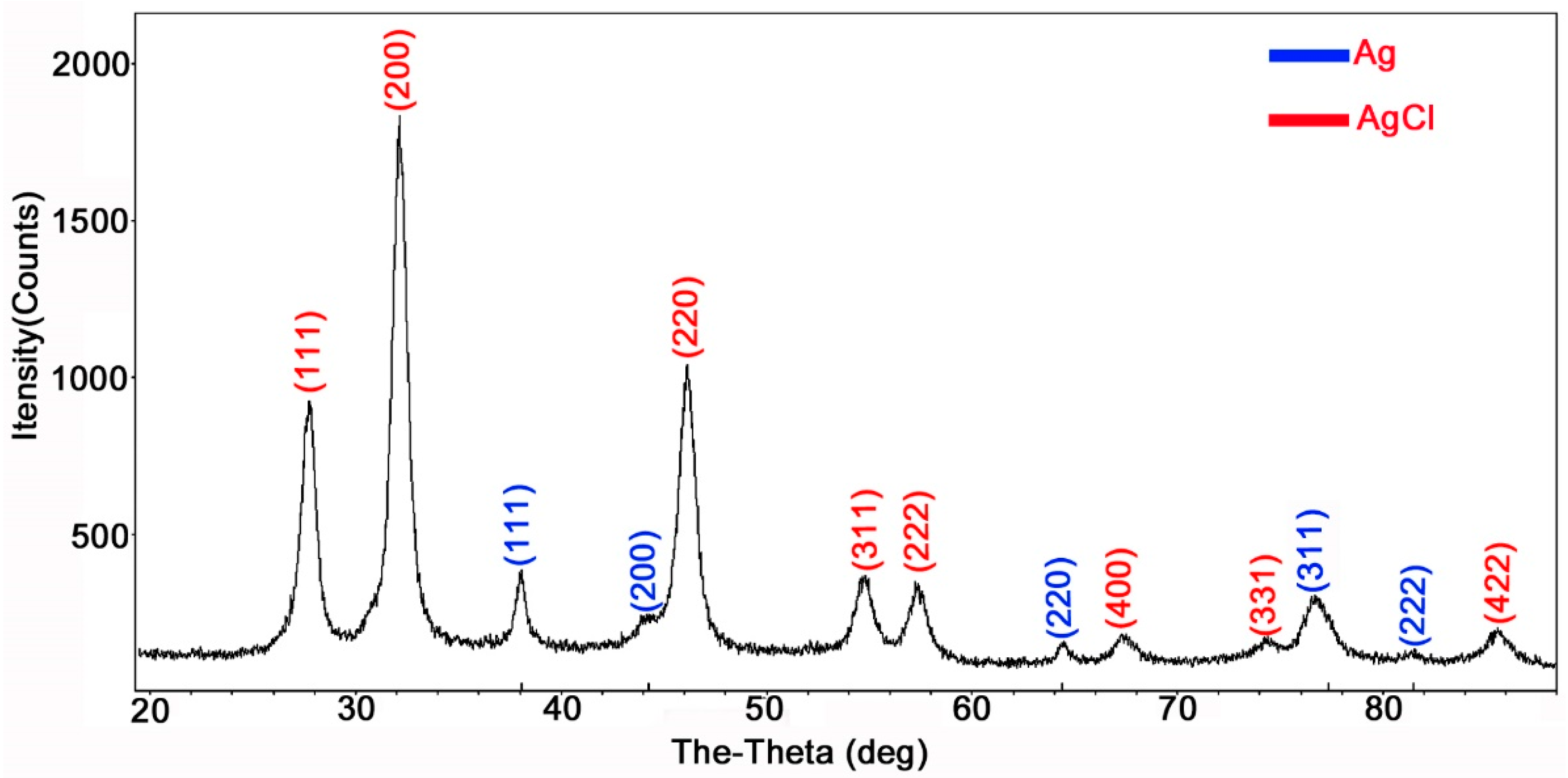
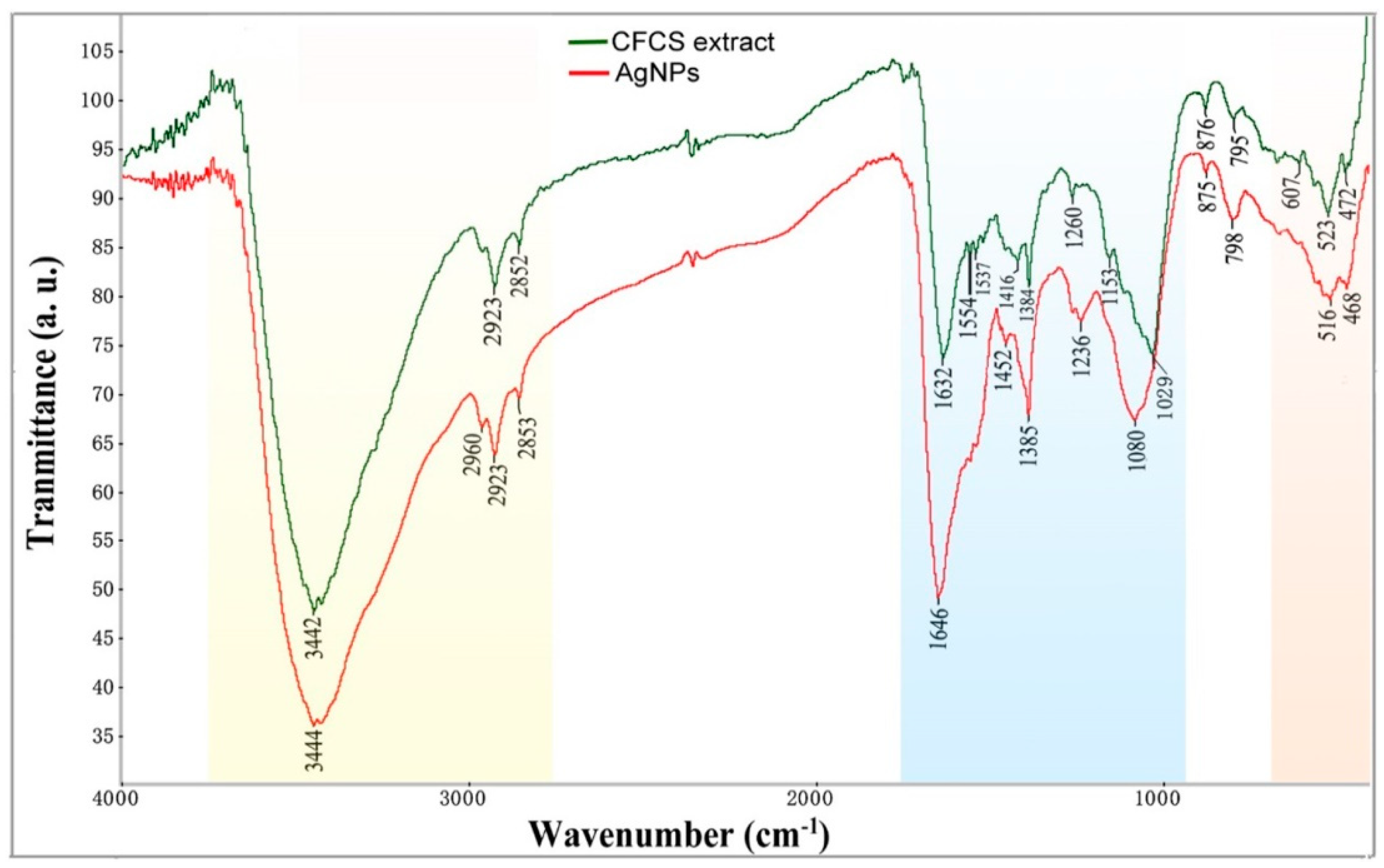
2.2. Characterizations of AgCl-NPs
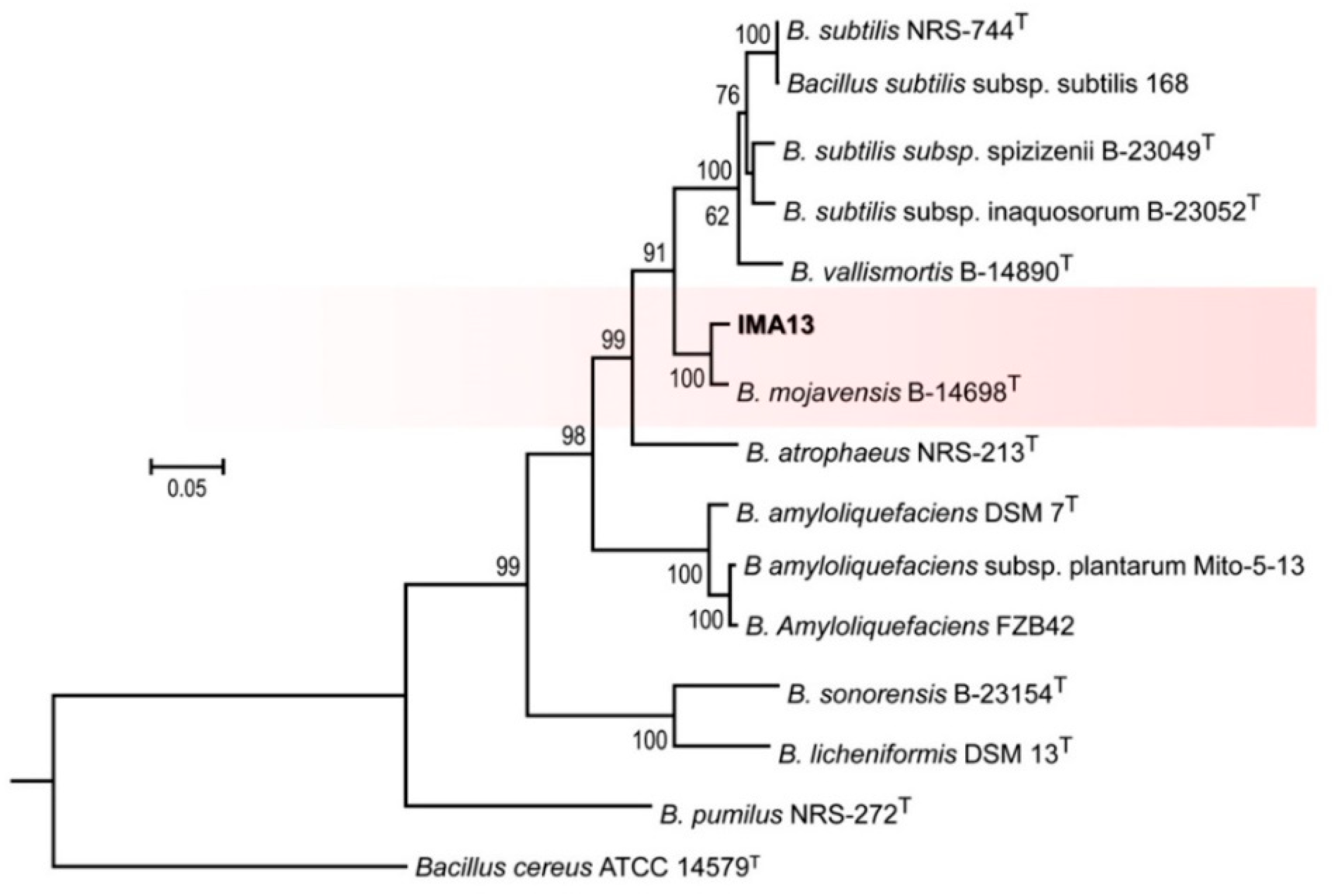
2.3. Identification of Strain
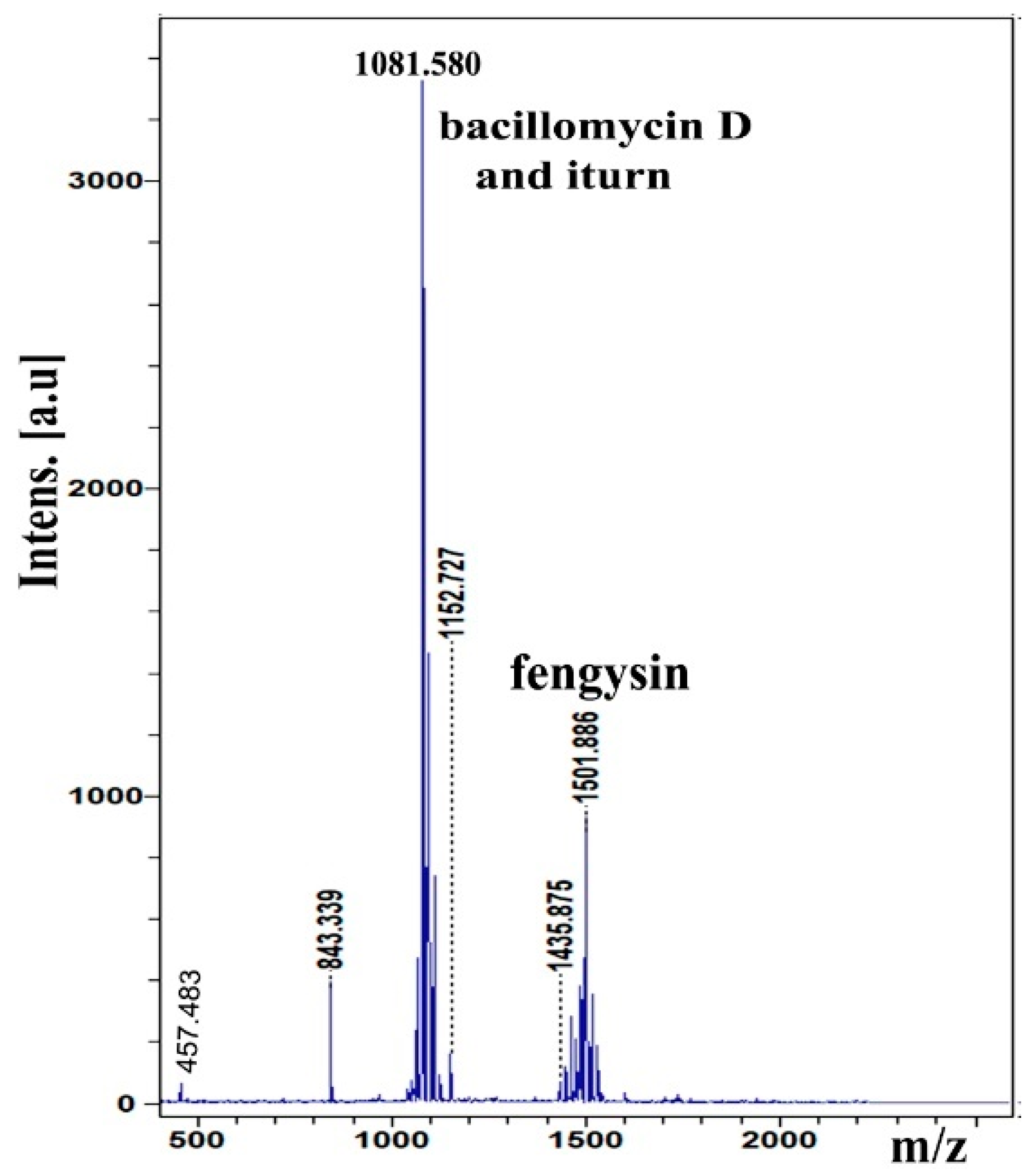
2.4. Analysis of Lipopeptide Compound
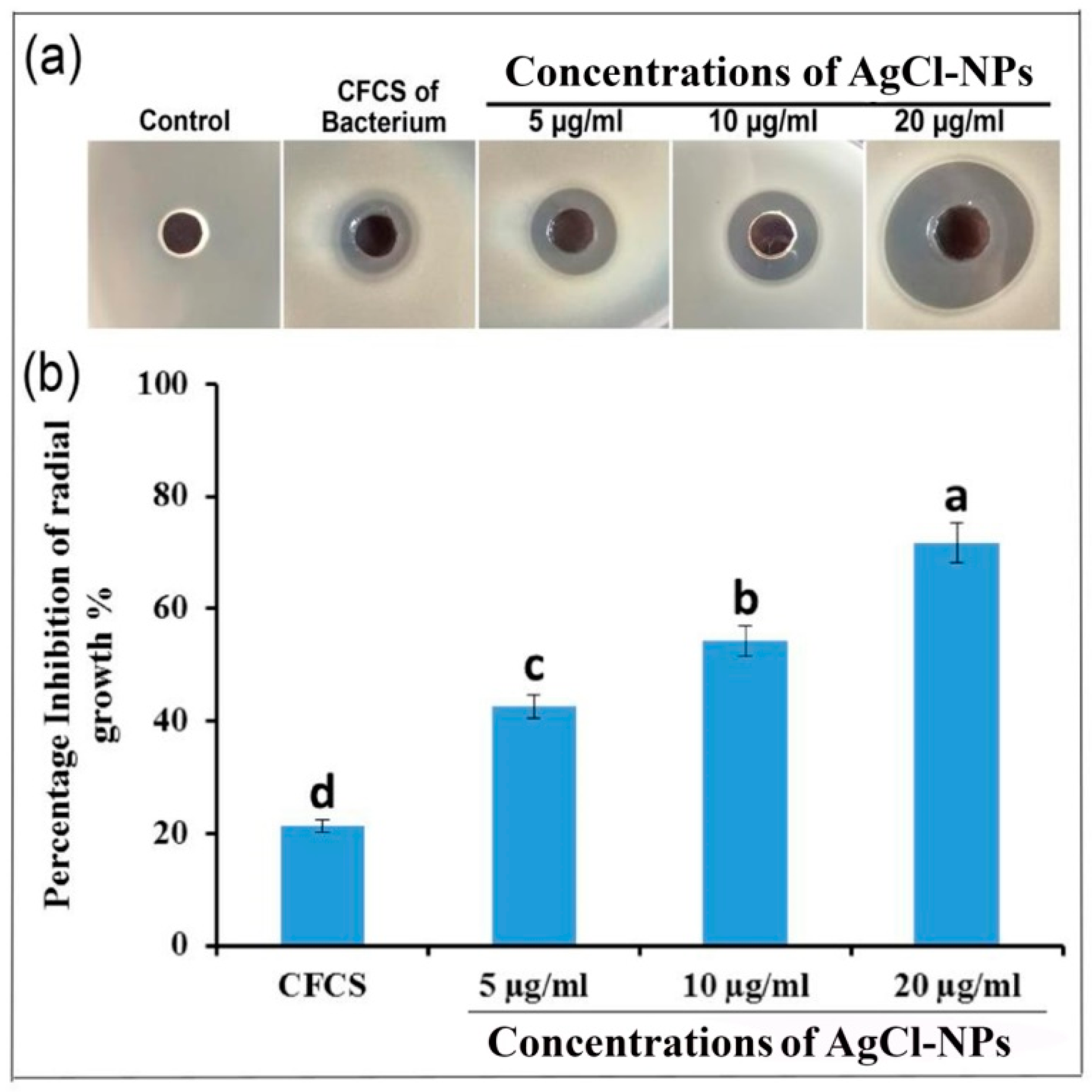
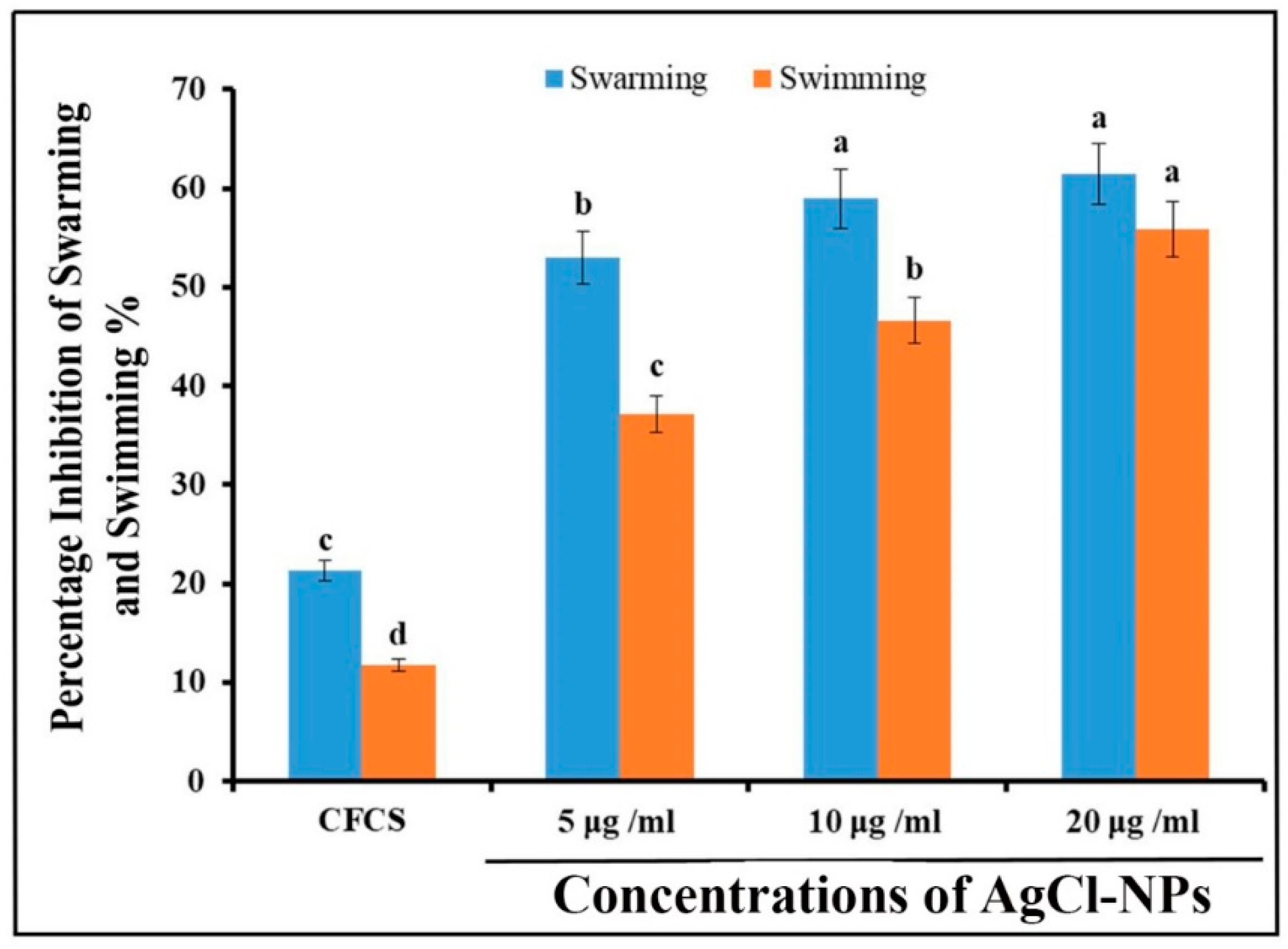
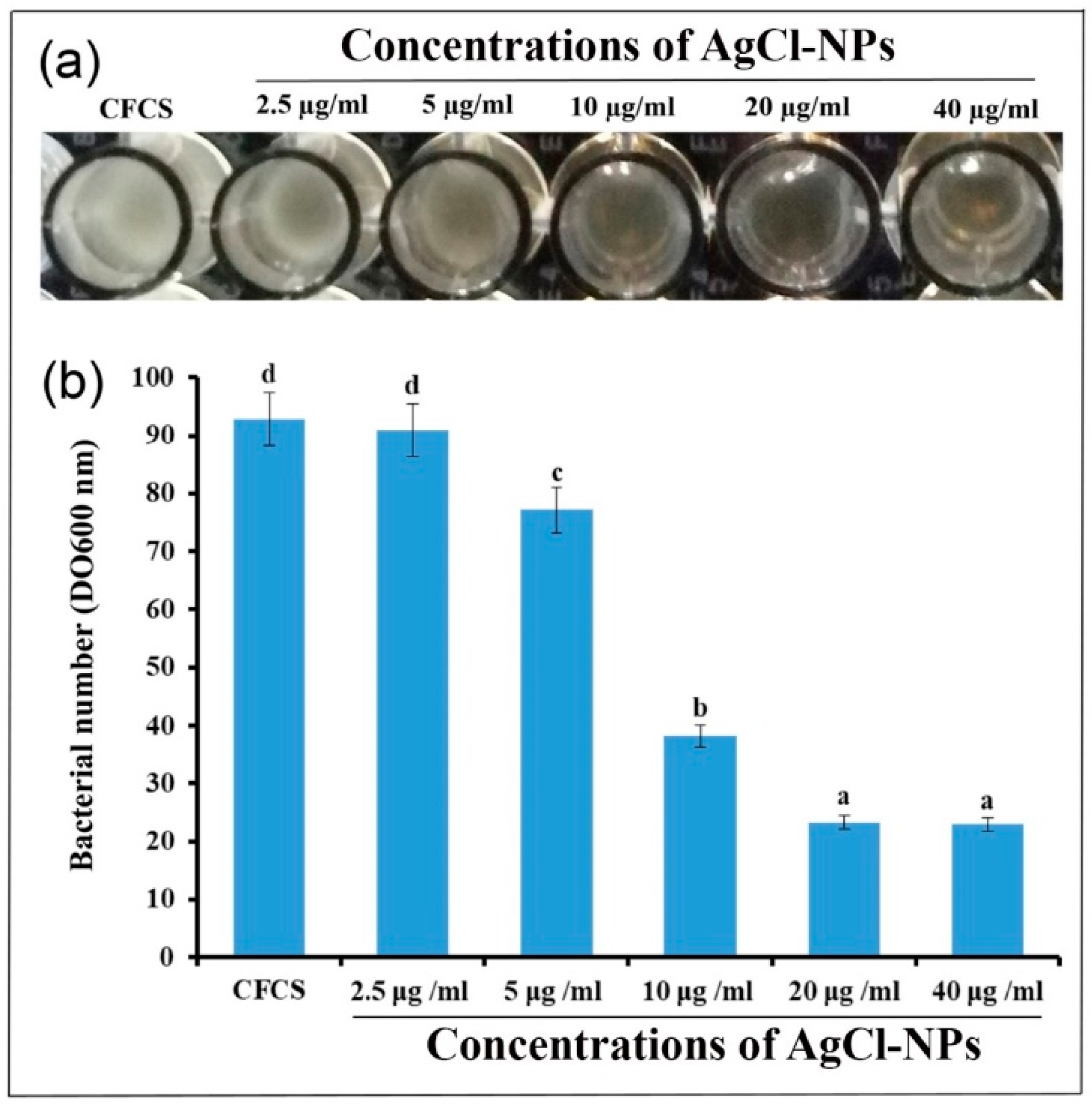
2.5. Antibacterial Activity of AgCl-NPs
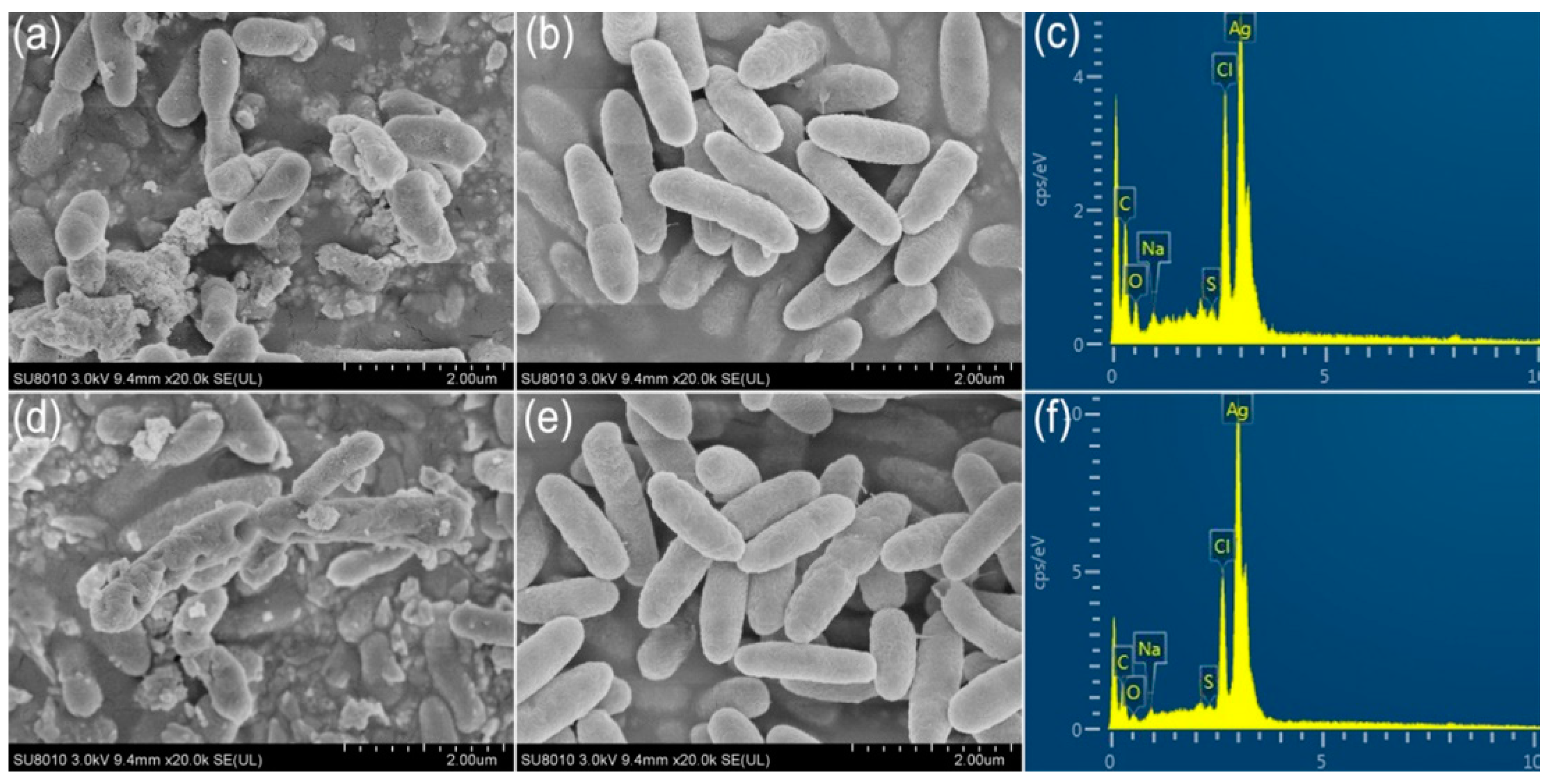
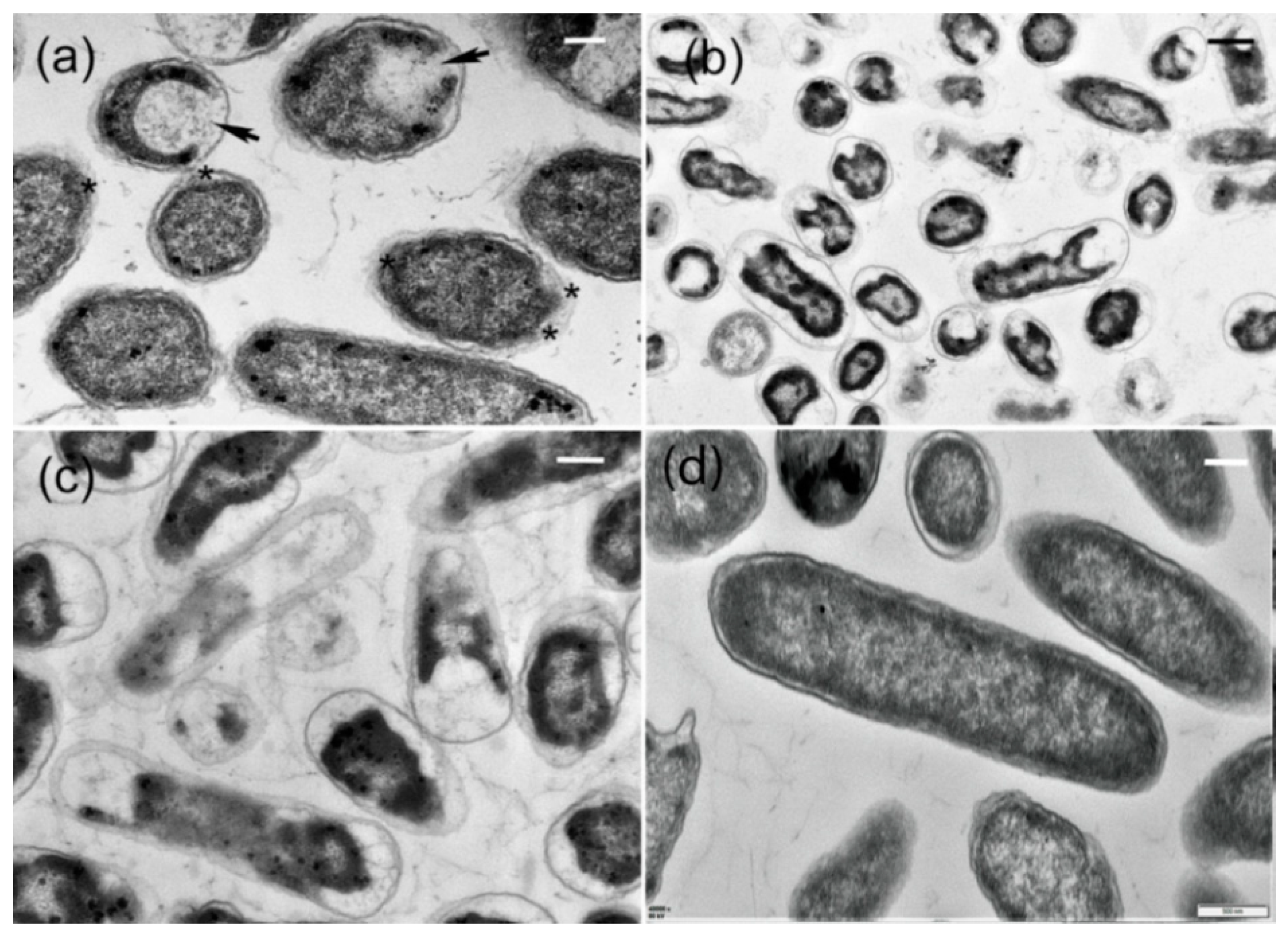
2.6. Ultrastructural Characteristics of AgCl-NPs Interaction with Pathogen
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Bacterial Isolation
4.2. Biosynthesis of AgCl-NPs
4.3. Characterizations of AgCl-NPs
4.4. Identification of Bacterium Strain IMA13
4.5. Phylogenetic Analysis
4.6. Lipopeptide Identification
4.7. Antibacterial Activity of AgCl-NPs
4.7.1. Agar Well Diffusion Assay
4.7.2. Motility Assay
4.7.3. Determination of Minimum Inhibitory Concentration (MIC)
4.8. Ultrastructural Characteristics of the AgCl-NP Interaction with Pathogen
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Peeters, N.; Guidot, A.; Vailleau, F.; Valls, M. Ralstonia solanacearum, a widespread bacterial plant pathogen in the post-genomic era. Mol. Plant Pathol. 2013, 14, 651–662. [Google Scholar]
- Elphinstone, J.G.; Allen, C.; Prior, P.; Hayward, A.C. The current bacterial wilt situation: A global overview. In Bacterial Wilt the Disease & the Ralstonia Solanacearum Species Complex; APS Press: Saint Paul, MN, USA, 2005; pp. 9–28. [Google Scholar]
- Genin, S. Molecular traits controlling host range and adaptation to plants in Ralstonia solanacearum. New Phytol. 2010, 187, 920–928. [Google Scholar] [CrossRef]
- Hayward, A.C. Biology and Epidemiology of Bacterial Wilt Caused by Pseudomonas Solanacearum. Annu. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef]
- Wicker, E.; Grassart, L.; Coranson-Beaudu, R.; Mian, D.; Guilbaud, C.; Fegan, M.; Prior, P. Ralstonia solanacearum Strains from Martinique (French West Indies) Exhibiting a New Pathogenic Potential. Appl. Environ. Microbiol. 2007, 73, 6790–6801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, B.; López, M.M.; Biosca, E.G. Survival strategies and pathogenicity of Ralstonia solanacearum phylotype II subjected to prolonged starvation in environmental water microcosms. Microbiology 2008, 154, 3590–3598. [Google Scholar] [CrossRef] [Green Version]
- Corral, J.; Sebastià, P.; Coll, N.S.; Barbé, J.; Aranda, J.; Valls, M. Twitching and swimming motility play a role in Ralstonia solanacearum pathogenicity. Msphere 2020, 5, e00740-19. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denny, T. Plant pathogenic Ralstonia species. In Plant-Associated Bacteria; Springer: Dordrecht, The Netherlands, 2007; pp. 573–644. [Google Scholar]
- Tomah, A.A.; Alamer, I.S.A.; Li, B.; Zhang, J.-Z. Mycosynthesis of Silver Nanoparticles Using Screened Trichoderma Isolates and Their Antifungal Activity against Sclerotinia sclerotiorum. Nanomaterials 2020, 10, 1955. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, K.B.; Sakthivel, N. Biological synthesis of metal nanoparticles by microbes. Adv. Colloid Interface Sci. 2010, 156, 1–13. [Google Scholar] [CrossRef]
- Schabes-Retchkiman, P.; Canizal, G.; Herrera-Becerra, R.; Zorrilla, C.; Liu, H.; Ascencio, J. Biosynthesis and characterization of Ti/Ni bimetallic nanoparticles. Opt. Mater. 2006, 29, 95–99. [Google Scholar] [CrossRef]
- Singh, P.; Kim, Y.J.; Singh, H.; Wang, C.; Hwang, K.H.; Farh, M.E.A.; Yang, D.C. Biosynthesis, characterization, and antimicrobial applications of silver nanoparticles. Int. J. Nanomed. 2015, 10, 2567–2577. [Google Scholar]
- Velusamy, P.; Kumar, G.V.; Jeyanthi, V.; Das, J.; Pachaiappan, R. Bio-Inspired Green Nanoparticles: Synthesis, Mechanism, and Antibacterial Application. Toxicol. Res. 2016, 32, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Elbeshehy, E.K.F.; Elazzazy, A.M.; Aggelis, G. Silver nanoparticles synthesis mediated by new isolates of Bacillus spp., nanoparticle characterization and their activity against Bean Yellow Mosaic Virus and human pathogens. Front. Microbiol. 2015, 6, 453. [Google Scholar] [CrossRef] [Green Version]
- Jalal, M.; Ansari, M.A.; Alzohairy, M.A.; Ali, S.G.; Khan, H.M.; Almatroudi, A.; Siddiqui, M.I. Anticandidal activity of biosynthesized silver nanoparticles: Effect on growth, cell morphology, and key virulence attributes of Candida species. Int. J. Nanomed. 2019, 14, 4667–4679. [Google Scholar] [CrossRef] [Green Version]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic Basis of Antimicrobial Actions of Silver Nanoparticles. Front. Microbiol. 2016, 7, 1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, N.; Bhargava, A.; Majumdar, S.; Tarafdar, J.C.; Panwar, J. Extracellular biosynthesis and characterization of silver nanoparticles using Aspergillus flavusNJP08: A mechanism perspective. Nanoscale 2010, 3, 635–641. [Google Scholar] [CrossRef]
- Shameli, K.; Bin Ahmad, M.; Jazayeri, S.D.; Sedaghat, S.; Shabanzadeh, P.; Jahangirian, H.; Mahdavi, M.; Abdollahi, Y. Synthesis and Characterization of Polyethylene Glycol Mediated Silver Nanoparticles by the Green Method. Int. J. Mol. Sci. 2012, 13, 6639–6650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooney, A.P.; Price, N.P.J.; Ehrhardt, C.; Swezey, J.L.; Bannan, J.D. Phylogeny and molecular taxonomy of the Bacillus subtilis species complex and description of Bacillus subtilis subsp. inaquosorum subsp. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 2429–2436. [Google Scholar] [CrossRef] [PubMed]
- Soussi, S.; Essid, R.; Hardouin, J.; Gharbi, D.; Elkahoui, S.; Tabbene, O.; Cosette, P.; Jouenne, T.; Limam, F. Utilization of Grape Seed Flour for Antimicrobial Lipopeptide Production by Bacillus amyloliquefaciens C5 Strain. Appl. Biochem. Biotechnol. 2018, 187, 1460–1474. [Google Scholar] [CrossRef]
- Pathak, K.; Keharia, H. Characterization of fungal antagonistic bacilli isolated from aerial roots of banyan (Ficus benghalensis) using intact-cell MALDI-TOF mass spectrometry (ICMS). J. Appl. Microbiol. 2013, 114, 1300–1310. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Zheng, Q.-W.; Wei, T.; Zhang, Z.-Q.; Zhao, C.-F.; Zhong, H.; Xu, Q.-Y.; Lin, J.-F.; Guo, L.-Q. Isolation and Characterization of Fengycins Produced by Bacillus amyloliquefaciens JFL21 and Its Broad-Spectrum Antimicrobial Potential against Multidrug-Resistant Foodborne Pathogens. Front. Microbiol. 2020, 11, 3319. [Google Scholar] [CrossRef]
- Vater, J.; Kablitz, B.; Wilde, C.; Franke, P.; Mehta, N.; Cameotra, S.S. Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry of Lipopeptide Biosurfactants in Whole Cells and Culture Filtrates of Bacillus subtilis C-1 Isolated from Petroleum Sludge. Appl. Environ. Microbiol. 2002, 68, 6210–6219. [Google Scholar] [CrossRef] [Green Version]
- Hanif, A.; Zhang, F.; Li, P.; Li, C.; Xu, Y.; Zubair, M.; Zhang, M.; Jia, D.; Zhao, X.; Liang, J.; et al. Fengycin Produced by Bacillus amyloliquefaciens FZB42 Inhibits Fusarium graminearum Growth and Mycotoxins Biosynthesis. Toxins 2019, 11, 295. [Google Scholar] [CrossRef] [Green Version]
- Saklani, V.; Suman, J.V.K.; Jain, K. Microbial synthesis of silver nanoparticles: A review. J. Biotechnol. Biomater. 2012, 13, 2718–2725. [Google Scholar] [CrossRef]
- Al-Shabib, N.A.; Husain, F.M.; Nadeem, M.; Khan, M.S.; Al-Qurainy, F.; Alyousef, A.A.; Arshad, M.; Khan, A.; Khan, J.M.; Alam, P.; et al. Bio-inspired facile fabrication of silver nanoparticles from in vitro grown shoots of Tamarix nilotica: Explication of its potential in impeding growth and biofilms of Listeria monocytogenes and assessment of wound healing ability. RSC Adv. 2020, 10, 30139–30149. [Google Scholar] [CrossRef]
- Spagnoletti, F.N.; Spedalieri, C.; Kronberg, M.F.; Giacometti, R. Extracellular biosynthesis of bactericidal Ag/AgCl nanoparticles for crop protection using the fungus Macrophomina phaseolina. J. Environ. Manag. 2018, 231, 457–466. [Google Scholar] [CrossRef]
- Alamri, S.A.; Hashem, M.; Nafady, N.A.; Sayed, M.A.; Alshehri, A.M.; Alshaboury, G.A. Controllable biogenic synthesis of intracellular silver/silver chloride nanoparticles by Meyerozyma guilliermondii KX008616. J. Microbiol. Biotechnol. 2018, 28, 917–930. [Google Scholar] [CrossRef] [Green Version]
- Ghiuta, I.; Croitoru, C.; Kost, J.; Wenkert, R.; Munteanu, D. Bacteria-Mediated Synthesis of Silver and Silver Chloride Nanoparticles and Their Antimicrobial Activity. Appl. Sci. 2021, 11, 3134. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Pohl, P. Synthesis of biogenic silver nanoparticles (Agcl-NPs) using a pulicaria vulgaris gaertn. aerial part extract and their application as antibacterial, antifungal and antioxidant agents. Nanomaterials 2020, 10, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado-Muñiz, M.; Luna, C.; Mendoza-Reséndez, R.; Barriga-Castro, E.D.; Soto-Rodriguez, S.; Ricque-Marie, D.; Cruz-Suarez, L.E. Silver nanoparticles against acute hepatopancreatic necrosis disease (AHPND) in shrimp and their depuration kinetics. Environ. Boil. Fishes 2019, 32, 2431–2445. [Google Scholar] [CrossRef]
- Siddiqui, M.R.H.; Adil, S.; Nour, K.; Assal, M.; Al-Warthan, A. Ionic liquid behavior and high thermal stability of silver chloride nanoparticles: Synthesis and characterization. Arab. J. Chem. 2013, 6, 435–438. [Google Scholar] [CrossRef] [Green Version]
- Zaki, S.; Eltarahony, M.; Elkady, M.; Abd-El-Haleem, D. The Use of Bioflocculant and Bioflocculant-Producing Bacillus mojavensis Strain 32A to Synthesize Silver Nanoparticles. J. Nanomater. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Iqtedar, M.; Aslam, M.; Akhyar, M.; Shehzaad, A.; Abdullah, R.; Kaleem, A. Extracellular biosynthesis, characterization, optimization of silver nanoparticles (AgNPs) using Bacillus mojavensis BTCB15 and its antimicrobial activity against multidrug resistant pathogens. Prep. Biochem. Biotechnol. 2019, 49, 136–142. [Google Scholar] [CrossRef]
- Blacutt, A.A.; Mitchell, T.R.; Bacon, C.W.; Gold, S.E. Bacillus mojavensis RRC101 Lipopeptides Provoke Physiological and Metabolic Changes During Antagonism Against Fusarium verticillioides. Mol. Plant-Microbe Interact. 2016, 29, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Ben Ayed, H.; Hmidet, N.; Béchet, M.; Chollet, M.; Chataigné, G.; Leclère, V.; Jacques, P.; Nasri, M. Identification and biochemical characteristics of lipopeptides from Bacillus mojavensis A21. Process. Biochem. 2014, 49, 1699–1707. [Google Scholar] [CrossRef]
- Snook, M.E.; Mitchell, T.; Hinton, D.M.; Bacon, C.W. Isolation and Characterization of Leu7-Surfactin from the Endophytic Bacterium Bacillus mojavensis RRC 101, a Biocontrol Agent for Fusarium verticillioides. J. Agric. Food Chem. 2009, 57, 4287–4292. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, N.; Hu, J.; Wang, S. Isolation and characterization of a new iturinic lipopeptide, mojavensin A produced by a marine-derived bacterium Bacillus mojavensis B0621A. J. Antibiot. 2012, 65, 317–322. [Google Scholar] [CrossRef] [Green Version]
- Bahrami-Teimoori, B.; Nikparast, Y.; Hojatianfar, M.; Akhlaghi, M.; Ghorbani, R.; Pourianfar, H.R. Characterisation and antifungal activity of silver nanoparticles biologically synthesised by Amaranthus retroflexus leaf extract. J. Exp. Nanosci. 2017, 12, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, S.; Poulose, E.K. Silver nanoparticles: Mechanism of antimicrobial action, synthesis, medical applications, and toxicity effects. Int. Nano Lett. 2012, 2, 32. [Google Scholar] [CrossRef] [Green Version]
- Amany, A.; Kheshen, E.; Sanaa, F.; Rab, G.E. Effect of reducing and protecting agents on size of silver nanoparticles and their anti-bacterial activity. J. Pharm. Chem. 2012, 4, 53–65. [Google Scholar]
- Krutyakov, Y.A.; Kudrinskiy, A.A.; Olenin, A.Y.; Lisichkin, G.V. Synthesis and properties of silver nanoparticles: Advances and prospects. Russ. Chem. Rev. 2008, 77, 233–257. [Google Scholar] [CrossRef]
- Xu, R.; Wang, D.; Zhang, J.; Li, Y. Shape-Dependent Catlytic Activity of Silver Nanoparticles for the Oxidation of Styrene. Chem. Asian J. 2006, 1, 888–893. [Google Scholar] [CrossRef]
- Abd Alamer, I.S.; Tomah, A.A.; Li, B.; Zhang, J.-Z. Isolation, Identification and Characterization of Rhizobacteria Strains for Biological Control of Bacterial Wilt (Ralstonia solanacearum) of Eggplant in China. Agriculture 2020, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Sotoodehnia, P.; Mazlan, N.; Mohd Saud, H.; Samsuri, W.A.; Habib, S.H.; Soltangheisi, A. Minimum inhibitory concentration of nano-silver bactericides for beneficial microbes and its effect on Ralstonia solanacearum and seed germination of Japanese Cucumber (Cucumis sativus). PeerJ 2019, 7, e6418. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, X.-L.; Zhu, S.-J.; Ojaghian, M.R.; Zhang, J.-Z. Biocontrol potential of Paenibacillus polymyxa against Verticillium dahliae infecting cotton plants. Biol. Control 2018, 127, 70–77. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. Mafft multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2011, 12, 335–337. [Google Scholar] [CrossRef]
- Sanchooli, N.; Saeidi, S.; Barani, H.K.; Sanchooli, E. In vitro antibacterial effects of silver nanoparticles synthesized using Verbena officinalis leaf extract on Yersinia ruckeri, Vibrio cholera and Listeria monocytogenes. Iran. J. Microbiol. 2018, 10, 400–408. [Google Scholar]
- Ali, K.A.; Yao, R.; Wu, W.; Masum, M.I.; Luo, J.; Wang, Y.; Zhang, Y.; An, Q.; Sun, G.; Li, B. Biosynthesis of silver nanoparticle from pomelo (Citrus Maxima) and their antibacterial activity against acidovorax oryzae RS-2. Mater. Res. Express 2020, 7, 015097. [Google Scholar] [CrossRef]
- Liu, H.; Tian, W.-X.; Ibrahim, M.; Li, B.; Zhang, G.-Q.; Zhu, B.; Xie, G.-L. Characterization of pilP, a gene required for twitching motility, pathogenicity, and biofilm formation of Acidovorax avenae subsp. avenae RS-1. Eur. J. Plant Pathol. 2012, 134, 551–560. [Google Scholar] [CrossRef]
- Saminu, S.; Saleh, M.I.A.; Abdulwahab, A.; Farra’u, U.; Babale, A.I. Evaluation of anti-diarrhoeal activity of L-citrulline in mice. J. Afr. Ass. Physiol. Sci. 2018, 6, 104–109. [Google Scholar]
- Ahmed, T.; Shahid, M.; Noman, M.; Niazi, M.B.K.; Zubair, M.; Almatroudi, A.; Khurshid, M.; Tariq, F.; Mumtaz, R.; Li, B. Bioprospecting a native silver-resistant Bacillus safensis strain for green synthesis and subsequent antibacterial and anticancer activities of silver nanoparticles. J. Adv. Res. 2020, 24, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, X.-L.; Zhu, S.-J.; Ojaghian, M.R.; Zhang, J.-Z. Data on the ultrastructural characteristics of Paenibacillus polymyxa isolates and biocontrol efficacy of P. polymyxa ShX301. Data Brief 2018, 21, 259–262. [Google Scholar] [CrossRef] [PubMed]
- D’Steel, R.G.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach; McGraw-Hill: New York, NY, USA, 1986. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers | Primer Sequence (5′–3′) |
|---|---|---|
| 16S | 27f | AGAGTTTGATCMTGGCTCAG |
| 1492r | GGYTACCTTGTTACGACTT | |
| gyrA | 42f | CAGTCAGGAAATGCGTACGTCCTT |
| 1066r | CAAGGTAATGCTCCAGGCATTGCT | |
| rpoB | 2292f | GACGTGGGATGGCTACAACT |
| 3354r | ATTGTCGCCTTTAACGATGG | |
| purH | 70f | ACAGAGCTTGGCGTTGAAGT |
| 1013r | GCTTCTTGGCTGAATGAAGG | |
| polC | 1505f | TTGTCGCTCAYAATGCAAGC |
| 2337r | YTCAAGCATTTCRTCTGTCG | |
| groEL | 550f | GAGCTTGAAGTKGTTGAAGG |
| 1497r | TGAGCGTGTWACTTTTGTWG |
| Species of Bacillus | Strain | GenBank No. | |||||
|---|---|---|---|---|---|---|---|
| 16S | gyrA | rpoB | purH | polC | groEL | ||
| Bacillus sp. | IMA13 | MZ310441.1 | MZ338583.1 | MZ338584.1 | MZ338585.1 | MZ338586.1 | MZ338587.1 |
| B. sonorensis | B-23154 | NR_116189.1 | EU138611.1 | EU138818.1 | EU138749.1 | EU138680.1 | EU138542.1 |
| B. pumilus | NRS-272 | NR_116191.1 | EU138655.1 | EU138862.1 | EU138793.1 | EU138724.1 | EU138586.1 |
| B. atrophaeus | NRS-213 | NR_116190.1 | EU138654.1 | EU138861.1 | EU138792.1 | EU138723.1 | EU138585.1 |
| B. subtilis | NRS-744 | NR_116192.1 | EU138658.1 | EU138865.1 | EU138796.1 | EU138727.1 | EU138589.1 |
| B. subtilis subsp. spizizenii | B-23049 | NR_116187.1 | EU138602.1 | EU138809.1 | EU138740.1 | EU138671.1 | EU138533.1 |
| B. subtilis subsp.inaquosorum | B-23052 | NR_116188.1 | EU138605.1 | EU138812.1 | EU138743.1 | EU138674.1 | EU138536.1 |
| B. vallismortis | B-14890 | NR_116186.1 | EU138601.1 | EU138808.1 | EU138739.1 | EU138670.1 | EU138532.1 |
| B. mojavensis | B-14698 | NR_116185.1 | EU138598.1 | EU138805.1 | EU138736.1 | EU138667.1 | EU138529.1 |
| B. subtilis subsp. subtilis | 168 | NC_000964.3:30279-31832 | NC_000964.3: 6994-9459 | NC_000964.3: 121919-125500 | NC_000964.3: 708594-710132 | NC_000964.3: 1727133-1731446 | NC_000964.3: 650234-651868 |
| B. cereus | ATCC 14579 | NC_004722.1: 9187-10741 | NC_004722.1: 6475-8946 | NC_004722.1: 113908-117441 | NC_004722.1: 309016-310551 | NC_004722.1: 3794523-3790222 | NC_004722.1: 257826-259460 |
| B. amyloliquefaciens | ATCC 23350 | NC_014551.1 9765-11318 | NC_014551.1: 7010-9469 | NC_014551.1: 122979-126560 | NC_014551.1: 668235-669773 | NC_014551.1: 1728311-1732624 | NC_014551.1: 572972-574606 |
| B. licheniformis | ATCC 14580 | NC_006322.1: 9713-11250 | NC_006322.1: 6900-9368 | NC_006270.3: 120951-124532 | NC_006322.1: 707082-708620 | NC_006322.1: 1832492-1836808 | NC_006322.1: 626726-628360 |
| B. amyloliquefaciens subsp. plantarum | FZB42 | NC_009725.1: 9760-11314 | NC_009725.1: 7081-9462 | NC_009725.1: 122626-126216 | NC_009725.1: 670548-672086 | NC_009725.1: 1642186-1646499 | NC_009725.1: 619805-621439 |
| B. amyloliquefaciens | Mito-5-13 | AB610829.1 | AB612173.1 | AB615267.1 | AB615405.1 | AB612200.1 | AB611006.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd Alamer, I.S.; Tomah, A.A.; Ahmed, T.; Li, B.; Zhang, J. Biosynthesis of Silver Chloride Nanoparticles by Rhizospheric Bacteria and Their Antibacterial Activity against Phytopathogenic Bacterium Ralstonia solanacearum. Molecules 2022, 27, 224. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27010224
Abd Alamer IS, Tomah AA, Ahmed T, Li B, Zhang J. Biosynthesis of Silver Chloride Nanoparticles by Rhizospheric Bacteria and Their Antibacterial Activity against Phytopathogenic Bacterium Ralstonia solanacearum. Molecules. 2022; 27(1):224. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27010224
Chicago/Turabian StyleAbd Alamer, Iman Sabah, Ali Athafah Tomah, Temoor Ahmed, Bin Li, and Jingze Zhang. 2022. "Biosynthesis of Silver Chloride Nanoparticles by Rhizospheric Bacteria and Their Antibacterial Activity against Phytopathogenic Bacterium Ralstonia solanacearum" Molecules 27, no. 1: 224. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27010224