
Characterization of Collagen from Three Genetic Lines (Gray, Red and F1) of Oreochromis niloticus (Tilapia) Skin in Young and Old Adults



 ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
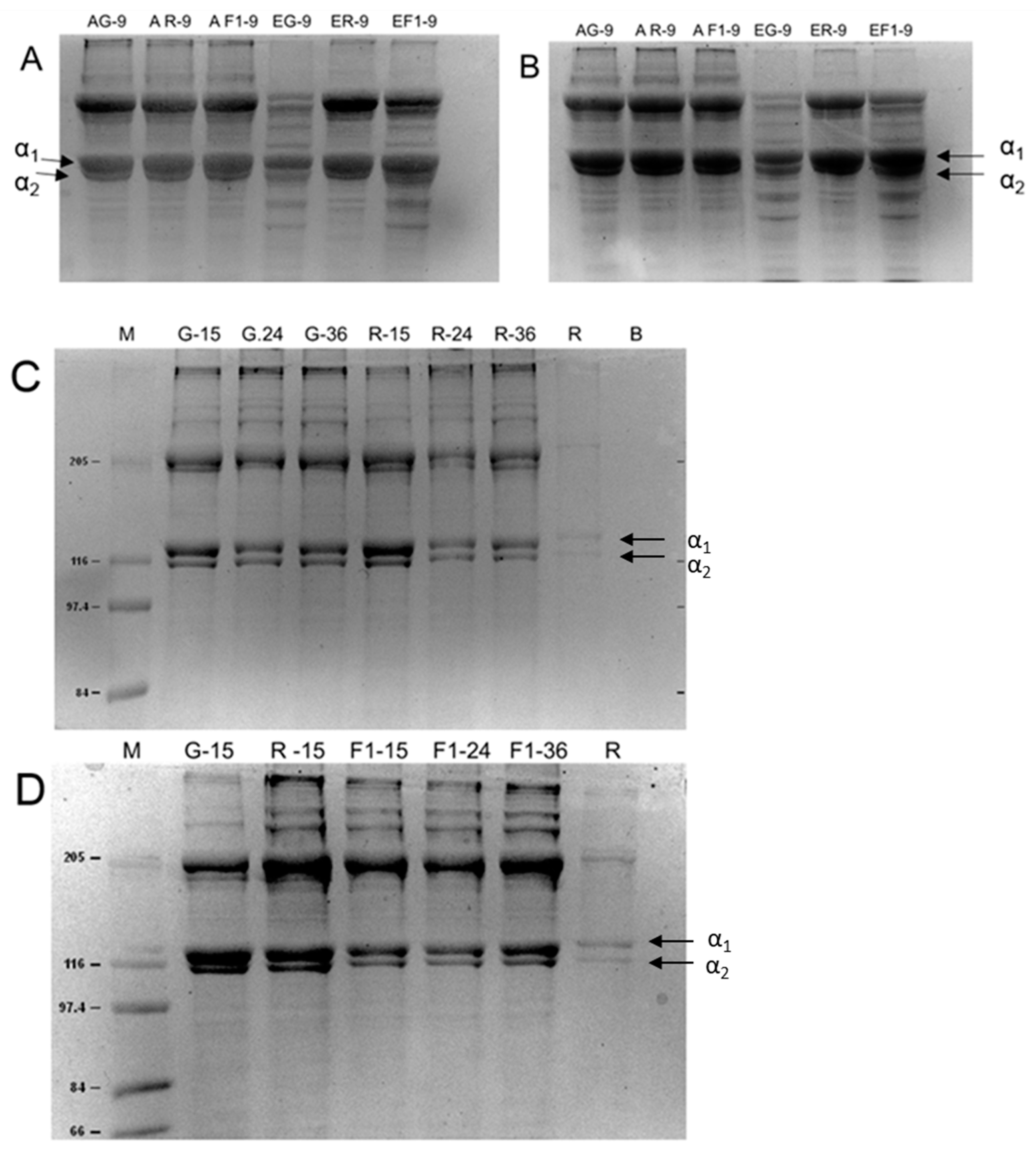
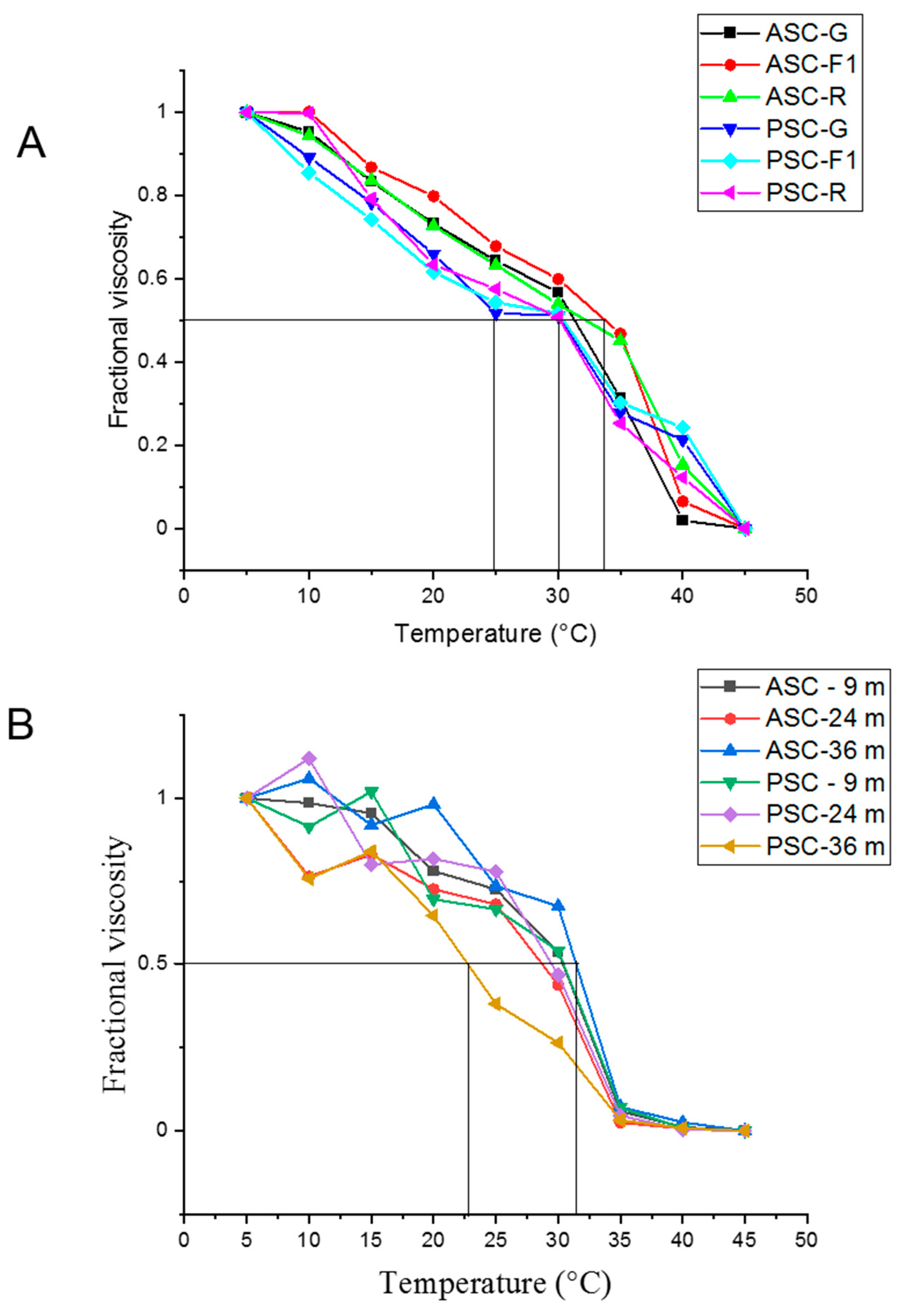
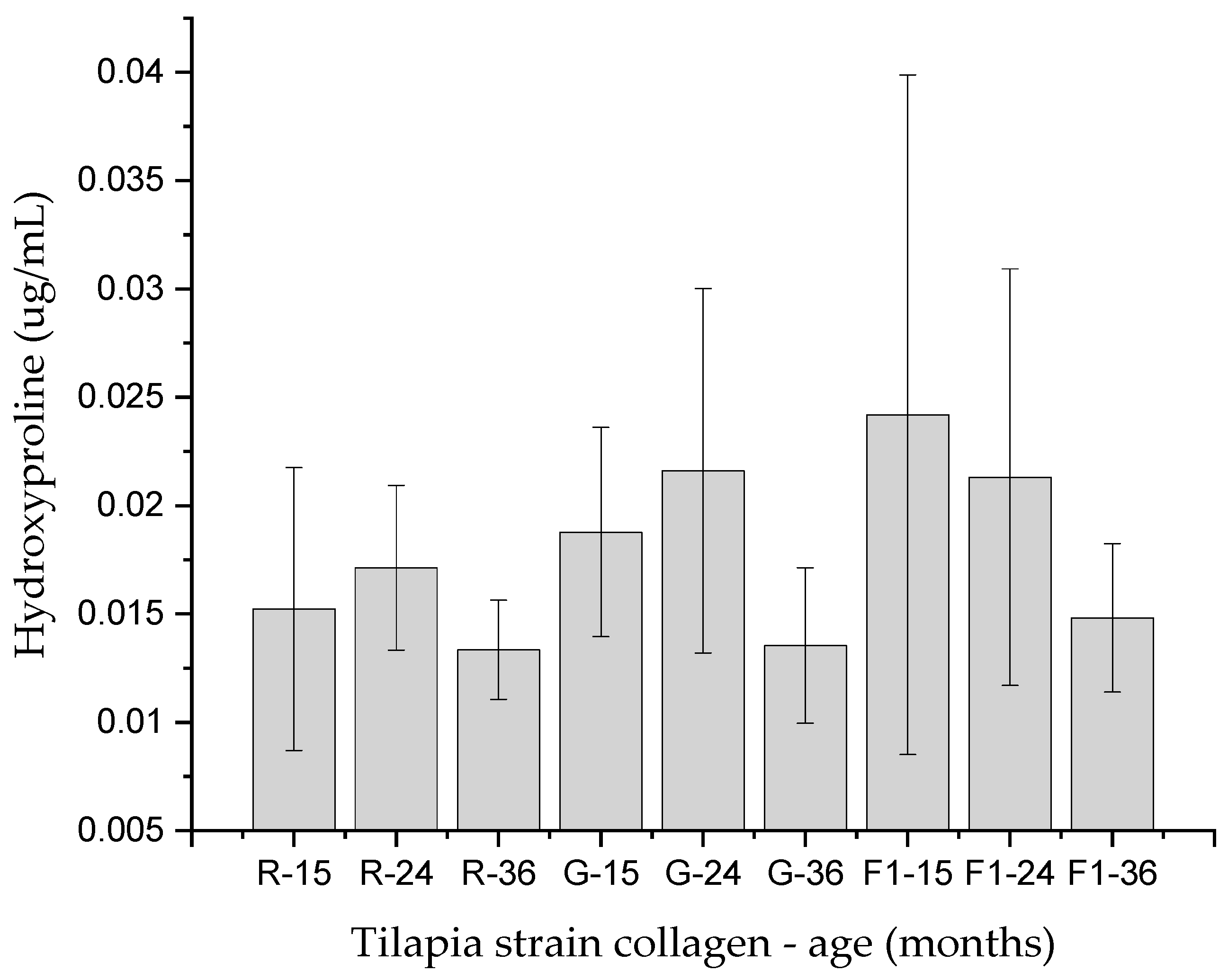
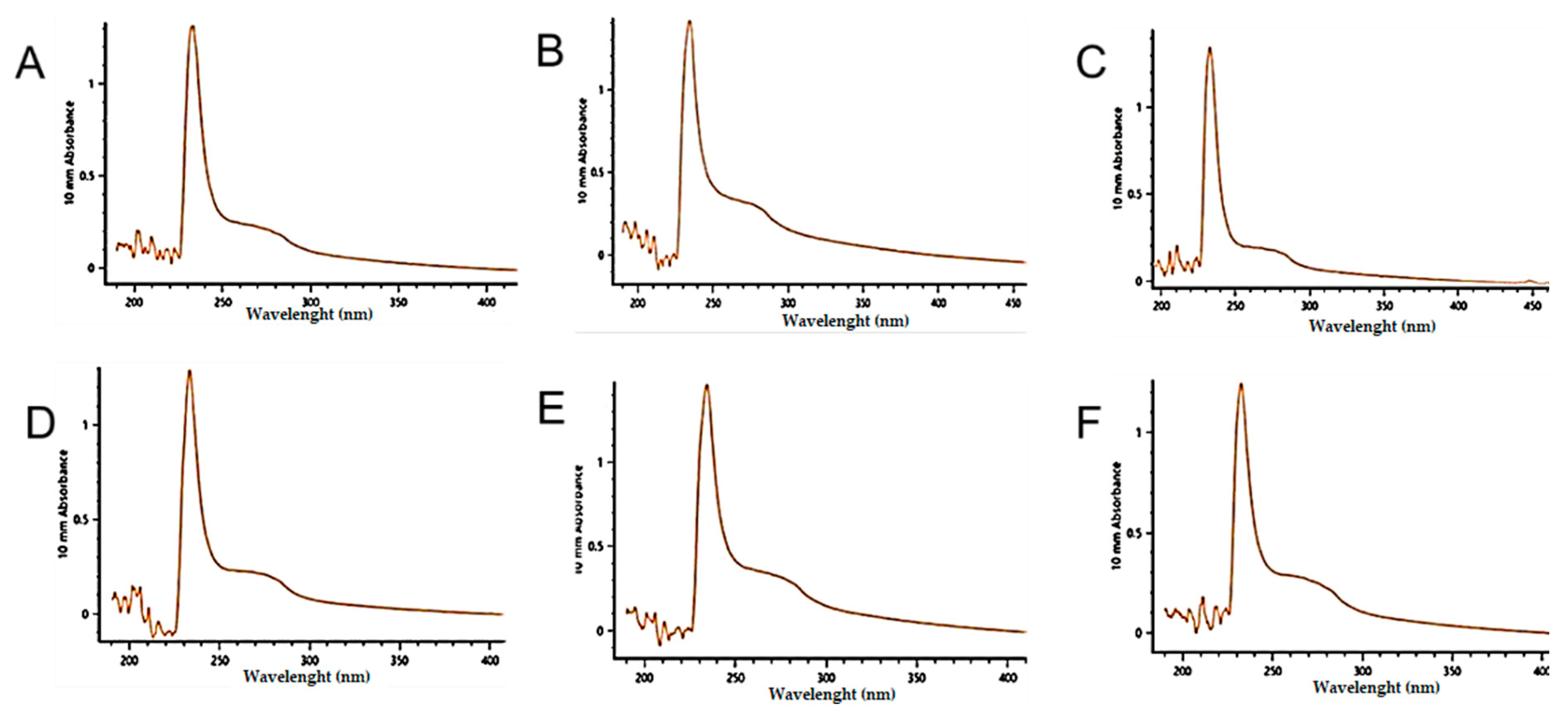
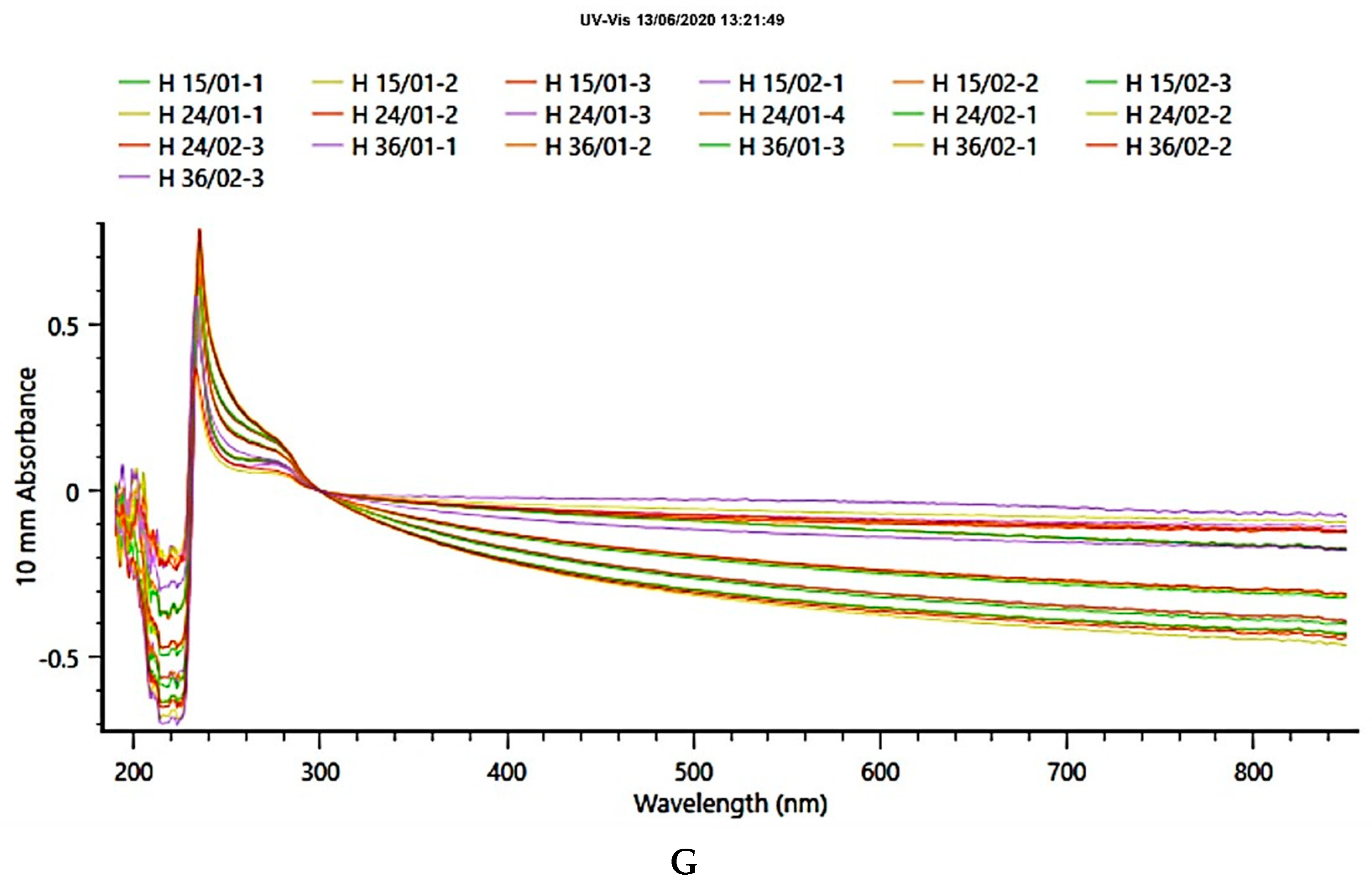
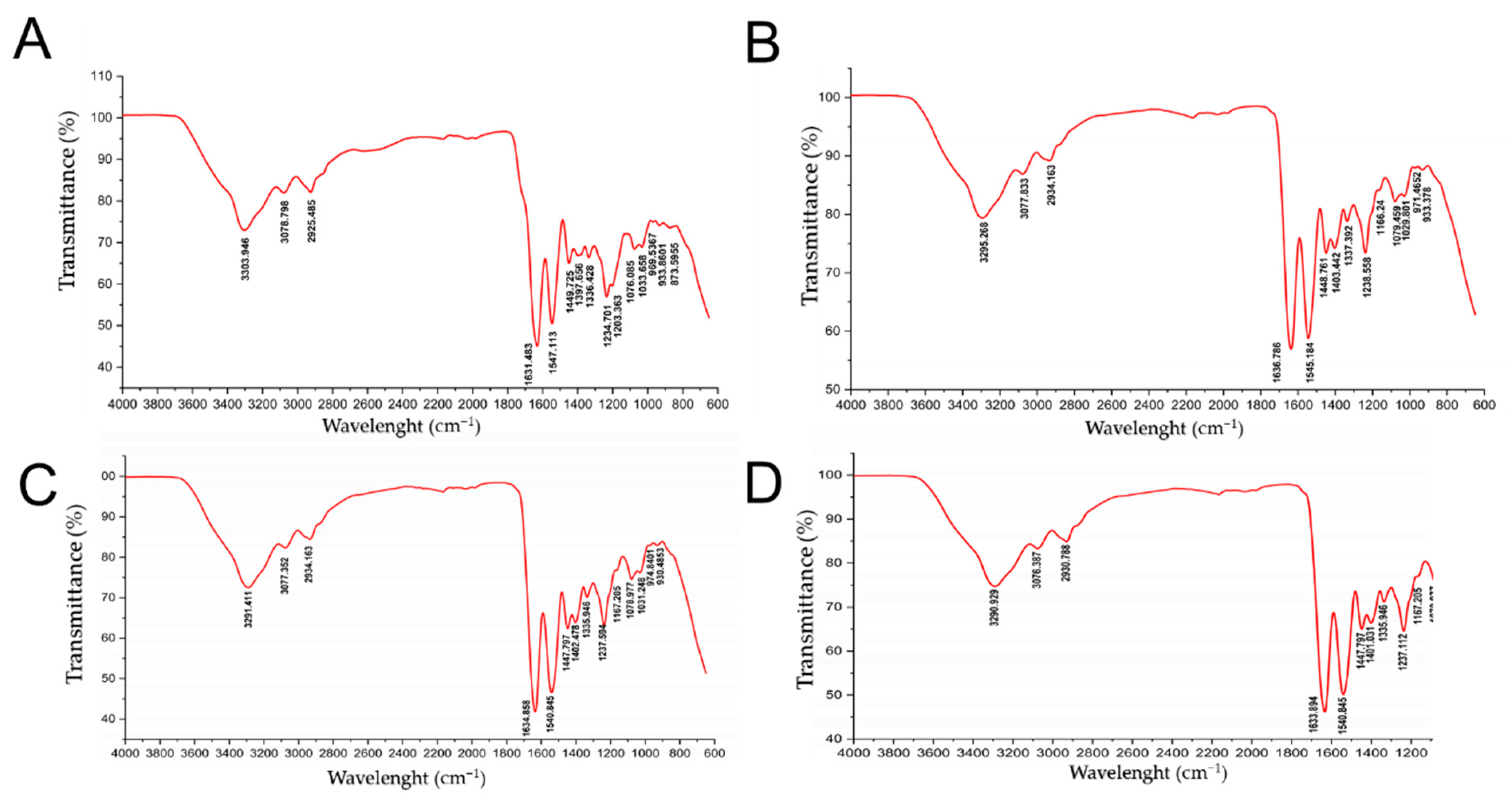
2. Results
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Collagen Extraction
4.3. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.4. Performance
4.5. Denaturation Temperature Determination (Based on Viscosity)
4.6. Hydroxyproline Quantification
4.7. Ultraviolet (UV) Scanning
4.8. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
4.9. X-ray Diffraction
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- León-López, A.; Morales-Peñaloza, A.; Martínez-Juárez, V.M.; Vargas-Torres, A.; Zeugolis, D.I.; Aguirre-Álvarez, G. Hydrolyzed Collagen—Sources and Applications. Molecules 2019, 24, 4031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupi, O. Prions in dermatology. J. Am. Acad. Dermatol. 2002, 46, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.H.; Lien, C.Y. Mini review of trace metal contamination status in East China Sea sediment. Mar. Pollut. Bull. 2020, 152, 110874. [Google Scholar] [CrossRef] [PubMed]
- Esiukova, E.; Zobkov, M.; Chubarenko, I. Data on microplastic contamination of the Baltic Sea bottom sediment samples in 2015–2016. Data Brief 2019, 28, 104887. [Google Scholar] [CrossRef]
- Barboza, L.G.A.; Lopes, C.; Oliveira, P.; Bessa, F.; Otero, V.; Henriques, B.; Raimundo, J.; Caetano, M.; Vale, C.; Guilhermino, L. Microplastics in wild fish from North East Atlantic Ocean and its potential for causing neurotoxic effects, lipid oxidative damage, and human health risks associated with ingestion exposure. Sci. Total Environ. 2019, 717, 134625. [Google Scholar] [CrossRef]
- Kakimoto, S.; Yoshimitsu, M.; Akutsu, K.; Kiyota, K.; Fujiwara, T.; Watanabe, T.; Kajimura, K.; Yamano, T. Concentrations of total mercury and methylmercury in red snow crabs (Chionoecetes japonicus) caught off the coast of Japan. Mar. Pollut. Bull. 2019, 145, 1–4. [Google Scholar] [CrossRef]
- FAO-Food and Agriculture Organization of the United Nations the State of World Fisheries and Aquaculture. Available online: http://www.fao.org/3/i9540en/i9540en.pdf (accessed on 15 October 2021).
- Hemker, A.K.; Nguyen, L.T.; Karwe, M.; Salvi, D. Effects of pressure-assisted enzymatic hydrolysis on functional and bioactive properties of tilapia (Oreochromis niloticus) by-product protein hydrolysates. LWT 2019, 122, 109003. [Google Scholar] [CrossRef]
- Ramshaw, J.A.M.; Shah, N.K.; Brodskyb, B. Gly-X-Y Tripeptide Frequencies in Collagen: A Context for Host–Guest Triple-Helical Peptides. J. Struct. Biol. 1998, 122, 86–91. [Google Scholar] [CrossRef]
- Berisio, R.; Granata, V.; Vitagliano, L.; Zagari, A. Imino Acids and Collagen Triple Helix Stability: Characterization of Collagen-like Polypeptides Containing Hyp-Hyp-Gly Sequence Repeats. J. Am. Chem. Soc. 2004, 126, 11402–11403. [Google Scholar] [CrossRef]
- Xu, S.; Gu, M.; Wu, K.; Li, G. Unraveling the Role of Hydroxyproline in Maintaining the Thermal Stability of the Collagen Triple Helix Structure Using Simulation. J. Phys. Chem. B 2019, 123, 7754–7763. [Google Scholar] [CrossRef]
- Ghanaeian, A.; Soheilifard, R. Mechanical elasticity of proline-rich and hydroxyproline-rich collagen-like triple-helices studied using steered molecular dynamics. J. Mech. Behav. Biomed. Mater. 2018, 86, 105–112. [Google Scholar] [CrossRef]
- Akita, M.; Nishikawa, Y.; Shigenobu, Y.; Ambe, D.; Morita, T.; Morioka, K.; Adachi, K. Correlation of proline, hydroxyproline and serine content, denaturation temperature and circular dichroism analysis of type I collagen with the physiological temperature of marine teleosts. Food Chem. 2020, 329, 126775. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.C.; Contin, R.F. Changes in the concentration of free and collagen-bound hydroxyproline in muscle tissue of the bluehead wrasse (Thalassoma bifasciatum) as a function of age. Comp. Biochem. Physiol. Part A Physiol. 1980, 66, 343–346. [Google Scholar] [CrossRef]
- Muyonga, J.H.; Cole, C.G.B.; Duodu, K.G. Characterisation of acid soluble collagen from skins of young and adult Nile perch (Lates niloticus). Food Chem. 2004, 85, 81–89. [Google Scholar] [CrossRef]
- Prieto, L.Z.A.; Salirrosas, F.R.D.; Arqueros, A.M.; Sanchez, T.L.C.; Gastañadui, R.D.; Fernádez, R.R. Obtención de Machos YY de Oreochromis Niloticus, Tilapia; EDUNT: Trujillo, Perú, 2018. [Google Scholar]
- Gómez-Marquez, J.L.; Peña-Mendoza, B.; Salgado-Ugarte, I.H.; Arredondo-Figueroa, J.L. Age and growth of the tilapia, Oreochromis niloticus (Perciformes: Cichlidae) from a tropical shallow lake in Mexico. Rev. Biol. Trop. 2008, 56, 875–884. [Google Scholar] [PubMed] [Green Version]
- Li, J.; Wang, M.C.; Qiao, Y.; Tian, Y.Y.; Liu, J.H.; Qin, S.; Wu, W.H. Extraction and characterization of type I collagen from skin of tilapia (Oreochromis niloticus) and its potential application in biomedical scaffold material for tissue engineering. Process. Biochem. 2018, 74, 156–163. [Google Scholar] [CrossRef]
- FAO-Food and Agriculture Organization of the United Nations Cultured Aquatic Species Information Programme. Available online: https://www.fao.org/fishery/collection/cultured-species/en (accessed on 15 October 2021).
- Romana-Eguia, M.R.R.; Ikeda, M.; Basiao, Z.U.; Taniguchi, N. Genetic diversity in farmed Asian Nile and red hybrid tilapia stocks evaluated from microsatellite and mitochondrial DNA analysis. Aquaculture 2004, 236, 131–150. [Google Scholar] [CrossRef]
- Arqueros, M.; Tuesta, L.S.; Prieto, Z. Diferenciación genética de tilapia roja y gris (Oreochromis niloticus) mediante microsa- télites y marcadores SCAR como indicadores del sexo genético. Rev. Peru. Biol. 2017, 24, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Parichy, D.M. Evolution of pigment cells and patterns: Recent insights from teleost fishes. Curr. Opin. Genet. Dev. 2021, 69, 88–96. [Google Scholar] [CrossRef]
- Wang, L.-M.; Luo, M.-K.; Yin, H.-R.; Zhu, W.-B.; Fu, J.-J.; Dong, Z.-J. Effects of background adaptation on the skin color of Malaysian red tilapia. Aquaculture 2020, 521, 735061. [Google Scholar] [CrossRef]
- Alfaro, A.D.T.; Fonseca, G.G.; Balbinot, E.; Machado, A.; Prentice, C. Physical and chemical properties of wami tilapia skin gelatin. Food Sci. Technol. 2013, 33, 592–595. [Google Scholar] [CrossRef] [Green Version]
- Le, T.M.T.; Nguyen, V.M.; Tran, T.T.; Takahashi, K.; Osako, K. Comparison of acid-soluble collagen characteristic from three important freshwater fish skins in Mekong Delta Region, Vietnam. J. Food Biochem. 2020, 44, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Moody, M.W.; Portier, R.J.; Bell, J.; Schexnayder, A.M.A.; Losso, J.N. Biochemical Properties of Black Drum and Sheepshead Seabream Skin Collagen. J. Agric. Food Chem. 2003, 51, 8088–8092. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, W.; Li, G. Isolation and characterisation of collagens from the skin of largefin longbarbel catfish (Mystus macropterus). Food Chem. 2009, 115, 826–831. [Google Scholar] [CrossRef]
- Chen, S.; Tang, L.; Su, W.; Weng, W.; Osako, K.; Tanaka, M. Separation and characterization of alpha-chain subunits from tilapia (Tilapia zillii) skin gelatin using ultrafiltration. Food Chem. 2015, 188, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Ohno, Y.; Miyauchi, Y.; Uchida, N. Fish skin type I collagen: Wide distribution of an α3 subunit in teleosts. Comp. Biochem. Physiol. Part B Biochem. 1987, 88, 27–34. [Google Scholar] [CrossRef]
- Gistelinck, C.; Gioia, R.; Gagliardi, A.; Tonelli, F.; Marchese, L.; Bianchi, L.; Landi, C.; Bini, L.; Huysseune, A.; Witten, P.E.; et al. Zebrafish Collagen Type I: Molecular and Biochemical Characterization of the Major Structural Protein in Bone and Skin. Sci. Rep. 2016, 6, 21540. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Hou, H.; Li, B.; Zhang, Y. Characterization of acid- and pepsin-soluble collagen extracted from the skin of Nile tilapia (Oreochromis niloticus). Int. J. Biol. Macromol. 2017, 99, 8–14. [Google Scholar] [CrossRef]
- Pal, G.K.; Suresh, P. Comparative assessment of physico-chemical characteristics and fibril formation capacity of thermostable carp scales collagen. Mater. Sci. Eng. C 2017, 70, 32–40. [Google Scholar] [CrossRef]
- Chen, J.; Li, L.; Yi, R.; Xu, N.; Gao, R.; Hong, B. Extraction and characterization of acid-soluble collagen from scales and skin of tilapia (Oreochromis niloticus). LWT 2016, 66, 453–459. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bi, C.; Li, X.; Xin, Q.; Han, W.; Shi, C.; Guo, R.; Shi, W.; Qiao, R.; Wang, X.; Zhong, J. Effect of extraction methods on the preparation of electrospun/electrosprayed microstructures of tilapia skin collagen. J. Biosci. Bioeng. 2019, 128, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T. Isolation of collagen from fish waste material—skin, bone and fins. Food Chem. 2000, 68, 277–281. [Google Scholar] [CrossRef]
- Bhagwat, P.; Dandge, P. Isolation, characterization and valorizable applications of fish scale collagen in food and agriculture industries. Biocatal. Agric. Biotechnol. 2016, 7, 234–240. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Kuo, J.-M.; Wu, S.-J.; Tsai, H.-T. Isolation and characterization of fish scale collagen from tilapia (Oreochromis sp.) by a novel extrusion–hydro-extraction process. Food Chem. 2016, 190, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Rios, D.E.; Rios, L.A.; Montoya, J.E.Z. Kinetic modeling of the alkaline deproteinization of Nile-tilapia skin for the production of collagen. Heliyon 2020, 6, e03854. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weight of Fillet and Subproducts | Age (Months) | ||
|---|---|---|---|
| 9 | 15 | 36 | |
| % w | |||
| Fillet | 33.04 | 29.13 | 32.84 |
| Subproducts skin weight | 3.55 | 3.01 | 3.25 |
| Subproducts total weight | 63.41 | 67.86 | 63.91 |
| Collagen Extraction Method | Skin Weight (g) | Lyophilized Collagen Weight (g) | Yield (%) |
|---|---|---|---|
| ASC Red | 8.93 ± 0.61 | 1.70 ± 0.16 | 19.04 ± 0.59 |
| ASC Gray | 6.93 ± 1.37 | 1.39 ± 0.12 | 20.06 ± 2.20 |
| ASC F1 | 8.53 ± 0.42 | 1.73 ± 0.16 | 20.28 ± 1.86 |
| PSC Red | 8.53 ± 0.42 | 1.80 ± 0.12 | 21.10 ± 1.59 |
| PSC Gray | 6.93 ± 1.37 | 1.47 ± 0.29 | 21.21 ± 1.14 |
| PSC F1 | 8.53 ± 0.42 | 1.80 ± 0.12 | 21.10 ± 1.59 |
| Region | CCS | R-9 | G-9 | F1-9 | R-15 | G-15 | F1-15 | R-24 | G-24 | F1-24 | R-36 | G-36 | F1-36 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amide A | 3304 | 3290 | 3290 | 3290 | 3295 | 3295 | 3294 | 3290 | 3290 | 3290 | 3292 | 3290 | 3290 |
| Amide B | 2925 | 2940 | 2940 | 2940 | 2933 | 2939 | 2929 | 2945 | 2940 | 2930 | 2931 | 2931 | 2930 |
| Amide I | 1631 | 1640 | 1640 | 1630 | 1632 | 1633 | 1631 | 1630 | 1640 | 1630 | 1632 | 1632 | 1631 |
| Amide II | 1547 | 1540 | 1540 | 1540 | 1545 | 1545 | 1544 | 1540 | 1540 | 1540 | 1542 | 1540 | 1540 |
| Amide III | 1235 | 1240 | 1240 | 1240 | 1238 | 1238 | 1238 | 1240 | 1240 | 1240 | 1240 | 1239 | 1239 |
| Strain Collagen | Months | Peak 1 (°) | Peak 2 (°) |
|---|---|---|---|
| STD | 7.15 | 20.64 | |
| Gray | 9 | 7.25 | 21.80 |
| 15 | 7.47 | 21.17 | |
| 23 | 7.48 | 21.86 | |
| 36 | 7.54 | 21.60 | |
| Red | 9 | 7.47 | 21.00 |
| 15 | 7.47 | 20.96 | |
| 23 | 7.70 | 21.58 | |
| 36 | 7.40 | 21.15 | |
| F1 (H) | 9 | 7.33 | 20.82 |
| 15 | 7.78 | 21.28 | |
| 23 | 7.46 | 21.44 | |
| 36 | 7.44 | 21.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reátegui-Pinedo, N.; Salirrosas, D.; Sánchez-Tuesta, L.; Quiñones, C.; Jáuregui-Rosas, S.R.; Barraza, G.; Cabrera, A.; Ayala-Jara, C.; Martinez, R.M.; Baby, A.R.; et al. Characterization of Collagen from Three Genetic Lines (Gray, Red and F1) of Oreochromis niloticus (Tilapia) Skin in Young and Old Adults. Molecules 2022, 27, 1123. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27031123
Reátegui-Pinedo N, Salirrosas D, Sánchez-Tuesta L, Quiñones C, Jáuregui-Rosas SR, Barraza G, Cabrera A, Ayala-Jara C, Martinez RM, Baby AR, et al. Characterization of Collagen from Three Genetic Lines (Gray, Red and F1) of Oreochromis niloticus (Tilapia) Skin in Young and Old Adults. Molecules. 2022; 27(3):1123. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27031123
Chicago/Turabian StyleReátegui-Pinedo, Nataly, David Salirrosas, Linda Sánchez-Tuesta, Claudio Quiñones, Segundo R. Jáuregui-Rosas, Gabriela Barraza, Angelita Cabrera, Carmen Ayala-Jara, Renata Miliani Martinez, André Rolim Baby, and et al. 2022. "Characterization of Collagen from Three Genetic Lines (Gray, Red and F1) of Oreochromis niloticus (Tilapia) Skin in Young and Old Adults" Molecules 27, no. 3: 1123. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27031123