2.1. The Effect of Aire on Molecules Related to DC Tolerance
Studies have shown that immature DCs maintain tolerance through the expression of low levels of related cell surface molecules [
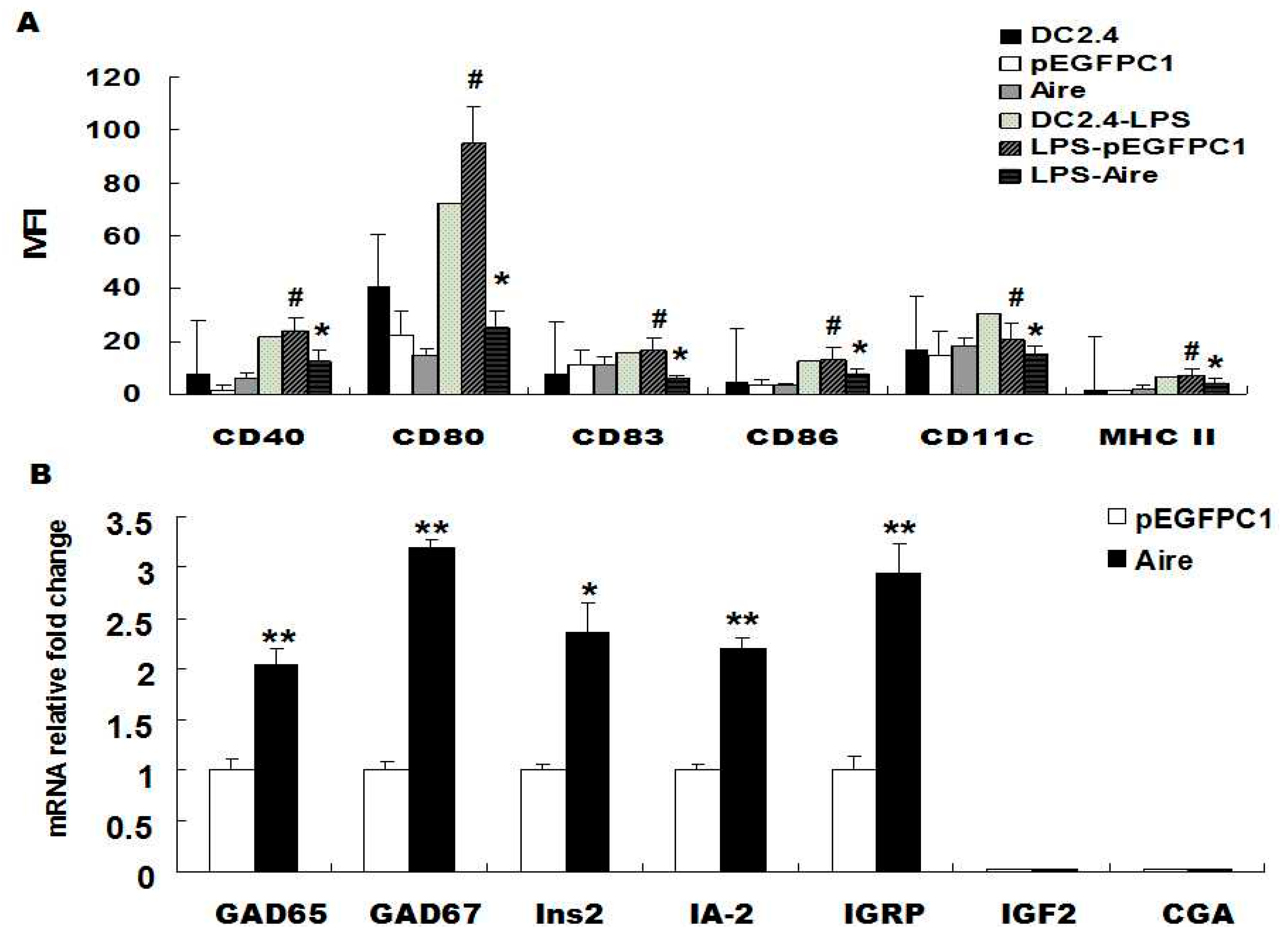
29]. Therefore, to investigate whether Aire could maintain the immature state of DCs, we examined the expression of cell surface molecules on unstimulated and lipopolysaccharide (LPS)-stimulated Aire cells. The results showed that the expression of CD40, CD80, CD83, CD86, CD11c and MHC-II was significantly lower in Aire cells stimulated with 10 μg/mL of LPS for 48 h compared to their expression levels in the control cells. No differences were observed in the expression levels of CD40, CD80, CD83, CD86, CD11c and major histocompatibility complex class
II (MHC II) between the two groups of unstimulated cells (
Figure 1A). The results were similar at 24 h post stimulation with LPS (
Supplementary Material, Figure S1). TRAs expressed in peripheral lymph nodes have been reported to be related to the clearance of autoreactive T cells and the maintenance of immune tolerance [
7,
8]. To verify the effects of Aire on the expression of T1D-related TRAs in DCs, we examined the mRNA expression levels of T1D-associated TRAs on Aire-expressing cells by quantitative reverse transcription polymerase chain reaction (RT-qPCR). The results showed that the mRNA levels of
glutamic acid decarboxylase 65/67 (
GAD65/67),
insulin-
2 (
Ins2),
insulinoma antigen 2 (IA-2) and
islet-specific glucose-6-phosphatase catalytic subunit-related protein (IGRP) were significantly elevated in the Aire cells compared with the control cells. The expression of
insulin-like growth factor 2 (IGF2) and
chorionic gonadotropin alpha chain (CGA) was not detected in either the Aire or control cells (
Figure 1B). In summary, Aire is one of the factors that maintains the immature state of DCs with or without stimulation by LPS and promotes the expression of T1D-related TRAs in DCs.
Figure 1.
Aire affected the maturation of DC2.4 and its TRA expression levels. (A) The expression of CD40, CD80, CD83, CD86, CD11c and MHC II were detected by FACS in Aire and control cells with or without stimulation with 10 μg/mL of LPS for 48 h; (B) GAD65/67, Ins2, IA-2, IGRP, IGF2, and CGA levels were detected by RT-qPCR in Aire and control cells. The data are shown as the expression levels relative to the expression of GAPDH and are depicted as the fold changes relative to the control cells normalized to 1. The data are expressed as the means±SD from three to five independent experiments. # p < 0.05, LPS stimulation vs. control; * p < 0.05, Aire vs. pEGFPC1, ** p < 0.01,Aire vs. pEGFPC1.
Figure 1.
Aire affected the maturation of DC2.4 and its TRA expression levels. (A) The expression of CD40, CD80, CD83, CD86, CD11c and MHC II were detected by FACS in Aire and control cells with or without stimulation with 10 μg/mL of LPS for 48 h; (B) GAD65/67, Ins2, IA-2, IGRP, IGF2, and CGA levels were detected by RT-qPCR in Aire and control cells. The data are shown as the expression levels relative to the expression of GAPDH and are depicted as the fold changes relative to the control cells normalized to 1. The data are expressed as the means±SD from three to five independent experiments. # p < 0.05, LPS stimulation vs. control; * p < 0.05, Aire vs. pEGFPC1, ** p < 0.01,Aire vs. pEGFPC1.
2.2. The Effect of Aire-Overexpressing DCs on the Tolerant Status of CD4+ T Cells
DC cells are capable of inducing the apoptosis of autoreactive CD4
+ T cells or functionally inactivating autoreactive CD4
+ T cells [
3,
4,
5], thereby reducing the production of interferon γ (IFN-γ), maintaining self-tolerance and eventually preventing the occurrence of autoimmune diseases. To investigate whether Aire-overexpressing DC2.4 cells impacted the apoptosis of autoreactive CD4
+ T cells, splenocytes derived from mice with STZ-induced T1D (STZ-T1D) and wild type (WT) mice were co-cultured with either Aire or control cells for 48 h. Then, we examined CD4
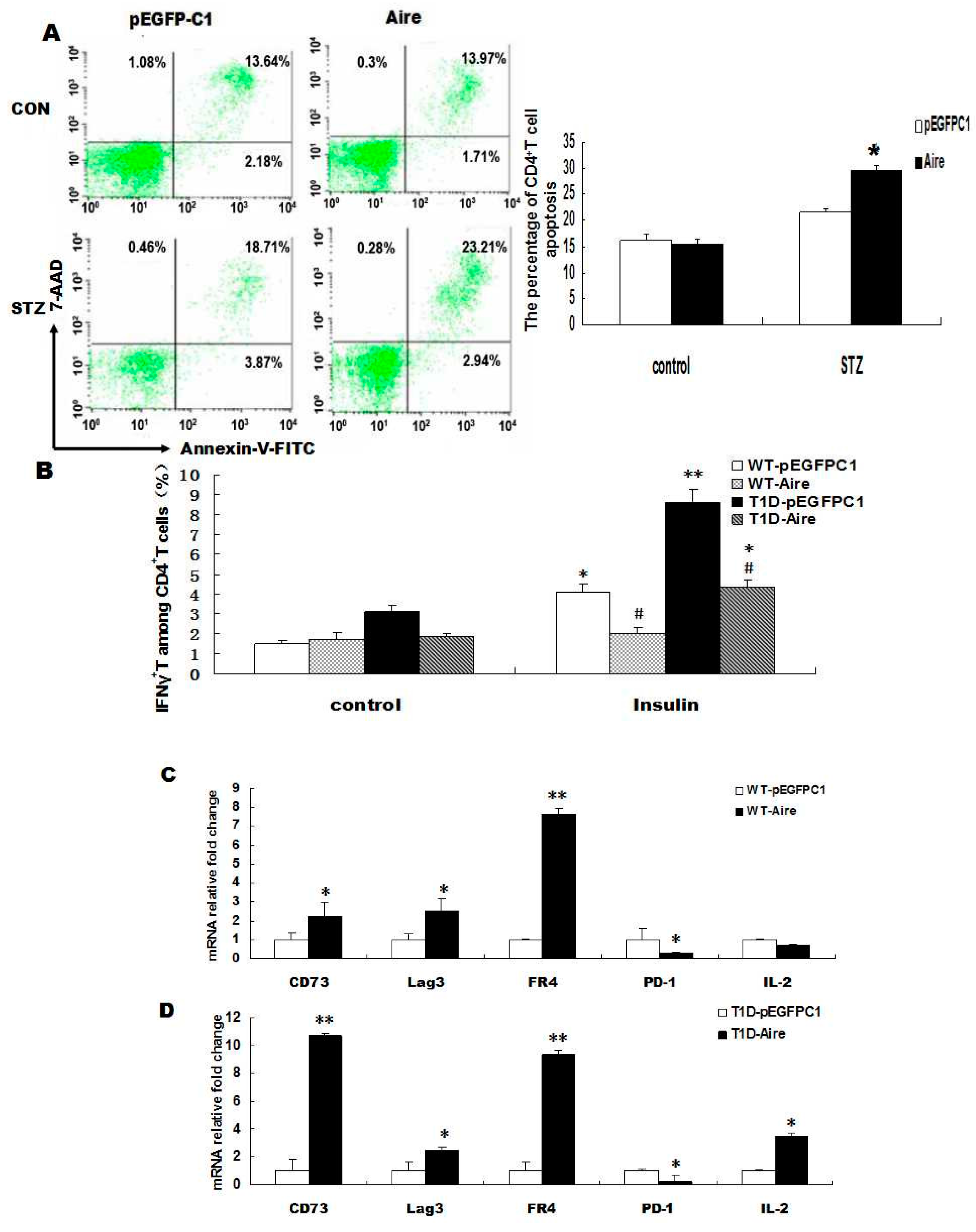
+ T cells apoptosis by flow cytometric analysis. The results showed that the apoptotic rate of STZ-T1D mouse-derived CD4
+ T cells was higher in the Aire group compared to the control group. In contrast, no significant difference was detected in the apoptotic rates of WT mouse-derived CD4
+ T cells between the two groups (
Figure 2A). These results demonstrated that Aire-overexpressing DCs induced the apoptosis of autoreactive CD4
+ T cells, suggesting that Aire DCs might induce CD4
+ T cell tolerance.
Next, we explored whether Aire-overexpressing DC2.4 cells induced functional inactivation of CD4
+ T cells. Splenocytes derived from STZ-T1D and WT mice were co-cultured with either Aire cells or control cells for 48 h and then stimulated with insulin for 24 h. The STZ-T1D mouse and WT mouse-derived splenocytes from the Aire group had significantly reduced numbers of CD4
+ IFN-γ
+ T cells compared to the splenocytes from the control group (
Figure 2B,
Figure S2). These results preliminarily indicated that Aire-transfected DCs might induce functional inactivation of CD4
+ T cells. However, the activation markers and other type of cytokines involved need to analyzed in future studies.
To confirm the tolerant status of CD4
+ T cells after induction by Aire-transfected DCs, we examined the mRNA expression levels of tolerance-related surface molecules by RT-qPCR, including CD73, lymphocyte-activation gene 3 (Lag3), folate receptor 4 (FR4) and programmed cell death protein 1 (PD-1), and the mRNA expression level of the cytokine interleukin 2 (IL-2) in T cells after co-culturing splenocytes and Aire cells [
30,
31]. The results showed that the mRNA expression levels of CD73, Lag3, FR4 and IL-2 were significantly higher in STZ-T1D mouse-derived splenocytes from the Aire group compared with those from the control group. In contrast, the expression of PD-1 was markedly decreased in the Aire group compared with the control group (
Figure 2D). The mRNA expression levels of CD73, Lag3 and FR4 were also higher in WT mouse-derived splenocytes from the Aire group, whereas PD-1 expression was lower in the Aire group compared to the control group. One possible explanation for the low expression of PD-1 in splenocytes from the Aire group may be related to PD-1’s dual role in T cells (
Figure 2C). In summary, Aire-transfected DCs promoted the apoptosis of STZ-T1D mouse-derived CD4
+ T cells, reduced the number of IFN-γ-producing autoreactive CD4
+ T cells in STZ-T1D and WT mouse and upregulated the expression of tolerance-related molecules in splenocytes, suggesting that Aire-overexpressing DCs may play a role in maintaining the tolerant state of CD4
+ T cells.
Figure 2.
Aire cells altered the tolerance status of CD4+ T cells. (A) Splenocytes from STZ-induced T1D mice and WT mice co-cultured with Aire or control cells for 48 h. The apoptosis of CD4+ T cells was analyzed by FACS using the Annexin V-7AAD kit; (B) Splenocytes from STZ-induced T1D mice or WT mice co-cultured with Aire or control cells for 48 h in the presence of insulin. The IFN-γ expression levels in CD4+ T cells were examined by FACS; (C,D) Splenocytes from STZ-induced T1D mice and WT mice co-cultured with Aire or control cells for 48 h. The CD73, Lag3, FR4, PD-1 and IL-2 levels were detected by RT-qPCR in the splenocytes. The results are presented as the percentage of positive cells ± SD (n = 5); each experiment was repeated at least three times. # p < 0.05, Insulin stimulation vs. control; * p < 0.05, ** p < 0.01, Aire vs. pEGFPC1.
Figure 2.
Aire cells altered the tolerance status of CD4+ T cells. (A) Splenocytes from STZ-induced T1D mice and WT mice co-cultured with Aire or control cells for 48 h. The apoptosis of CD4+ T cells was analyzed by FACS using the Annexin V-7AAD kit; (B) Splenocytes from STZ-induced T1D mice or WT mice co-cultured with Aire or control cells for 48 h in the presence of insulin. The IFN-γ expression levels in CD4+ T cells were examined by FACS; (C,D) Splenocytes from STZ-induced T1D mice and WT mice co-cultured with Aire or control cells for 48 h. The CD73, Lag3, FR4, PD-1 and IL-2 levels were detected by RT-qPCR in the splenocytes. The results are presented as the percentage of positive cells ± SD (n = 5); each experiment was repeated at least three times. # p < 0.05, Insulin stimulation vs. control; * p < 0.05, ** p < 0.01, Aire vs. pEGFPC1.
2.3. The Mechanism by Which Aire-Overexpressing DCs Induces Functional Inactivation of CD4+ T Cells
The T cell receptors (TCRs) located on the surface of the autoreactive T cells recognize the major histocompatibility complex class II (MHC II) complexes formed after DC-mediated antigen presentation. Antigen recognition activates the intracellular TCR signaling pathways and promotes the production of IFN-γ by activated T cells, resulting in autoimmune diseases [
32]. Therefore, the next issue to explore was whether Aire cells influenced IFN-γ production in CD4
+ T cells derived from STZ-T1D mice by affecting TCR signaling pathways. STZ-T1D and WT mouse-derived splenocytes were co-cultured with either Aire cells or control cells and then stimulated with an anti-CD3 antibody, concanavalin A (ConA) or insulin. Subsequently, we examined the key molecules of the TCR signaling pathways in CD4
+ T cells
by fluorescence-activated cell sorting (FACS), including Ca
2+ and phosphorylated extracellular signal-regulated kinase (p-ERK) [
33]. The results showed that the Ca
2+ and p-ERK levels after stimulation with the CD3 antibody and insulin were lower in the STZ-T1D and WT mouse-derived CD4
+ T cells from the Aire group compared with those from the control group. Although the levels of Ca
2+ and p-ERK were elevated in STZ-T1D and WT mouse-derived CD4
+ T cells upon stimulation with the unrelated control ConA compared to the unstimulated CD4
+T cells, there were no significant difference in the levels of Ca
2+ and p-ERK between the Aire and control groups after ConA stimulation (
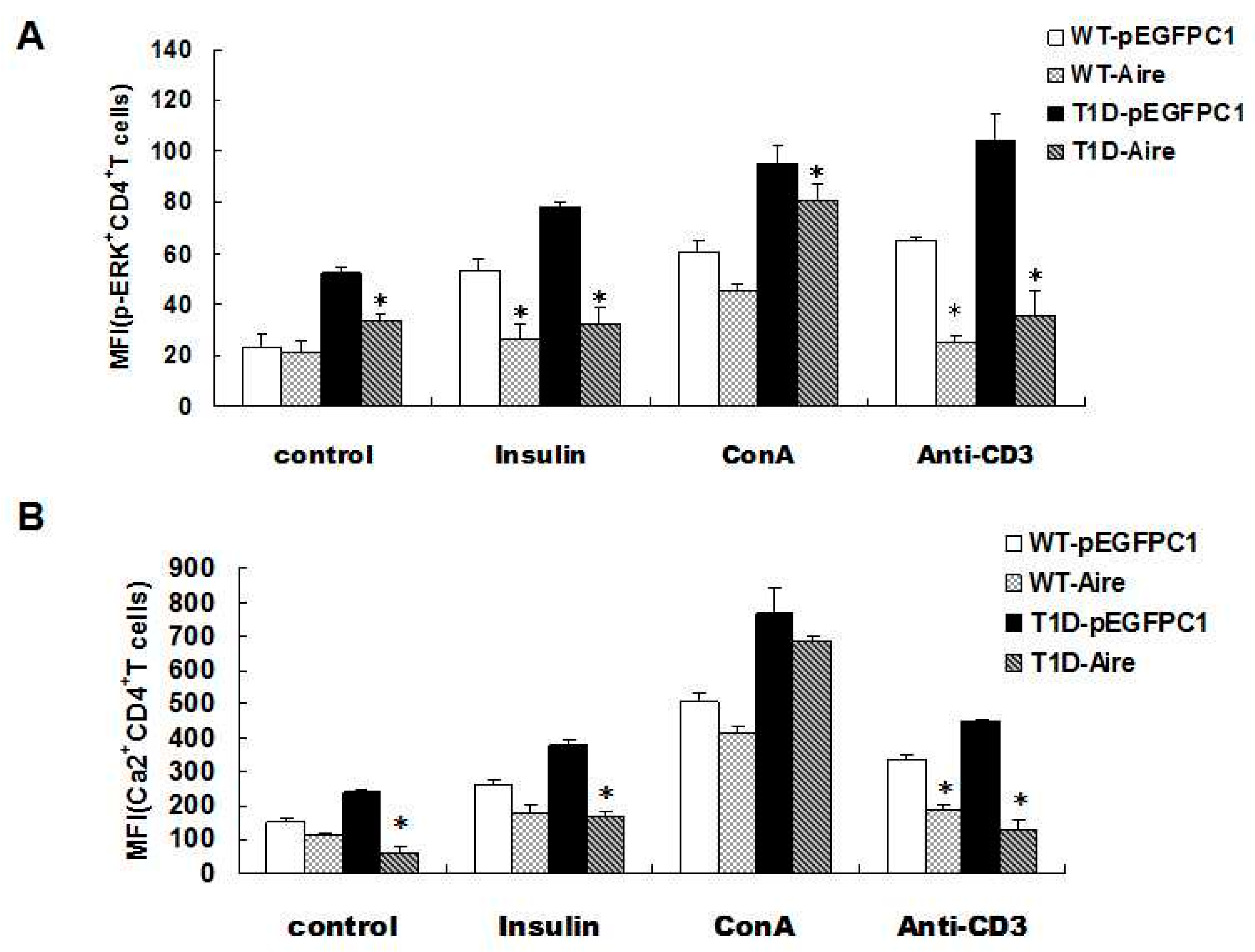
Figure 3A,B,
Supplementary Material, Figure S3). These results indicated that Aire-overexpressing DCs reduced the mRNA expression levels of key molecules of the TCR signaling pathways (Ca
2+ and p-ERK) in STZ-T1D mouse-derived CD4
+ T cells, resulting in the functional inactivation of autoreactive CD4
+ T cells. Thus, this may be one pathway by which Aire induces tolerance in autoreactive T cells.
Figure 3.
Aire cells affected TCR signaling in CD4+ T cells among the splenocytes from STZ-T1D mice. The splenocytes from STZ-induced T1D mice and WT mice co-cultured with Aire or control cells in the presence of insulin, ConA or an anti-CD3 antibody for 48 h. (A) The p-ERK expression levels; and (B) the concentration of Ca2+ in CD4+T cells were examined by FACS. The results are presented as the average percentage ± SD (n = 5); each experiment was repeated at least three times. * p < 0.05, Aire vs. pEGFPC1.
Figure 3.
Aire cells affected TCR signaling in CD4+ T cells among the splenocytes from STZ-T1D mice. The splenocytes from STZ-induced T1D mice and WT mice co-cultured with Aire or control cells in the presence of insulin, ConA or an anti-CD3 antibody for 48 h. (A) The p-ERK expression levels; and (B) the concentration of Ca2+ in CD4+T cells were examined by FACS. The results are presented as the average percentage ± SD (n = 5); each experiment was repeated at least three times. * p < 0.05, Aire vs. pEGFPC1.
2.4. The Effects of CD4+T Cells Induced by Aire-Overexpressing DCs on the Incidence of T1D in Mice
Our previous study demonstrated that the expression levels of IFN-γ, Ca
2+ and p-ERK were markedly reduced in CD4
+ T cells after splenocytes derived from WT mice were co-cultured with Aire cells and stimulated with insulin. These results led us to speculate whether Aire-induced CD4
+ T cells would inhibit or delay the occurrence of T1D in STZ-induced mice. In the following experiment, splenocytes derived from WT mice were co-cultured with Aire cells or control cells for 48h. The purity (
Supplementary Material, Figure S4A) and the rate of apoptotic T-cells (
Supplementary Material, Figure S4B) were detected. A low percentage of aptoptotic T cell was detected in each group, and there was no difference between the Aire-expressing DC group and control group. Then the splenocytes were intravenously injected into WT mice through the
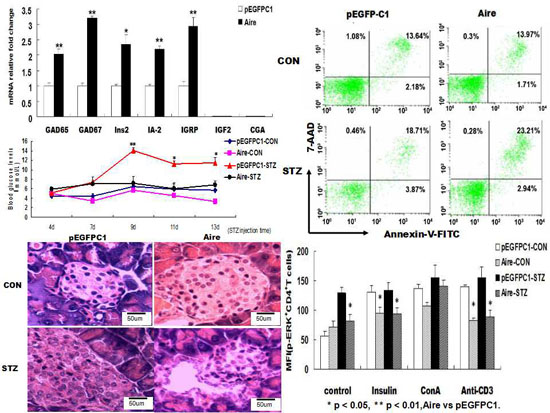
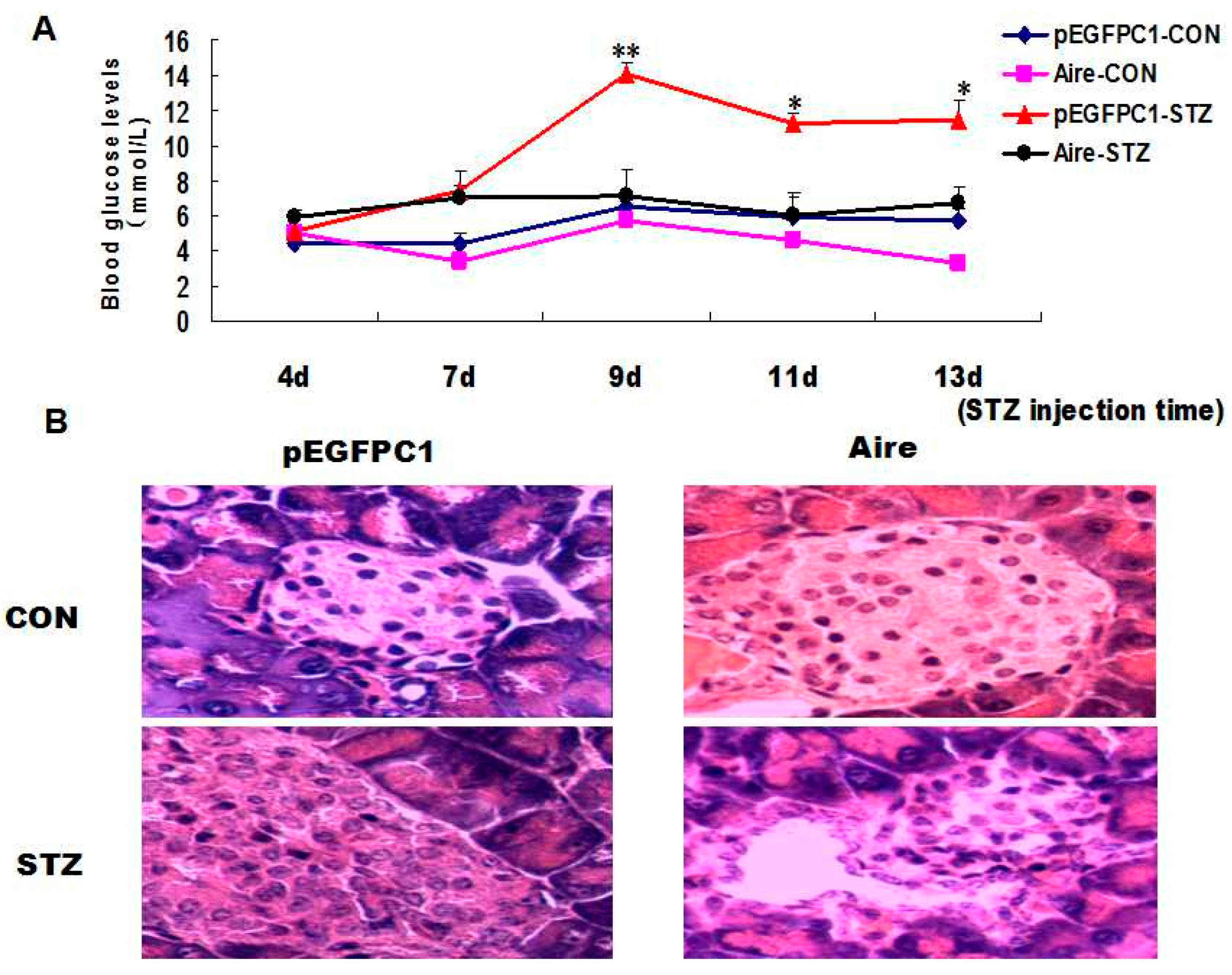
tail vein. Subsequently, we established a mouse model of T1D induced with STZ and monitored the blood glucose levels. The results showed that the T1D model mice developed typical symptoms. However, the blood glucose levels were significantly lower in mice transplanted with splenocytes of the Aire group compared to those in mice transplanted with splenocytes of the control group (
Figure 4A). After two weeks, the pancreatic tissues were collected and subjected to hematoxylin and eosin (H & E) staining. The degree of inflammatory infiltration was significantly lower in the pancreatic tissues of mice transplanted with splenocytes from the Aire group (
Figure 4B).
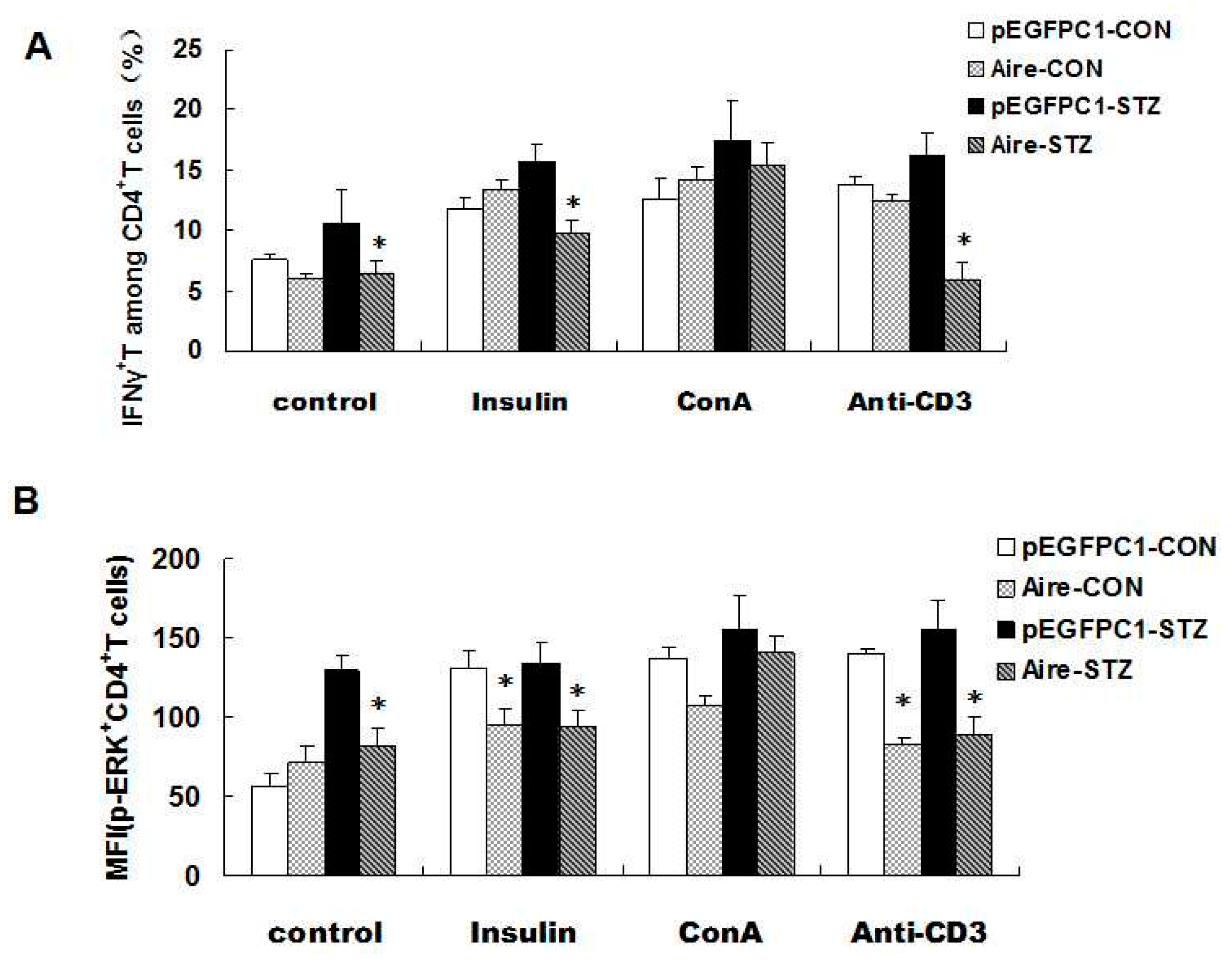
Finally, we investigated the expression of IFN-γ and p-EPK in CD4
+ T cells simulated with an anti-CD3 antibody, ConA and insulin. The results demonstrated that the expression of IFN-γ and p-EPK after anti-CD3 antibody and insulin treatment was significantly lower in CD4
+ T cells from mice transplanted with splenocytes from the Aire group. Moreover, no significant difference was detected between the Aire group and the control group following ConA stimulation (
Figure 5A,B), indicating that the two groups of splenocytes did not differ in cell viability. Taken together, our data suggested that Aire cell-induced splenocytes delayed the occurrence of T1D in STZ-induced mice.
Figure 4.
Adoptive transfer of Aire cell-co-cultured splenocytes delayed the development of type 1 diabetes. Splenocytes from WT mice were co-cultured with Aire or control cells for 48 h. Then, the lymphocytes were collected and injected into WT mice by the tail vein, followed by the induction of diabetes with STZ. (A) The blood glucose levels in mice transferred spleen lymphocytes; (B) the histology of pancreas of mice transferred spleen lymphocytes with H&E staining, n = 5 in each group, * p < 0.05, ** p < 0.01, Aire vs. pEGFPC1; Original magnification, ×200.
Figure 4.
Adoptive transfer of Aire cell-co-cultured splenocytes delayed the development of type 1 diabetes. Splenocytes from WT mice were co-cultured with Aire or control cells for 48 h. Then, the lymphocytes were collected and injected into WT mice by the tail vein, followed by the induction of diabetes with STZ. (A) The blood glucose levels in mice transferred spleen lymphocytes; (B) the histology of pancreas of mice transferred spleen lymphocytes with H&E staining, n = 5 in each group, * p < 0.05, ** p < 0.01, Aire vs. pEGFPC1; Original magnification, ×200.
Figure 5.
Adoptive transfer of Aire cells-co-cultured splenocytes suppressed autoreactive T cell activation by interfering with TCR signaling. Splenocytes from WT mice were co-cultured with Aire or control cells for 48 h. Then, the lymphocytes were isolated and injected into WT mice by tail vein followed by the induction of diabetes with STZ. The splenocytes were harvested and stimulated with insulin, ConA or CD3 antibody for 48 h prior to detection of the IFN-γ expression levels (A); and p-ERK expression levels (B) in CD4+ T cells by FACS. The results are presented as the mean values ± SD. Each experiment was repeated three to five times, and each group was compared with the pEGFPC1 group; * p < 0.05, Aire vs. pEGFPC1.
Figure 5.
Adoptive transfer of Aire cells-co-cultured splenocytes suppressed autoreactive T cell activation by interfering with TCR signaling. Splenocytes from WT mice were co-cultured with Aire or control cells for 48 h. Then, the lymphocytes were isolated and injected into WT mice by tail vein followed by the induction of diabetes with STZ. The splenocytes were harvested and stimulated with insulin, ConA or CD3 antibody for 48 h prior to detection of the IFN-γ expression levels (A); and p-ERK expression levels (B) in CD4+ T cells by FACS. The results are presented as the mean values ± SD. Each experiment was repeated three to five times, and each group was compared with the pEGFPC1 group; * p < 0.05, Aire vs. pEGFPC1.
2.5. Discussion
Aire is primarily expressed in mTECs in the central immune system. Aire induces central tolerance by regulating the expression of the TRA genes and inducing the negative selection of self-reactive T cells and the production of regulatory T cells [
2,
10,
11,
12]. In the periphery,
Aire is mainly expressed on DCs. However, the function and significance of Aire in the periphery remain unclear [
9,
17,
18,
19]. The present study primarily explored the effects of
Aire expression in peripheral DCs on the function and activity of peripheral CD4
+ T cells.
DCs can be divided into two major types based on their maturation status: immature and mature DCs. Immature and mature DCs display different phenotypes. Moreover, DCs with different maturation states exert distinct effects on T cells. Mature DCs induce effective immune responses by expressing costimulatory molecules and by highly expressing MHC II. Immature DCs often lack costimulatory molecules, express low levels of MHC II and exert immunosuppressive activities. Studies have shown that extrathymic Aire-expressing cells (eTACs) expressing low levels of CD80 and CD86 may serve as tolerant cells to induce antigen-specific immune tolerance. Additionally, stimulation of Toll-like receptors (TLRs) failed to reverse eTAC-induced immune tolerance [
34,
35,
36]. The above findings are consistent with our results obtained using Aire-overexpressing DC2.4 cells. The present study showed that both Aire-overexpressing DC2.4 cells and control cells expressed low levels of costimulatory molecules (CD40, CD80, CD83, CD86, and CD11c) and MHC II-like molecules, which was consistent with the phenotypic characteristics of immature DCs. DC maturation following treatment with the DC activator LPS was accompanied by the upregulation of the expression of costimulatory molecules and MHC II-like molecules on the cell surface, which was consistent with previous reports [
37]. However, the expression levels of costimulatory molecules and MHC II on the surface of the Aire cells were downregulated compared to the control cells after administration of the DC activator. Therefore, we speculated that Aire inhibited the expression of costimulatory molecules and MHC II in activated DCs, thereby maintaining the immature state of DCs and playing a role in maintaining peripheral immune tolerance.
One study showed that DCs present in the periphery also expressed TRAs and played a role similar to mTECs [
38] in which they induced the clearance of autoreactive T cells and prevented the incidence of autoimmune diseases. Aire
+ DCs expressed a high level of Ins2 compared to the total mouse splenocytes [
22]. Therefore, we speculated that the expression of Aire in peripheral DCs affected peripheral immune tolerance by regulating the expression of TRAs. Our results showed that Aire-transfected DCs significantly enhanced the expression of T1D-associated TRAs, such as GAD65/67, Ins2, IA-2 and IGRP. These data demonstrated that Aire expression in peripheral DCs might play a role similar to its role in the central immune system. Thus, Aire induced the clearance of peripheral autoreactive T cells and participated in immune tolerance through the regulation of the expression of TRAs on DCs.
Previous reports have shown that the
production of significant levels of IFN-γ by T cells is the main cause of autoimmune diseases such as T1D. The peripheral immune system maintains a state of tolerance by restricting IFN-γ
+ T cells to a low percentage of total CD4
+ T cells [
39,
40,
41]. In the present study, splenocytes derived from STZ-T1D mice and WT mice were co-cultured with Aire-overexpressing DCs and then stimulated with insulin, which resulted in a significant reduction in the number of CD4
+ IFN-γ
+ T cells among the total splenocytes. These results indicated that Aire-overexpressing DCs played a role in preventing the occurrence of T1D by inhibiting the production of IFN-γ by CD4
+ T cells or inducing the apoptosis of IFN-γ-producing autoreactive CD4
+ T cells.
CD4
+ T cells recognize specific antigens presented by MHC II molecules through TCRs and then receive stimulatory signals from the engaged TCRs. In the presence of synergistic actions of costimulatory molecules, TCR signaling pathways will be fully activated and further promote the production and release of downstream cytokines, such as IFN-γ, thereby eliciting immune responses. Loss of costimulatory molecules leads to the inactivation of TCR signaling pathways [
42]. The results of our previous experiments showed that Aire inhibited the expression of costimulatory molecules and MHC II during the process of DC activation. Therefore, we investigated whether the Aire-overexpressing DC-induced reduction in the number of CD4
+ IFN-γ
+ T cells in mouse splenocytes was caused by the inactivation of TCR signaling pathways in CD4
+ T cells. In the present study, splenocytes derived from STZ-T1D and WT mice were co-cultured with Aire-overexpressing DCs and then stimulated with insulin. The results showed that the levels of key molecules of the TCR signaling pathways (Ca
2+ and p-ERK) were significantly decreased in STZ-T1D and WT mouse-derived CD4
+ T cells after induction with Aire-overexpressing DCs. These results suggest a mechanism in which Aire suppresses the activity of CD4
+ T cells by inhibiting the expression of costimulatory molecules during the process of DC activation and inhibiting TCR signaling pathways in CD4
+ T cells, eventually leading to a reduction in the number of CD4
+ IFN-γ
+ T cells.
The expression of TRA genes in the thymus provides an opportunity for developing T cells to contact peripheral self-antigens and induces the apoptosis and clearance of CD4
+ T cells that exhibit high affinity for the TRAs [
43,
44,
45]. The above phenomenon also exists in the periphery [
3,
4]. The results of our previous experiments showed that DCs transfected with Aire exhibited significantly upregulated expression of T1D-related TRAs. Therefore, we examined CD4
+ T cell apoptosis in STZ-T1D and WT mouse-derived splenocytes after co-culturing with Aire-overexpressing DCs. Our results were consistent with previous reports in the literature that demonstrated that Aire-transfected DCs significantly enhanced the apoptosis of STZ-T1D mouse-derived CD4
+ T cells, whereas they exerted no significant effect on WT mouse-derived CD4
+ T cells. These results indicated that Aire induced the apoptosis of autoreactive CD4
+ T cells by promoting the expression of TRAs in DCs, which may represent another mechanism for the maintenance of immune tolerance. However, one cannot rule out the possibility that Aire-expressing cells may induce CD4
+ T cell tolerance through other means, such as direct cell-cell contact or soluble mediators. We have measured the level of some soluble mediators, such as TGF-β was upregulated in Aire overexpressing cells. This may contribute to this effect. We also tried to investigate the effect using transwell cocultures, but unexpectedly there was no change in the number of regulatory T cells. We therefore think this effect may also depend on direct cell-cell contact. We will further investigate the other mechanisms involved the effect of Aire overexpression in our future study. In summary, Aire-overexpressing DCs maintain CD4
+ T cell tolerance by inducing the functional inactivation and apoptosis of autoreactive CD4
+ T cells.
Other studies have reported that T1D is caused by autoreactive T cells. Once activated, autoreactive T cells enter into pancreatic tissues, form inflammatory infiltrates and produce large amounts of IFN-γ, which leads to the further destruction of pancreatic β cells. Eventually, pancreatic β cells lose the ability to secrete insulin [
40,
41]. In a previous study, Aire was transfected into bone marrow cells. Aire was shown to promote the expression of TRAs in bone marrow cells, such as myelin oligodendrocyte glycoprotein (Mog). Moreover, adoptive transfer of Aire-overexpressing bone marrow cells delays the occurrence of experimental autoimmune encephalomyelitis (EAE) in Mog-induced mice [
21]. Previously, we demonstrated that DCs transfected with Aire induced the inactivation of CD4
+ T cells using
in vitro assays. To investigate whether CD4
+ T cells were capable of resisting activation after the induction of tolerance and truly played a role in preventing the occurrence of T1D, splenocytes derived from WT mice were
co-cultured with Aire-overexpressing DCs and then adoptively transferred into WT mice. Then, the mice were induced with STZ. The blood glucose levels were significantly reduced in mice adoptively transferred with Aire-overexpressing DC-induced splenocytes. Furthermore, H&E staining of mouse pancreatic tissues showed a low degree of inflammatory infiltration. The IFN-γ and p-EPK expression levels were markedly reduced in CD4
+ T cells among the total splenocytes following insulin stimulation. These results indicated that CD4
+ T cells induced with Aire-overexpressing DCs were capable of delaying the occurrence of T1D in STZ-induced mice.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




