
Downregulation of Runx2 by 1,25-Dihydroxyvitamin D3 Induces the Transdifferentiation of Osteoblasts to Adipocytes

Abstract
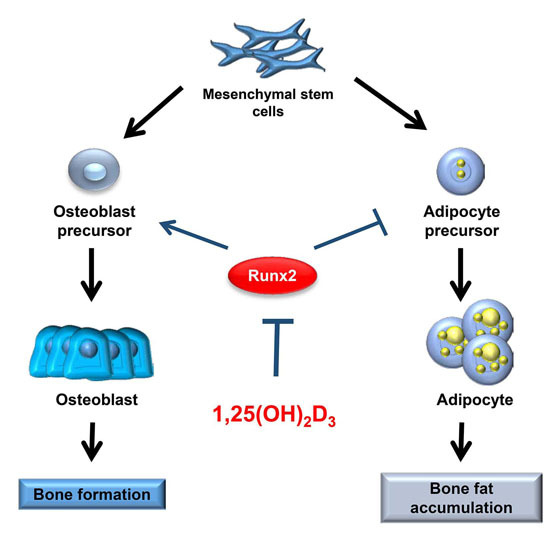
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
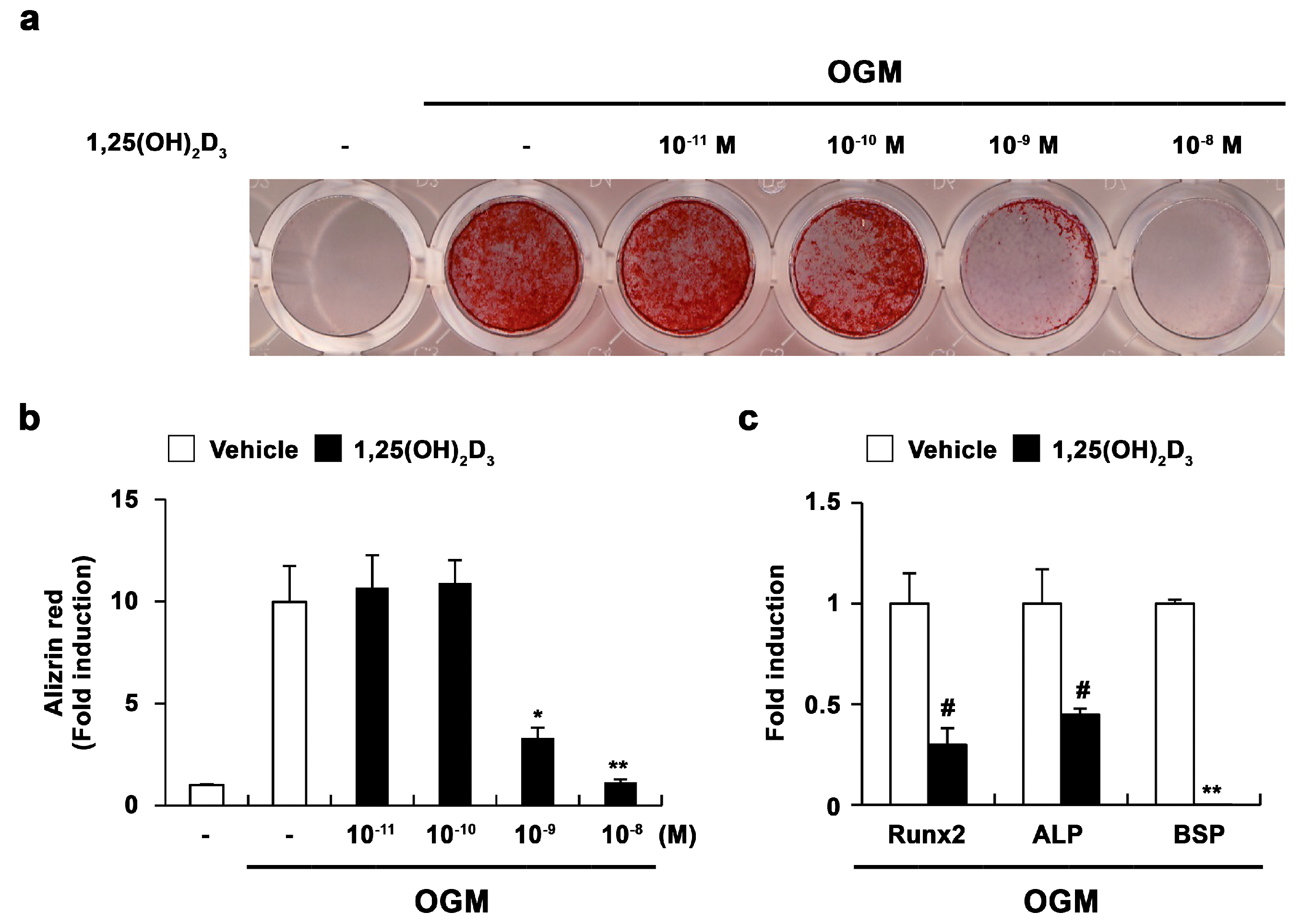
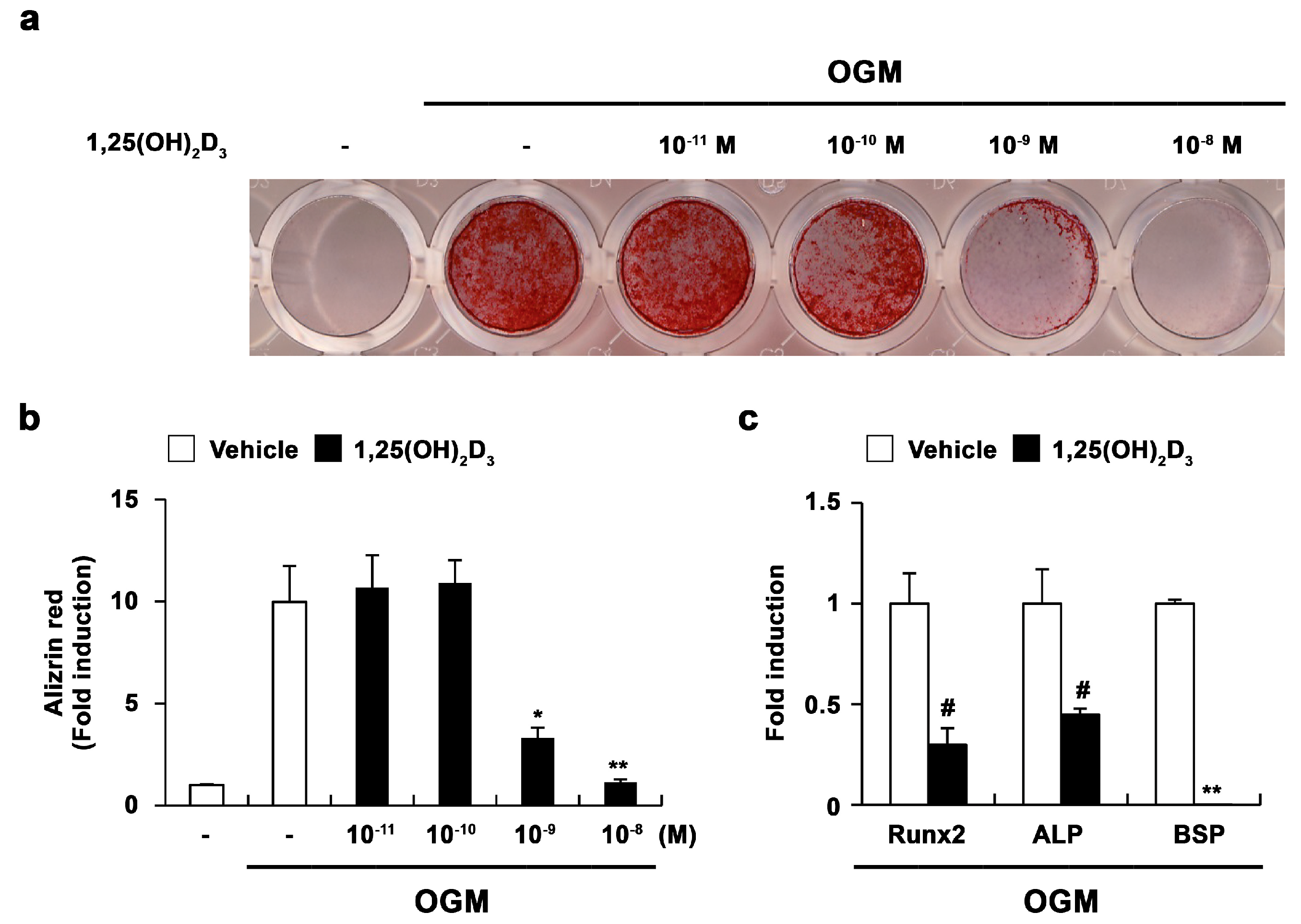
2.1. 1,25-Dihydroxyvitamin D3 (1,25(OH)2D3) Inhibits Osteoblast Differentiation and Runx2 Expression in Primary Osteoblasts
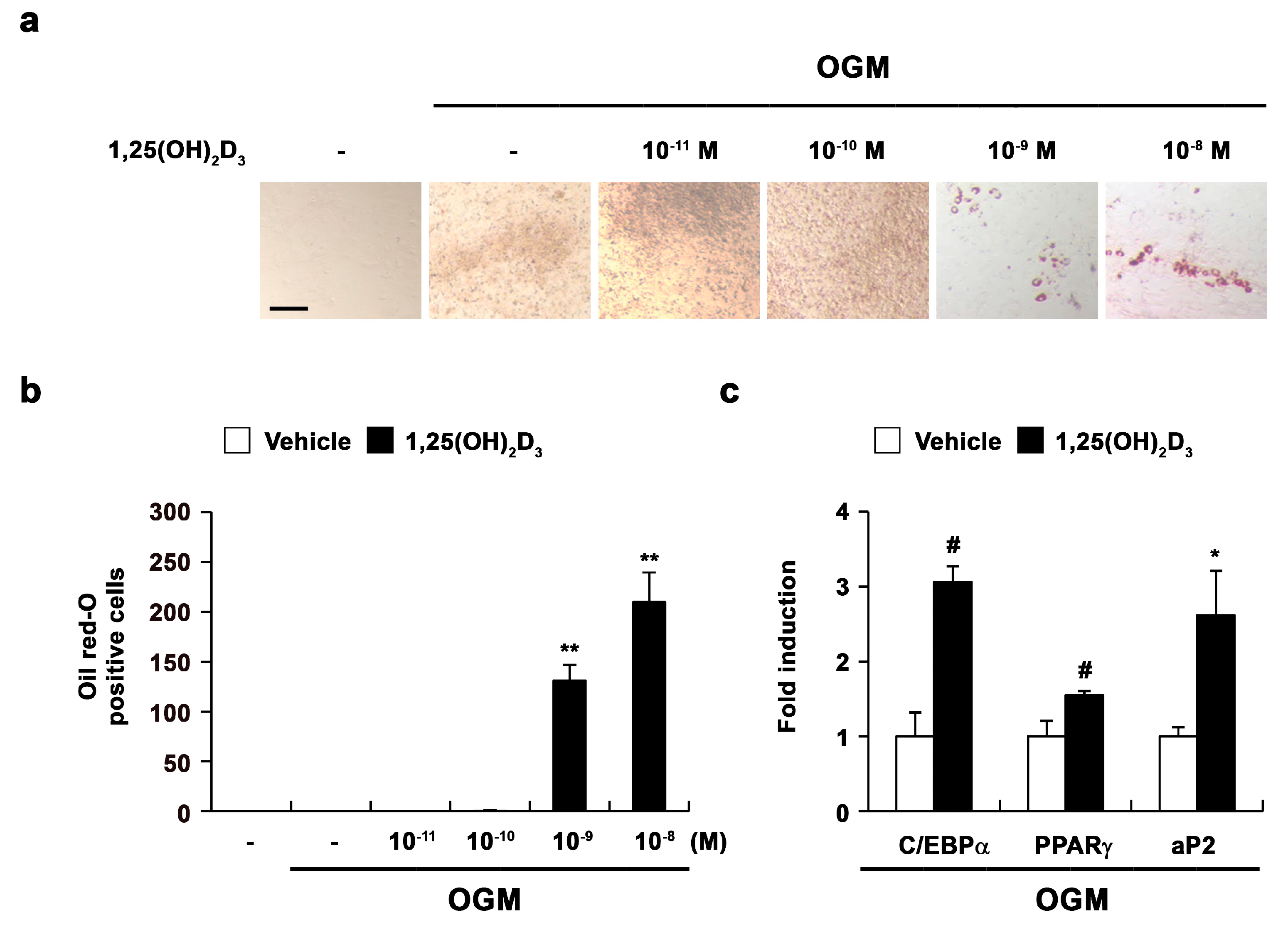
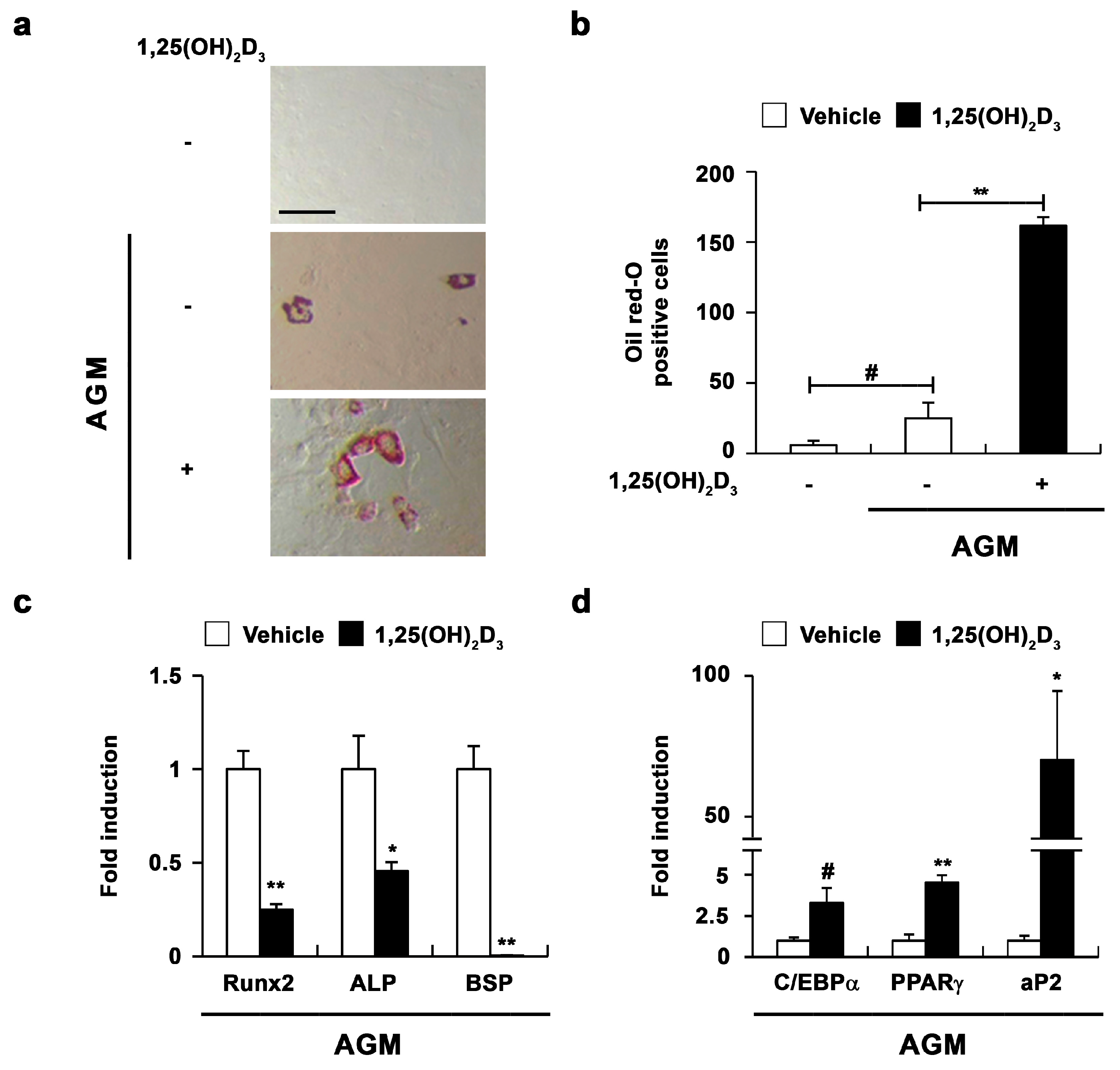
2.2. 1,25(OH)2D3 Induces the Transdifferentiation of Osteoblasts to Adipocytes
2.3. 1,25(OH)2D3 Induces Adipocyte Differentiation in Osteoblasts
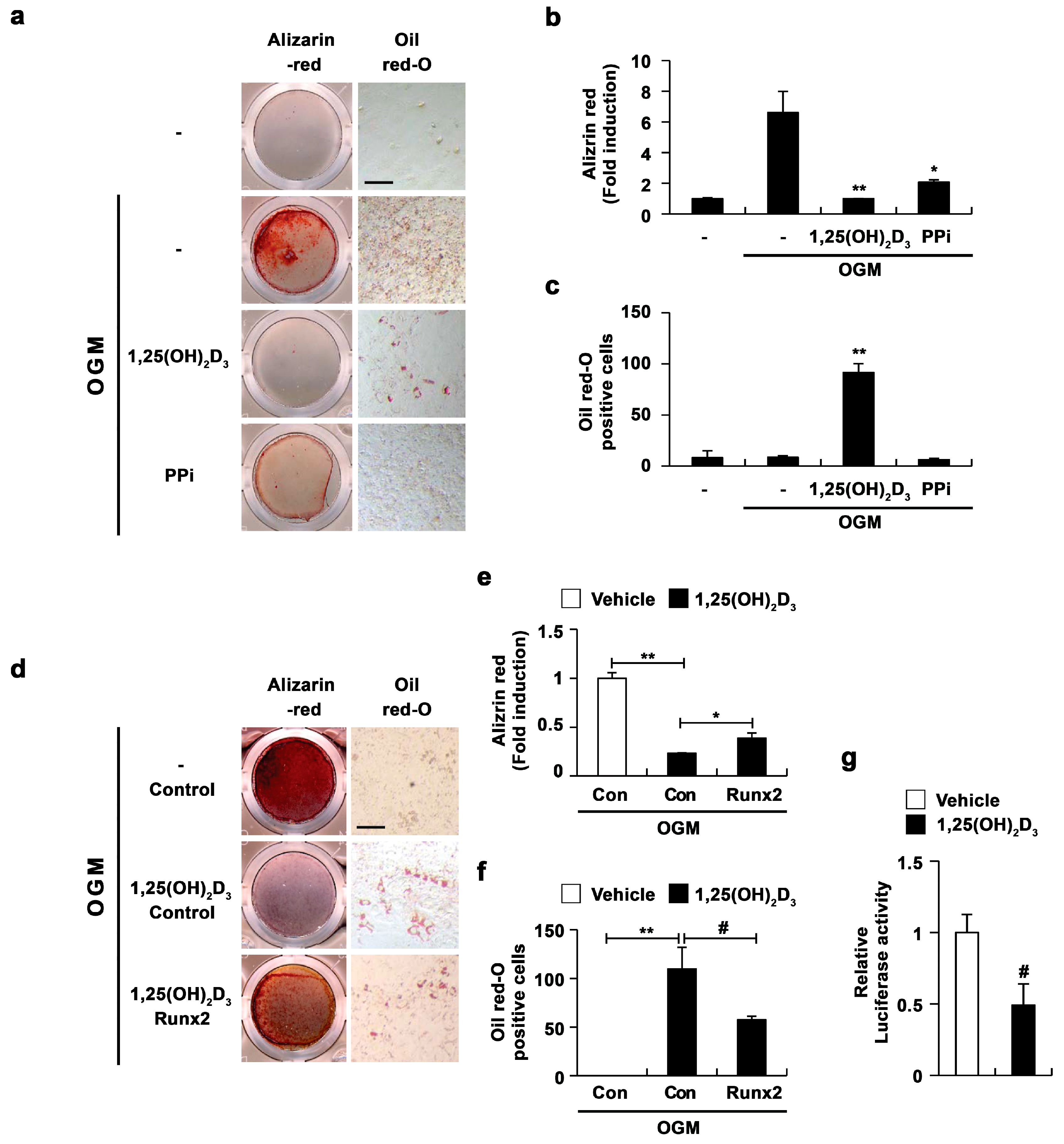
2.4. 1,25(OH)2D3 Induces the Transdifferentiation of Osteoblasts to Adipocytes via the Regulation of Runx2 Expression
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Osteoblast Differentiation
4.3. Adipocyte Differentiation
4.4. Real-Time Polymerase Chain Reaction (PCR)
4.5. Luciferase Assay
4.6. Retroviral Gene Transduction
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 1,25(OH)2D3 | 1,25-Dihydroxyvitamin D3 |
| ALP | Alkaline phosphatase |
| BSP | Bone sialoprotein |
| CEBP-α | CCAAT/enhancer binding protein-α |
| PPAR-γ | Peroxisome proliferator-activated receptor-γ |
| PPi | Pyrophosphate |
| VDR | Vitamin D receptor |
| VDRE | Vitamin D response element |
References
- Walsh, M.C.; Kim, N.; Kadono, Y.; Rho, J.; Lee, S.Y.; Lorenzo, J.; Choi, Y. Osteoimmunology: Interplay between the immune system and bone metabolism. Annu. Rev. Immunol. 2006, 24, 33–63. [Google Scholar] [CrossRef] [PubMed]
- Cappariello, A.; Maurizi, A.; Veeriah, V.; Teti, A. The great beauty of the osteoclast. Arch. Biochem. Biophys. 2014, 558, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, N. Signaling pathways in osteoclast differentiation. Chonnam Med. J. 2016, 52, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Suda, T. Vitamin D: Calcium and bone homeostasis during evolution. BoneKEy Rep. 2014, 3, 480. [Google Scholar] [CrossRef] [PubMed]
- Lips, P.; van Schoor, N.M. The effect of vitamin D on bone and osteoporosis. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Lips, P. Vitamin D physiology. Prog. Biophys. Mol. Biol. 2006, 92, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Beresford, J.N.; Gallagher, J.A.; Russell, R.G. 1,25-Dihydroxyvitamin D3 and human bone-derived cells in vitro: Effects on alkaline phosphatase, type I collagen and proliferation. Endocrinology 1986, 119, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.P.; Lee, N.; Wang, K.C.; Soong, Y.K.; Huang, K.E. Effect of estrogen and 1α,25(OH)2-vitamin D3 on the activity and growth of human primary osteoblast-like cells in vitro. Fertil. Steril. 2002, 77, 1038–1043. [Google Scholar] [CrossRef]
- van Driel, M.; Koedam, M.; Buurman, C.J.; Hewison, M.; Chiba, H.; Uitterlinden, A.G.; Pols, H.A.; van Leeuwen, J.P. Evidence for auto/paracrine actions of vitamin D in bone: 1α-Hydroxylase expression and activity in human bone cells. FASEB J. 2006, 20, 2417–2419. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ducy, P.; Karsenty, G. 1,25-Dihydroxyvitamin D3 inhibits osteocalcin expression in mouse through an indirect mechanism. J. Biol. Chem. 1997, 272, 110–116. [Google Scholar] [PubMed]
- Lieben, L.; Masuyama, R.; Torrekens, S.; Van Looveren, R.; Schrooten, J.; Baatsen, P.; Lafage-Proust, M.H.; Dresselaers, T.; Feng, J.Q.; Bonewald, L.F.; et al. Normocalcemia is maintained in mice under conditions of calcium malabsorption by vitamin D-induced inhibition of bone mineralization. J. Clin. Investig. 2012, 122, 1803–1815. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of osteoblast differentiation by transcription factors. J. Cell. Biochem. 2006, 99, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted disruption of CBFA1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef]
- Otto, F.; Thornell, A.P.; Crompton, T.; Denzel, A.; Gilmour, K.C.; Rosewell, I.R.; Stamp, G.W.; Beddington, R.S.; Mundlos, S.; Olsen, B.R.; et al. CBFA1, a candidate gene for cleidocranial dysplasia syndrome, is essential for osteoblast differentiation and bone development. Cell 1997, 89, 765–771. [Google Scholar] [CrossRef]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBP α and PPAR γ controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef]
- Savopoulos, C.; Dokos, C.; Kaiafa, G.; Hatzitolios, A. Adipogenesis and osteoblastogenesis: Trans-differentiation in the pathophysiology of bone disorders. Hippokratia 2011, 15, 18–21. [Google Scholar] [PubMed]
- Ryan, K.J.; Daniel, Z.C.; Craggs, L.J.; Parr, T.; Brameld, J.M. Dose-dependent effects of vitamin D on transdifferentiation of skeletal muscle cells to adipose cells. J. Endocrinol. 2013, 217, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Drissi, H.; Pouliot, A.; Koolloos, C.; Stein, J.L.; Lian, J.B.; Stein, G.S.; van Wijnen, A.J. 1,25-(OH)2-vitamin D3 suppresses the bone-related Runx2/CBFA1 gene promoter. Exp. Cell Res. 2002, 274, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Han, M.S.; Che, X.; Cho, G.H.; Park, H.R.; Lim, K.E.; Park, N.R.; Jin, J.S.; Jung, Y.K.; Jeong, J.H.; Lee, I.K.; et al. Functional cooperation between vitamin D receptor and Runx2 in vitamin D-induced vascular calcification. PLoS ONE 2013, 8, e83584. [Google Scholar] [CrossRef] [PubMed]
- Kamei, Y.; Kawada, T.; Kazuki, R.; Ono, T.; Kato, S.; Sugimoto, E. Vitamin D receptor gene expression is up-regulated by 1,25-dihydroxyvitamin D3 in 3T3-L1 preadipocytes. Biochem. Biophys. Res. Commun. 1993, 193, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Kawada, T.; Kamei, Y.; Sugimoto, E. The possibility of active form of vitamins A and D as suppressors on adipocyte development via ligand-dependent transcriptional regulators. Int. J. Obes. Relat. Metab. Disord. 1996, 20, S52–S57. [Google Scholar] [PubMed]
- Sato, M.; Hiragun, A. Demonstration of 1α,25-dihydroxyvitamin D3 receptor-like molecule in ST 13 and 3T3 L1 preadipocytes and its inhibitory effects on preadipocyte differentiation. J. Cell. Physiol. 1988, 135, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Nimitphong, H.; Holick, M.F.; Fried, S.K.; Lee, M.J. 25-hydroxyvitamin D3 and 1,25-dihydroxyvitamin D3 promote the differentiation of human subcutaneous preadipocytes. PLoS ONE 2012, 7, e52171. [Google Scholar] [CrossRef] [PubMed]
- Bellows, C.G.; Wang, Y.H.; Heersche, J.N.; Aubin, J.E. 1,25-dihydroxyvitamin D3 stimulates adipocyte differentiation in cultures of fetal rat calvaria cells: Comparison with the effects of dexamethasone. Endocrinology 1994, 134, 2221–2229. [Google Scholar] [PubMed]
- Komori, T. Requisite roles of Runx2 and CBFB in skeletal development. J. Bone Miner. Metab. 2003, 21, 193–197. [Google Scholar] [PubMed]
- Enomoto, H.; Furuichi, T.; Zanma, A.; Yamana, K.; Yoshida, C.; Sumitani, S.; Yamamoto, H.; Enomoto-Iwamoto, M.; Iwamoto, M.; Komori, T. Runx2 deficiency in chondrocytes causes adipogenic changes in vitro. J. Cell Sci. 2004, 117, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Gori, F.; Thomas, T.; Hicok, K.C.; Spelsberg, T.C.; Riggs, B.L. Differentiation of human marrow stromal precursor cells: Bone morphogenetic protein-2 increases OSF2/CBFA1, enhances osteoblast commitment, and inhibits late adipocyte maturation. J. Bone Miner. Res. 1999, 14, 1522–1535. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Suzuki, K.; Hozumi, A.; Goto, H.; Tomita, M.; Koseki, H.; Yamashita, S.; Osaki, M. Potentiation of osteoclastogenesis by adipogenic conversion of bone marrow-derived mesenchymal stem cells. Biomed. Res. 2014, 35, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Yoshizawa, T.; Nagai, Y.; Yamato, H.; Fukumoto, S.; Sekine, K.; Kato, S.; Matsumoto, T.; Fujita, T. Stimulation of osteoclast formation by 1,25-dihydroxyvitamin D requires its binding to vitamin D receptor (VDR) in osteoblastic cells: Studies using vdr knockout mice. Endocrinology 1999, 140, 1005–1008. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, R.; Kitazawa, S. Vitamin D3 augments osteoclastogenesis via vitamin D-responsive element of mouse rankl gene promoter. Biochem. Biophys. Res. Commun. 2002, 290, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.M.; Eslick, G.D.; Nowson, C.; Smith, C.; Bensoussan, A. Use of calcium or calcium in combination with vitamin D supplementation to prevent fractures and bone loss in people aged 50 years and older: A meta-analysis. Lancet 2007, 370, 657–666. [Google Scholar] [CrossRef]
- Sanders, K.M.; Stuart, A.L.; Williamson, E.J.; Simpson, J.A.; Kotowicz, M.A.; Young, D.; Nicholson, G.C. Annual high-dose oral vitamin D and falls and fractures in older women: A randomized controlled trial. JAMA 2010, 303, 1815–1822. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, K.; Kim, I.; Seong, S.; Kim, N. NRROS negatively regulates osteoclast differentiation by inhibiting RANKL-mediated NF-κB and reactive oxygen species pathways. Mol. Cell 2015, 38, 904–910. [Google Scholar]
- Kim, K.; Kim, J.H.; Kim, I.; Lee, J.; Seong, S.; Park, Y.W.; Kim, N. MicroRNA-26a regulates RANKL-induced osteoclast formation. Mol. Cell 2015, 38, 75–80. [Google Scholar]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Seong, S.; Kim, K.; Kim, I.; Jeong, B.-C.; Kim, N. Downregulation of Runx2 by 1,25-Dihydroxyvitamin D3 Induces the Transdifferentiation of Osteoblasts to Adipocytes. Int. J. Mol. Sci. 2016, 17, 770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050770
Kim JH, Seong S, Kim K, Kim I, Jeong B-C, Kim N. Downregulation of Runx2 by 1,25-Dihydroxyvitamin D3 Induces the Transdifferentiation of Osteoblasts to Adipocytes. International Journal of Molecular Sciences. 2016; 17(5):770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050770
Chicago/Turabian StyleKim, Jung Ha, Semun Seong, Kabsun Kim, Inyoung Kim, Byung-Chul Jeong, and Nacksung Kim. 2016. "Downregulation of Runx2 by 1,25-Dihydroxyvitamin D3 Induces the Transdifferentiation of Osteoblasts to Adipocytes" International Journal of Molecular Sciences 17, no. 5: 770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17050770