
Control of Appetite and Food Preference by NMDA Receptor and Its Co-Agonist d-Serine


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. The Importance of Appetite and Food Preference in Obesity
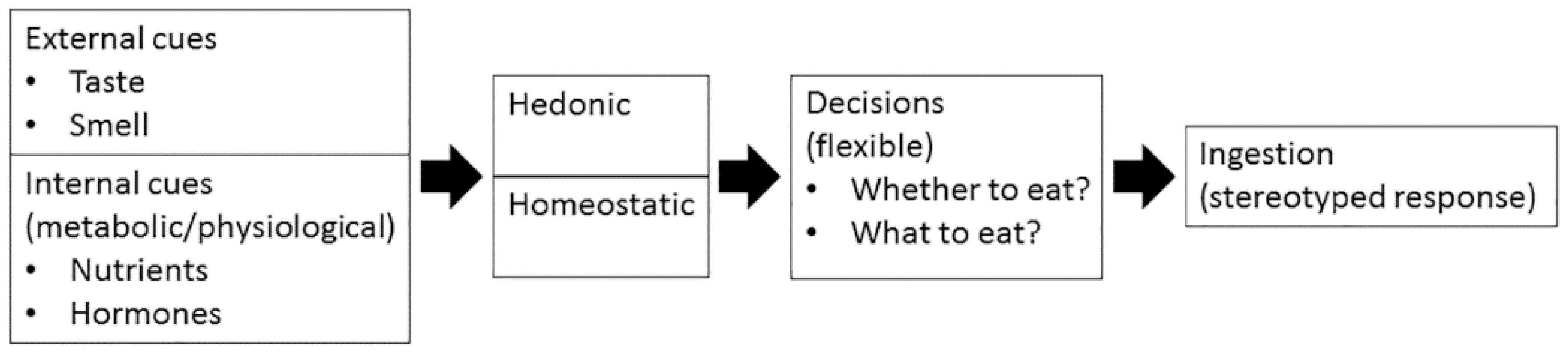
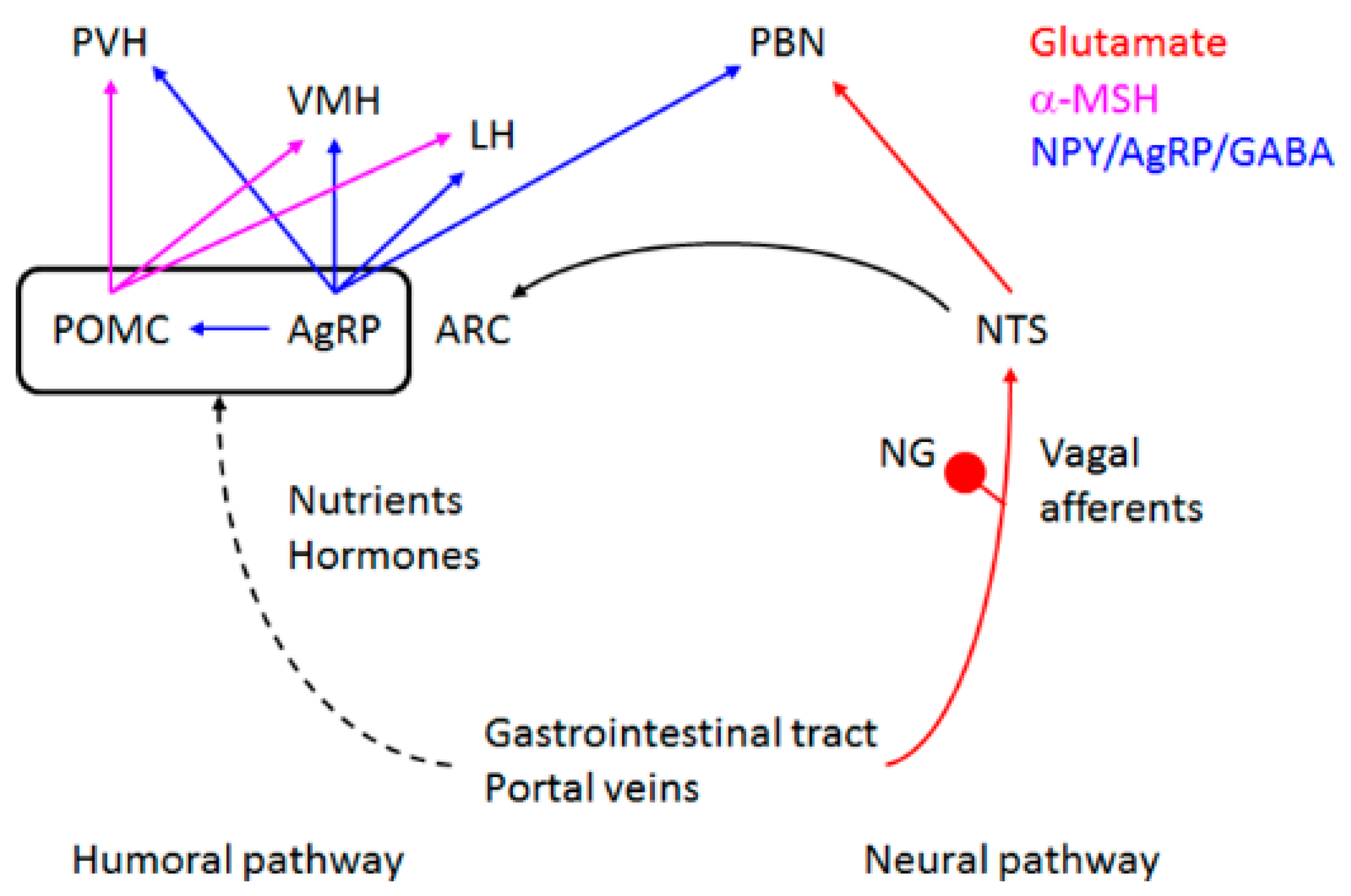
2. How Are Appetite and Food Preference Controlled?
3. NMDA Receptor and Its Co-Agonists Glycine and d-Serine
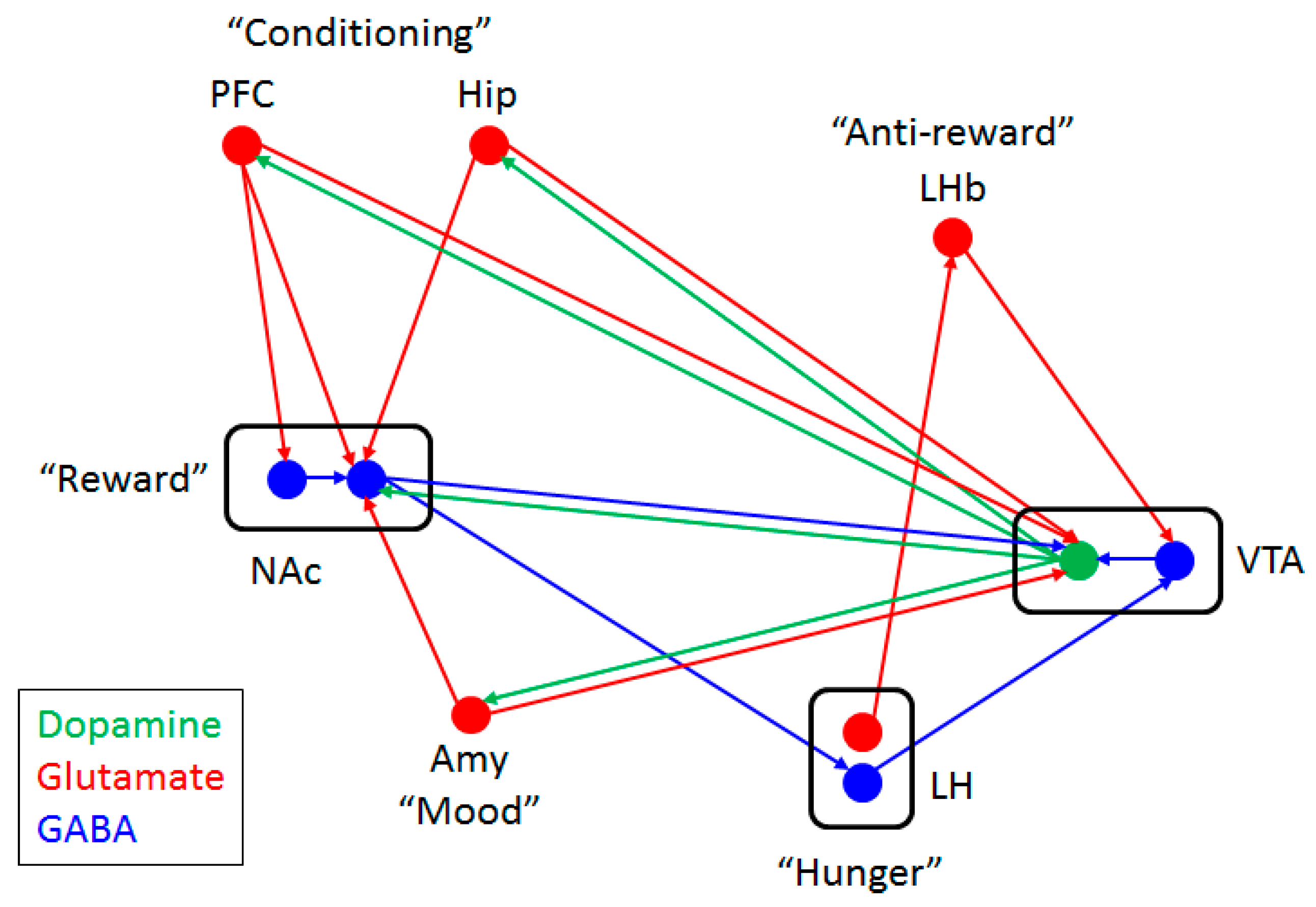
4. Glutamate-Dopamine Cross-Talk
5. Control of Appetite and Food Preference by NMDA Receptor and Its Co-Agonist d-Serine
6. Can d-Serine Be Used to Prevent Obesity?
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| NMDA receptor | N-methyl-d-aspartate receptor |
| WHO | World Health Organization |
| BMI | body mass index |
| POMC | proopiomelanocortin |
| AgRP | agouti-related peptide |
| α-MSH | alpha melanocyte-stimulating hormone |
| GABA | gamma amino butyric acid |
| VTA | ventral tegmental area |
| GLP-1 | glucagon-like peptide-1 |
| CSF | cerebrospinal fluid |
| SR | serine racemase |
| DAAO | d-amino acid oxidase |
| cAMP | 3′,5′-cyclic adenosine monophosphate |
| PKA | protein kinase A |
| DARPP-32 | dopamine- and cAMP-regulated phosphoprotein, molecular weight 32 kilo Dalton |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| HFD | high-fat diet |
| HSD | high-sucrose diet |
| HPD | high-protein diet |
References
- World Health Organization. Obesity and Overweight. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 15 February 2016).
- Lim, S.S.; Vos, T.; Flaxman, A.D.; Danaei, G.; Shibuya, K.; Adair-Rohani, H.; Amann, M.; Anderson, H.R.; Andrews, K.G.; Aryee, M.; et al. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: A systematic analysis for the global burden of disease study 2010. Lancet 2012, 380, 2224–2260. [Google Scholar] [CrossRef]
- Forouzanfar, M.H.; Alexander, L.; Anderson, H.R.; Bachman, V.F.; Biryukov, S.; Brauer, M.; Burnett, R.; Casey, D.; Coates, M.M.; Cohen, A.; et al. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks in 188 countries, 1990–2013: A systematic analysis for the global burden of disease study 2013. Lancet 2015, 386, 2287–2323. [Google Scholar] [CrossRef]
- Maskarinec, G.; Erber, E.; Grandinetti, A.; Verheus, M.; Oum, R.; Hopping, B.N.; Schmidt, M.M.; Uchida, A.; Juarez, D.T.; Hodges, K.; et al. Diabetes incidence based on linkages with health plans: The multiethnic cohort. Diabetes 2009, 58, 1732–1738. [Google Scholar] [CrossRef] [PubMed]
- Chiu, M.; Austin, P.C.; Manuel, D.G.; Shah, B.R.; Tu, J.V. Deriving ethnic-specific BMI cutoff points for assessing diabetes risk. Diabetes Care 2011, 34, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T. Age-associated weight gain, leptin, and sirt1: A possible role for hypothalamic SIRT1 in the prevention of weight gain and aging through modulation of leptin sensitivity. Front. Endocrinol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Kitamura, T. Roles of FOXO1 and SIRT1 in the central regulation of food intake. Endocr. J. 2010, 57, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Sims, E.A.; Goldman, R.F.; Gluck, C.M.; Horton, E.S.; Kelleher, P.C.; Rowe, D.W. Experimental obesity in man. Trans. Assoc. Am. Phys. 1968, 81, 153–170. [Google Scholar] [PubMed]
- The International Food Information Council Foundation. The 2015 Food & Health Survey: Consumer Attitudes toward Food Safety, Nutrition & Health; The International Food Information Council Foundation: Washington, DC, USA, 2015. [Google Scholar]
- Keen-Rhinehart, E.; Ondek, K.; Schneider, J.E. Neuroendocrine regulation of appetitive ingestive behavior. Front. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Basiri, M.L.; Stuber, G.D. Multimodal signal integration for feeding control. Cell 2016, 165, 522–523. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Robinson, T.E. Parsing reward. Trends Neurosci. 2003, 26, 507–513. [Google Scholar] [CrossRef]
- Palmiter, R.D. Is dopamine a physiologically relevant mediator of feeding behavior? Trends Neurosci. 2007, 30, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.J.; Hollon, N.G.; Phillips, P.E. Pavlovian valuation systems in learning and decision making. Curr. Opin. Neurobiol. 2012, 22, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M.; Jones-Gotman, M.; Dagher, A. Feeding-induced dopamine release in dorsal striatum correlates with meal pleasantness ratings in healthy human volunteers. Neuroimage 2003, 19, 1709–1715. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Telang, F. Overlapping neuronal circuits in addiction and obesity: Evidence of systems pathology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 3191–3200. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Kringelbach, M.L. Pleasure systems in the brain. Neuron 2015, 86, 646–664. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.S.; Berridge, K.C.; Aldridge, J.W. Disentangling pleasure from incentive salience and learning signals in brain reward circuitry. Proc. Natl. Acad. Sci. USA 2011, 108, E255–E264. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Globa, A.K.; Mills, F.; Naef, L.; Qiao, M.; Bamji, S.X.; Borgland, S.L. Consumption of palatable food primes food approach behavior by rapidly increasing synaptic density in the vta. Proc. Natl. Acad. Sci. USA 2016, 113, 2520–2525. [Google Scholar] [CrossRef] [PubMed]
- Norgren, R.; Hajnal, A.; Mungarndee, S.S. Gustatory reward and the nucleus accumbens. Physiol. Behav. 2006, 89, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Schultz, W. Predictive reward signal of dopamine neurons. J. Neurophysiol. 1998, 80, 1–27. [Google Scholar] [PubMed]
- Opland, D.M.; Leinninger, G.M.; Myers, M.G., Jr. Modulation of the mesolimbic dopamine system by leptin. Brain Res. 2010, 1350, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Alhadeff, A.L.; Rupprecht, L.E.; Hayes, M.R. GLP-1 neurons in the nucleus of the solitary tract project directly to the ventral tegmental area and nucleus accumbens to control for food intake. Endocrinology 2012, 153, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Abizaid, A.; Liu, Z.W.; Andrews, Z.B.; Shanabrough, M.; Borok, E.; Elsworth, J.D.; Roth, R.H.; Sleeman, M.W.; Picciotto, M.R.; Tschop, M.H.; et al. Ghrelin modulates the activity and synaptic input organization of midbrain dopamine neurons while promoting appetite. J. Clin. Investig. 2006, 116, 3229–3239. [Google Scholar] [CrossRef] [PubMed]
- Jerlhag, E.; Egecioglu, E.; Dickson, S.L.; Douhan, A.; Svensson, L.; Engel, J.A. Ghrelin administration into tegmental areas stimulates locomotor activity and increases extracellular concentration of dopamine in the nucleus accumbens. Addict. Biol. 2007, 12, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Figlewicz, D.P.; Evans, S.B.; Murphy, J.; Hoen, M.; Baskin, D.G. Expression of receptors for insulin and leptin in the ventral tegmental area/substantia nigra (VTA/SN) of the rat. Brain Res. 2003, 964, 107–115. [Google Scholar] [CrossRef]
- Leshan, R.L.; Opland, D.M.; Louis, G.W.; Leinninger, G.M.; Patterson, C.M.; Rhodes, C.J.; Munzberg, H.; Myers, M.G., Jr. Ventral tegmental area leptin receptor neurons specifically project to and regulate cocaine- and amphetamine-regulated transcript neurons of the extended central amygdala. J. Neurosci. 2010, 30, 5713–5723. [Google Scholar] [CrossRef] [PubMed]
- Fadel, J.; Deutch, A.Y. Anatomical substrates of orexin-dopamine interactions: Lateral hypothalamic projections to the ventral tegmental area. Neuroscience 2002, 111, 379–387. [Google Scholar] [CrossRef]
- Davis, J.F.; Choi, D.L.; Shurdak, J.D.; Krause, E.G.; Fitzgerald, M.F.; Lipton, J.W.; Sakai, R.R.; Benoit, S.C. Central melanocortins modulate mesocorticolimbic activity and food seeking behavior in the rat. Physiol. Behav. 2011, 102, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, I.; Horstmann, A.; Jurado, M.A.; Garolera, M.; Chaudhry, S.J.; Margulies, D.S.; Villringer, A.; Neumann, J. Reward processing in obesity, substance addiction and non-substance addiction. Obes. Rev. 2014, 15, 853–869. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Tomasi, D.; Baler, R.D. The addictive dimensionality of obesity. Biol. Psychiatry 2013, 73, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Stice, E.; Spoor, S.; Bohon, C.; Veldhuizen, M.G.; Small, D.M. Relation of reward from food intake and anticipated food intake to obesity: A functional magnetic resonance imaging study. J. Abnorm. Psychol. 2008, 117, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Stice, E.; Spoor, S.; Bohon, C.; Small, D.M. Relation between obesity and blunted striatal response to food is moderated by TaqIA A1 allele. Science 2008, 322, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, D.G. The glutamatergic nerve terminal. Eur. J. Biochem. 1993, 212, 613–631. [Google Scholar] [CrossRef] [PubMed]
- Niciu, M.J.; Kelmendi, B.; Sanacora, G. Overview of glutamatergic neurotransmission in the nervous system. Pharmacol. Biochem. Behav. 2012, 100, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Bliss, T.V.; Collingridge, G.L. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 1993, 361, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: Impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Cheriyan, J.; Mezes, C.; Zhou, N.; Balsara, R.D.; Castellino, F.J. Heteromerization of ligand binding domains of N-methyl-d-aspartate receptor requires both coagonists, l-glutamate and glycine. Biochemistry 2015, 54, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.W.; Ascher, P. Glycine potentiates the NMDA response in cultured mouse brain neurons. Nature 1987, 325, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Kleckner, N.W.; Dingledine, R. Requirement for glycine in activation of NMDA-receptors expressed in xenopus oocytes. Science 1988, 241, 835–837. [Google Scholar] [CrossRef] [PubMed]
- Mothet, J.P.; Parent, A.T.; Wolosker, H.; Brady, R.O., Jr.; Linden, D.J.; Ferris, C.D.; Rogawski, M.A.; Snyder, S.H. D-serine is an endogenous ligand for the glycine site of the N-methyl-d-aspartate receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 4926–4931. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.W. Native glycine receptor subtypes and their physiological roles. Neuropharmacology 2009, 56, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, Z.; Dai, Z.; Yang, Y.; Wang, J.; Wu, G. Glycine metabolism in animals and humans: Implications for nutrition and health. Amino Acids 2013, 45, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, A.; Nishikawa, T.; Hayashi, T.; Fujii, N.; Harada, K.; Oka, T.; Takahashi, K. The presence of free D-serine in rat brain. FEBS Lett. 1992, 296, 33–36. [Google Scholar] [CrossRef]
- Berger, A.J.; Dieudonne, S.; Ascher, P. Glycine uptake governs glycine site occupancy at NMDA receptors of excitatory synapses. J. Neurophysiol. 1998, 80, 3336–3340. [Google Scholar] [PubMed]
- Matsui, T.; Sekiguchi, M.; Hashimoto, A.; Tomita, U.; Nishikawa, T.; Wada, K. Functional comparison of d-serine and glycine in rodents: The effect on cloned NMDA receptors and the extracellular concentration. J. Neurochem. 1995, 65, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Papouin, T.; Ladepeche, L.; Ruel, J.; Sacchi, S.; Labasque, M.; Hanini, M.; Groc, L.; Pollegioni, L.; Mothet, J.P.; Oliet, S.H. Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell 2012, 150, 633–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, A.; Kumashiro, S.; Nishikawa, T.; Oka, T.; Takahashi, K.; Mito, T.; Takashima, S.; Doi, N.; Mizutani, Y.; Yamazaki, T.; et al. Embryonic development and postnatal changes in free d-aspartate and d-serine in the human prefrontal cortex. J. Neurochem. 1993, 61, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T. Analysis of free D-serine in mammals and its biological relevance. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 3169–3183. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.J.; Molliver, M.E.; Snyder, S.H. d-serine, an endogenous synaptic modulator: Localization to astrocytes and glutamate-stimulated release. Proc. Natl. Acad. Sci. USA 1995, 92, 3948–3952. [Google Scholar] [CrossRef] [PubMed]
- Kartvelishvily, E.; Shleper, M.; Balan, L.; Dumin, E.; Wolosker, H. Neuron-derived d-serine release provides a novel means to activate N-methyl-d-aspartate receptors. J. Biol. Chem. 2006, 281, 14151–14162. [Google Scholar] [CrossRef] [PubMed]
- Wolosker, H.; Sheth, K.N.; Takahashi, M.; Mothet, J.P.; Brady, R.O., Jr.; Ferris, C.D.; Snyder, S.H. Purification of serine racemase: Biosynthesis of the neuromodulator D-serine. Proc. Natl. Acad. Sci. USA 1999, 96, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Takayasu, N.; Hashimoto, A.; Sato, Y.; Tamaki, R.; Tsukamoto, H.; Kobayashi, H.; Noda, S. The serine racemase mRNA is predominantly expressed in rat brain neurons. Arch. Histol. Cytol. 2007, 70, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Weimer, W.R.; Neims, A.H. Hog cerebellar d-amino acid oxidase and its histochemical and immunofluorescent localization. J. Neurochem. 1977, 28, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.C.; Tsai, G.E.; Ma, C.L.; Ehmsen, J.T.; Mustafa, A.K.; Han, L.; Jiang, Z.I.; Benneyworth, M.A.; Froimowitz, M.P.; Lange, N.; et al. Targeted disruption of serine racemase affects glutamatergic neurotransmission and behavior. Mol. Psychiatry 2009, 14, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Horio, M.; Kohno, M.; Fujita, Y.; Ishima, T.; Inoue, R.; Mori, H.; Hashimoto, K. Levels of d-serine in the brain and peripheral organs of serine racemase (Srr) knock-out mice. Neurochem. Int. 2011, 59, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, Y.; Konno, R.; Sasabe, J.; Ueno, K.; Tojo, Y.; Mita, M.; Aiso, S.; Hamase, K. Alteration of intrinsic amounts of d-serine in the mice lacking serine racemase and d-amino acid oxidase. Amino Acids 2012, 43, 1919–1931. [Google Scholar] [CrossRef] [PubMed]
- Savignac, H.M.; Corona, G.; Mills, H.; Chen, L.; Spencer, J.P.; Tzortzis, G.; Burnet, P.W. Prebiotic feeding elevates central brain derived neurotrophic factor, N-methyl-d-aspartate receptor subunits and d-serine. Neurochem. Int. 2013, 63, 756–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardoni, F.; Bellone, C. Modulation of the glutamatergic transmission by dopamine: A focus on parkinson, huntington and addiction diseases. Front. Cell Neurosci. 2015, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Cepeda, C.; Buchwald, N.A.; Levine, M.S. Neuromodulatory actions of dopamine in the neostriatum are dependent upon the excitatory amino acid receptor subtypes activated. Proc. Natl. Acad. Sci. USA 1993, 90, 9576–9580. [Google Scholar] [CrossRef] [PubMed]
- Cepeda, C.; Levine, M.S. Dopamine and N-methyl-d-aspartate receptor interactions in the neostriatum. Dev. Neurosci. 1998, 20, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.S.; Li, Z.; Cepeda, C.; Cromwell, H.C.; Altemus, K.L. Neuromodulatory actions of dopamine on synaptically-evoked neostriatal responses in slices. Synapse 1996, 24, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Ladepeche, L.; Yang, L.; Bouchet, D.; Groc, L. Regulation of dopamine d1 receptor dynamics within the postsynaptic density of hippocampal glutamate synapses. PLoS ONE 2013, 8, e74512. [Google Scholar] [CrossRef] [PubMed]
- Snyder, G.L.; Fienberg, A.A.; Huganir, R.L.; Greengard, P. A dopamine/D1 receptor/protein kinase a/dopamine- and camp-regulated phosphoprotein (MR 32 kda)/protein phosphatase-1 pathway regulates dephosphorylation of the NMDA receptor. J. Neurosci. 1998, 18, 10297–10303. [Google Scholar] [PubMed]
- Snyder, G.L.; Allen, P.B.; Fienberg, A.A.; Valle, C.G.; Huganir, R.L.; Nairn, A.C.; Greengard, P. Regulation of phosphorylation of the GluR1 AMPA receptor in the neostriatum by dopamine and psychostimulants in vivo. J. Neurosci. 2000, 20, 4480–4488. [Google Scholar] [PubMed]
- Geisler, S.; Wise, R.A. Functional implications of glutamatergic projections to the ventral tegmental area. Rev. Neurosci. 2008, 19, 227–244. [Google Scholar] [CrossRef] [PubMed]
- Carr, D.B.; Sesack, S.R. Projections from the rat prefrontal cortex to the ventral tegmental area: Target specificity in the synaptic associations with mesoaccumbens and mesocortical neurons. J. Neurosci. 2000, 20, 3864–3873. [Google Scholar] [PubMed]
- D’Souza, M.S. Glutamatergic transmission in drug reward: Implications for drug addiction. Front. Neurosci. 2015, 9, 404. [Google Scholar] [CrossRef] [PubMed]
- Petrovich, G.D. Forebrain circuits and control of feeding by learned cues. Neurobiol. Learn. Mem. 2010, 95, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.J.; Nestler, E.J. The brain reward circuitry in mood disorders. Nat. Rev. Neurosci. 2013, 14, 609–625. [Google Scholar] [CrossRef] [PubMed]
- Guard, D.B.; Swartz, T.D.; Ritter, R.C.; Burns, G.A.; Covasa, M. NMDA NR2 receptors participate in CCK-induced reduction of food intake and hindbrain neuronal activation. Brain Res. 2009, 1266, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Ritter, R.C. A tale of two endings: Modulation of satiation by NMDA receptors on or near central and peripheral vagal afferent terminals. Physiol. Behav. 2011, 105, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Shoham, S.; Javitt, D.C.; Heresco-Levy, U. Chronic high-dose glycine nutrition: Effects on rat brain cell morphology. Biol. Psychiatry 2001, 49, 876–885. [Google Scholar] [CrossRef]
- Sorrels, T.L.; Bostock, E. Induction of feeding by 7-chlorokynurenic acid, a strychnine-insensitive glycine binding site antagonist. Brain Res. 1992, 572, 265–268. [Google Scholar] [CrossRef]
- Tejas-Juarez, J.G.; Cruz-Martinez, A.M.; Lopez-Alonso, V.E.; Garcia-Iglesias, B.; Mancilla-Diaz, J.M.; Floran-Garduno, B.; Escartin-Perez, R.E. Stimulation of dopamine D4 receptors in the paraventricular nucleus of the hypothalamus of male rats induces hyperphagia: Involvement of glutamate. Physiol. Behav. 2014, 133, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.A.; Ritter, R.C. NMDA-type glutamate receptors participate in reduction of food intake following hindbrain melanocortin receptor activation. Am. J. Physiol. Regul Integr. Comp. Physiol. 2015, 308, R1–R9. [Google Scholar] [CrossRef] [PubMed]
- Guard, D.B.; Swartz, T.D.; Ritter, R.C.; Burns, G.A.; Covasa, M. Blockade of hindbrain NMDA receptors containing NR2 subunits increases sucrose intake. Am. J. Physiol. Regul Integr. Comp. Physiol. 2009, 296, R921–R928. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.; Campos, C.; Herzog, T.; Covasa, M.; Czaja, K.; Ritter, R.C. Reduction of food intake by cholecystokinin requires activation of hindbrain NMDA-type glutamate receptors. Am. J. Physiol. Regul Integr. Comp. Physiol. 2011, 301, R448–R455. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Soden, M.E.; Zweifel, L.S.; Palmiter, R.D. Genetic identification of a neural circuit that suppresses appetite. Nature 2013, 503, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zheng, R.; Srisai, D.; McKnight, G.S.; Palmiter, R.D. NR2B subunit of the NMDA glutamate receptor regulates appetite in the parabrachial nucleus. Proc. Natl. Acad. Sci. USA 2013, 110, 14765–14770. [Google Scholar] [CrossRef] [PubMed]
- Resch, J.M.; Maunze, B.; Phillips, K.A.; Choi, S. Inhibition of food intake by pacap in the hypothalamic ventromedial nuclei is mediated by NMDA receptors. Physiol. Behav. 2014, 133, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.M.; Van Swieten, M.; Basiri, M.L.; Blair, G.A.; Kantak, P.; Stuber, G.D. Lateral hypothalamic area glutamatergic neurons and their projections to the lateral habenula regulate feeding and reward. J. Neurosci. 2016, 36, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Doane, D.F.; Lawson, M.A.; Meade, J.R.; Kotz, C.M.; Beverly, J.L. Orexin-induced feeding requires NMDA receptor activation in the perifornical region of the lateral hypothalamus. Am. J. Physiol. Regul Integr. Comp. Physiol. 2007, 293, R1022–R1026. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.M.; Curras, M.C.; Dao, J.; Jamal, F.A.; Turkowski, C.A.; Goel, R.K.; Gillard, E.R.; Wolfsohn, S.D.; Stanley, B.G. Lateral hypothalamic NMDA receptor subunits NR2A and/or NR2B mediate eating: Immunochemical/behavioral evidence. Am. J. Physiol. 1999, 276, R880–R891. [Google Scholar] [PubMed]
- Stanley, B.G.; Willett, V.L., III; Donias, H.W.; Dee, M.G., II; Duva, M.A. Lateral hypothalamic NMDA receptors and glutamate as physiological mediators of eating and weight control. Am. J. Physiol. 1996, 270, R443–R449. [Google Scholar] [PubMed]
- Sheng, Z.; Santiago, A.M.; Thomas, M.P.; Routh, V.H. Metabolic regulation of lateral hypothalamic glucose-inhibited orexin neurons may influence midbrain reward neurocircuitry. Mol. Cell Neurosci. 2014, 62, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Westerink, B.H.; Kwint, H.F.; de Vries, J.B. Eating-induced dopamine release from mesolimbic neurons is mediated by NMDA receptors in the ventral tegmental area: A dual-probe microdialysis study. J. Neurochem. 1997, 69, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Kinoshita, Y.; Matsui, S.; Kakuta, S.; Yokota-Hashimoto, H.; Kinoshita, K.; Iwasaki, Y.; Kinoshita, T.; Yada, T.; Amano, N.; et al. N-methyl-d-aspartate receptor coagonist d-serine suppresses intake of high-preference food. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R561–R575. [Google Scholar] [CrossRef] [PubMed]
- Balu, D.T.; Li, Y.; Puhl, M.D.; Benneyworth, M.A.; Basu, A.C.; Takagi, S.; Bolshakov, V.Y.; Coyle, J.T. Multiple risk pathways for schizophrenia converge in serine racemase knockout mice, a mouse model of NMDA receptor hypofunction. Proc. Natl. Acad. Sci. USA 2013, 110, E2400–E2409. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, A.; Chiba, S. Effect of systemic administration of d-serine on the levels of d- and l-serine in several brain areas and periphery of rat. Eur. J. Pharmacol. 2004, 495, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Labrie, V.; Wong, A.H.; Roder, J.C. Contributions of the d-serine pathway to schizophrenia. Neuropharmacology 2012, 62, 1484–1503. [Google Scholar] [CrossRef] [PubMed]
- Tsai, G.; Yang, P.; Chung, L.C.; Lange, N.; Coyle, J.T. d-serine added to antipsychotics for the treatment of schizophrenia. Biol. Psychiatry 1998, 44, 1081–1089. [Google Scholar] [CrossRef]
- Kantrowitz, J.T.; Malhotra, A.K.; Cornblatt, B.; Silipo, G.; Balla, A.; Suckow, R.F.; D’Souza, C.; Saksa, J.; Woods, S.W.; Javitt, D.C. High dose d-serine in the treatment of schizophrenia. Schizophr. Res. 2010, 121, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Levin, R.; Dor-Abarbanel, A.E.; Edelman, S.; Durrant, A.R.; Hashimoto, K.; Javitt, D.C.; Heresco-Levy, U. Behavioral and cognitive effects of the N-methyl-d-aspartate receptor co-agonist d-serine in healthy humans: Initial findings. J. Psychiatr. Res. 2015, 61, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Eto, S.; Yamaguchi, M.; Bounoshita, M.; Mizukoshi, T.; Miyano, H. High-throughput comprehensive analysis of d- and l-amino acids using ultra-high performance liquid chromatography with a circular dichroism (CD) detector and its application to food samples. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 3317–3325. [Google Scholar] [CrossRef] [PubMed]
- Casado, F.J.; Sanchez, A.H.; Rejano, L.; Montano, A. d-amino acid formation in sterilized alkali-treated olives. J. Agric. Food Chem. 2007, 55, 3503–3507. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T. Metabolism and functions of brain d-serine in mammals: Relevance to neuropsychiatric disorders. Seikagaku 2008, 80, 267–276. [Google Scholar] [PubMed]
- Kawai, M.; Sekine-Hayakawa, Y.; Okiyama, A.; Ninomiya, Y. Gustatory sensation of (l)- and (d)-amino acids in humans. Amino Acids 2012, 43, 2349–2358. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, Y.; Amadio, A.; Vincent, P.; Descheemaeker, A.; Oliet, S.H.; Dallel, R.; Voisin, D.L. Neuropathic pain depends upon d-serine co-activation of spinal NMDA receptors in rats. Neurosci. Lett. 2015, 603, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Sasabe, J.; Chiba, T.; Yamada, M.; Okamoto, K.; Nishimoto, I.; Matsuoka, M.; Aiso, S. d-serine is a key determinant of glutamate toxicity in amyotrophic lateral sclerosis. EMBO J. 2007, 26, 4149–4159. [Google Scholar] [CrossRef] [PubMed]
- Sasabe, J.; Miyoshi, Y.; Suzuki, M.; Mita, M.; Konno, R.; Matsuoka, M.; Hamase, K.; Aiso, S. d-amino acid oxidase controls motoneuron degeneration through d-serine. Proc. Natl. Acad. Sci. USA 2012, 109, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.; Paul, P.; Chen, H.J.; Morris, A.; Payling, M.; Falchi, M.; Habgood, J.; Panoutsou, S.; Winkler, S.; Tisato, V.; et al. Familial amyotrophic lateral sclerosis is associated with a mutation in d-amino acid oxidase. Proc. Natl. Acad. Sci. USA 2010, 107, 7556–7561. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; de Belleroche, J. The role of d-serine and glycine as co-agonists of NMDA receptors in motor neuron degeneration and amyotrophic lateral sclerosis (ALS). Front. Synaptic Neurosci. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Serafini, G.; Gonda, X.; Rihmer, Z.; Pompili, M.; Girardi, P.; Nasrallah, H.A.; Amore, M. NMDA receptor antagonists for depression: Critical considerations. Ann. Clin. Psychiatry 2015, 27, 213–220. [Google Scholar] [PubMed]
- Curcio, L.; Podda, M.V.; Leone, L.; Piacentini, R.; Mastrodonato, A.; Cappelletti, P.; Sacchi, S.; Pollegioni, L.; Grassi, C.; D'Ascenzo, M. Reduced d-serine levels in the nucleus accumbens of cocaine-treated rats hinder the induction of NMDA receptor-dependent synaptic plasticity. Brain 2013, 136, 1216–1230. [Google Scholar] [CrossRef] [PubMed]
- D’Ascenzo, M.; Podda, M.V.; Grassi, C. The role of d-serine as co-agonist of NMDA receptors in the nucleus accumbens: Relevance to cocaine addiction. Front. Synaptic Neurosci. 2014, 6. [Google Scholar] [CrossRef]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaki, T.; Matsui, S.; Kitamura, T. Control of Appetite and Food Preference by NMDA Receptor and Its Co-Agonist d-Serine. Int. J. Mol. Sci. 2016, 17, 1081. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071081
Sasaki T, Matsui S, Kitamura T. Control of Appetite and Food Preference by NMDA Receptor and Its Co-Agonist d-Serine. International Journal of Molecular Sciences. 2016; 17(7):1081. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071081
Chicago/Turabian StyleSasaki, Tsutomu, Sho Matsui, and Tadahiro Kitamura. 2016. "Control of Appetite and Food Preference by NMDA Receptor and Its Co-Agonist d-Serine" International Journal of Molecular Sciences 17, no. 7: 1081. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071081