
Extraordinary Adaptive Plasticity of Colorado Potato Beetle: “Ten-Striped Spearman” in the Era of Biotechnological Warfare


 ,
,
Abstract
:
{kind=link}
{kind=link}
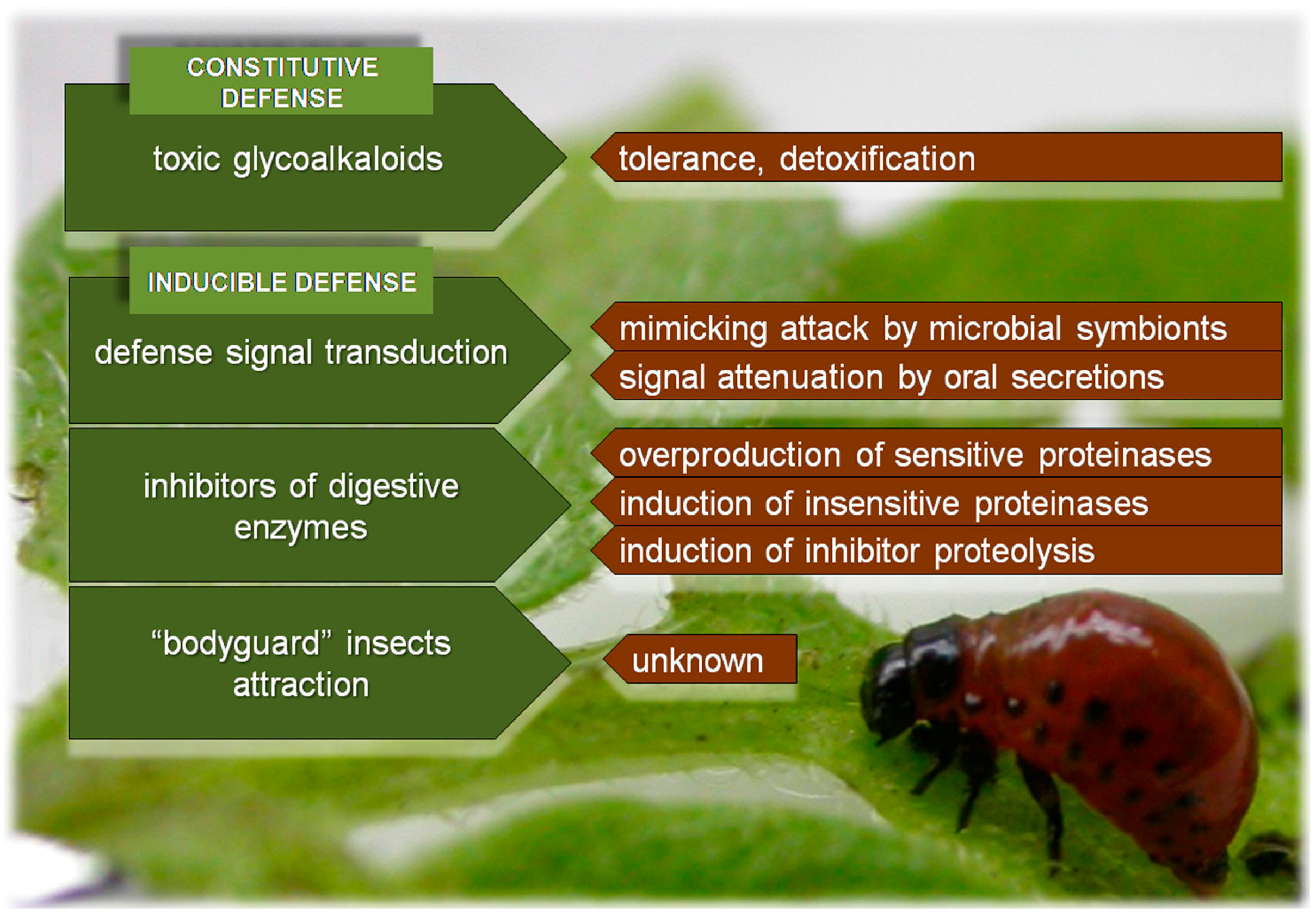{kind=link}
1. Introduction
2. From Innocuous Beetle to Superpest
3. Go behind Enemy Lines
4. One More in a Line
5. Compensating for Loss
6. Hopes for the Future?
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alyokhin, A.; Vincent, C.; Giordanengo, P. Insect Pests of Potato: Global Perspectives on Biology and Management, 1st ed.; Academic Press: Oxford, UK, 2013; p. 613. [Google Scholar]
- Hare, D.J. Impact of defoliation by the Colorado potato beetle on potato yields. J. Econ. Entomol. 1980, 73, 369–373. [Google Scholar] [CrossRef]
- Maharijaya, A.; Vosman, B. Managing the Colorado potato beetle; the need for resistance breeding. Euphytica 2015, 204, 487–501. [Google Scholar] [CrossRef]
- Casagrande, R.A. The Colorado potato beetle: 125 Years of mismanagement. Bull. Entomol. Soc. Am. 1987, 33, 142–150. [Google Scholar] [CrossRef]
- Johnson, C.G. International dispersal of insects and insect-born viruses. Eur. J. Plant Pathol. 1967, 73, 21–43. [Google Scholar]
- Grapputo, A.; Boman, S.; Lindström, L.; Lyytinen, A.; Mappes, J. The voyage of an invasive species across continents: Genetic diversity of North American and European Colorado potato beetle populations. Mol. Ecol. 2005, 14, 4207–4219. [Google Scholar] [CrossRef] [PubMed]
- Jolivet, P. The Colorado beetle menaces Asia. Leptinotarsa decemlineata Say 1824 Col. Chrysomelidae. Entomologiste 1991, 47, 29–48. [Google Scholar]
- Alyokhin, A.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. Colorado potato beetle resistance to insecticides. Am. J. Potato Res. 2008, 85, 395–413. [Google Scholar] [CrossRef]
- Agosta, S.J. On ecological fitting, plant-insect associations, herbivore host shifts, and host plant selection. Oikos 2006, 114, 556–565. [Google Scholar] [CrossRef]
- Izzo, V.M.; Mercer, N.; Armstrong, J.; Chen, Y.H. Variation in host usage among geographic populations of Leptinotarsa decemlineata, the Colorado potato beetle. J. Pest Sci. 2014, 87, 597–608. [Google Scholar] [CrossRef]
- Forister, M.L.; Ehmer, A.G.; Futuyma, D.J. The genetic architecture of a niche: Variation and covariation in host use traits in the Colorado potato beetle. J. Evol. Biol. 2007, 20, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Boman, S. Ecological and Genetic Factors Contributing to Invasion Success: The Northern Spread of the Colorado Potato Beetle (Leptinotarsa decemlineata); University of Jyväskylä: Jyväskylä, Finland, 2008. [Google Scholar]
- Weber, D. Colorado beetle: Pest on the move. Pestic. Outlook 2003, 14, 256–259. [Google Scholar] [CrossRef]
- Hazzard, R.V.; Ferro, D.N.; van Driesche, R.G.; Tuttle, A.F. Mortality of eggs of Colorado potato beetle (Coleoptera: Chrysomelidae) from predation by Coleomegilla maculate (Coleoptera: Coccinellidae). Environ. Entomol. 1991, 20, 841–848. [Google Scholar] [CrossRef]
- Biever, K.D.; Chauvin, R.L. Suppression of the Colorado potato beetle (Coleoptera: Chrysomelidae) with augmentative releases of predaceous stinkbugs (Hemiptera: Pentatomidae). J. Econ. Entomol. 1992, 85, 720–726. [Google Scholar] [CrossRef]
- Weber, D.C.; Rowley, D.L.; Greenstone, M.H.; Athanas, M.M. Prey preference and host suitability of the predatory and parasitoid carabid beetle, Lebia grandis, for several species of Leptinotarsa beetles. J. Insect Sci. 2006, 6, 9. [Google Scholar] [CrossRef]
- Jermy, T. The introduction of Perillus bioculatus into Europe to control the Colorado beetle. EPPO Bull. 1980, 10, 475–479. [Google Scholar] [CrossRef]
- Whalon, M.E.; Mota-Sanchez, D.; Hollingworth, R.M.; Gutierrez, R. Arthropod Pesticide Resistance Database, 2012. Available online: www.pesticideresistance.org (accessed on 1 June 2016).
- Whalon, M.E.; Mota-Sanchez, D.; Hollongworth, R.M. Analysis of global pesticide resistance in Arthropods. In Global Pesticide Resistance in Arthropods; Whalon, M.E., Mota-Sanchez, D., Hollongworth, R.M., Eds.; CAB International: London, UK, 2008; pp. 5–132. [Google Scholar]
- Forgash, A.J. Insecticide resistance in the Colorado potato beetle. In Proceedings of the Symposium on the Colorado Potato Beetle, XVII International Congress of Entomology, Hamburg, Germany, 20–26 August 1984; Ferro, D.N., Voss, R.H., Eds.; Massachusetts Experiment Station, College of Food and Natural Resources, University of Massachusetts at Amherst: Amherst, MA, USA, 1985; pp. 33–52. [Google Scholar]
- Olson, E.J.; Dively, G.P.; Nelson, J.O. Baseline susceptibility to imidacloprid and cross resistance patterns in Colorado potato beetle (Coleoptera: Chrysomelidae) populations. J. Econ. Entomol. 2000, 93, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-Y.; Bishop, B.A.; Grafius, E.J. Inheritance and synergism of resistance to imidacloprid in the Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2000, 93, 1508–1514. [Google Scholar] [CrossRef] [PubMed]
- Scott, I.M.; Tolman, J.H.; MacArthur, D.C. Insecticide resistance and cross-resistance development in Colorado potato beetle Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae) populations in Canada 2008–2011. Pest Manag. Sci. 2015, 71, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.J.; Jermy, T.; Butt, B.A. The influence of an alternate host plant on the fecundity of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Ann. Entomol. Soc. Am. 1980, 73, 197–199. [Google Scholar] [CrossRef]
- Alyokhin, A.V.; Ferro, D.N. Reproduction and dispersal of summer-generation Colorado potato beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 1999, 28, 425–430. [Google Scholar] [CrossRef]
- Labandeira, C.C. A paleobiologic perspective on plant–insect interactions. Curr. Opin. Plant Biol. 2013, 16, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Hare, J.D. How insect herbivores drive the evolution of plants. Science 2012, 338, 50–51. [Google Scholar] [CrossRef] [PubMed]
- Alyokhin, A.; Mota-Sanchez, D.; Baker, M.; Snyder, W.E.; Menasha, S.; Whalon, M.; Divelyf, G.; Moarsic, W.F. The Red Queen in a potato field: Integrated pest management versus chemical dependency in Colorado potato beetle control. Pest Manag. Sci. 2015, 71, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Schütz, S.; Klein, B.; Hummel, H.E. Host plant selection of the Colorado potato beetle as influenced by damaged induced volatiles of the potato plant. Naturwissenschaften 1997, 84, 212–217. [Google Scholar]
- Dickens, J.C. Behavioral response of larvae of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) to host plant volatile blends attractive to adults. Agric. For. Entomol. 2002, 4, 309–314. [Google Scholar] [CrossRef]
- Jermy, T. Hypotheses on oligophagy: How far the case of the Colorado potato beetle supports them. In Novel Aspects of the Biology of Chrysomelidae; Jolivet, P.H., Cox, M.L., Petitpierre, E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 127–139. [Google Scholar]
- Müller, C.; Renwick, J.A.A. Different phagostimulants in potato foliage for Manduca sexta and Leptinotarsa decemlineata. Chemoecology 2001, 11, 37–41. [Google Scholar] [CrossRef]
- Visser, J.H.; Ave, D.A. General green leaf volatiles in the olfactory orientation of the Colorado beetle, Leptinotarsa decemlineata. Entomol. Exp. Appl. 1978, 24, 738–749. [Google Scholar] [CrossRef]
- Bolter, J.C.; Dicke, M.; van Loon, J.J.A.; Visser, J.H.; Posthumus, M.A. Attraction of Colorado potato beetle to herbivore-damaged plants during herbivory and after its termination. J. Chem. Ecol. 1997, 23, 1003–1023. [Google Scholar] [CrossRef]
- Signoretti, A.G.C.; Peñaflor, M.F.G.V.; Bento, J.M.S. Fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), female moths respond to herbivore-induced corn volatiles. Neotrop. Entomol. 2012, 41, 22–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickens, J.C. Plant volatiles moderate response to aggregation pheromone in Colorado potato beetle. J. Appl. Entomol. 2006, 130, 26–31. [Google Scholar] [CrossRef]
- Hare, J.D. Growth of Leptinotarsa decemlineata larvae in response to simultaneous variation in protein and glycoalkaloid concentration. J. Chem. Ecol. 1987, 13, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Armer, C.A. Colorado potato beetle toxins revisited: Evidence the beetle does not sequester host plant glycoalkaloids. J. Chem. Ecol. 2004, 30, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Moural, T.W.; Nelson, D.R.; Palli, S.R. A specialist herbivore pest adaptation to xenobiotics through up-regulation of multiple Cytochrome P450s. Sci. Rep. 2016, 6, 20421. [Google Scholar] [CrossRef] [PubMed]
- Lyytinen, A.; Lindstrom, L.; Mappes, J.; Julkunen-Tiitto, R.; Fasulati, S.R.; Tiilikkala, K. Variability in host plant chemistry: Behavioural responses and life-history parameters of the Colorado potato beetle (Leptinotarsa decemlineata). Chemoecology 2007, 17, 51–56. [Google Scholar] [CrossRef]
- Heil, M.; Ibarra-Laclette, E.; Adame-Álvarez, R.M.; Martínez, O.; Ramirez-Chávez, E.; Molina-Torres, J.; Herrera-Estrella, L. How plants sense wounds: Damaged-self recognition is based on plant-derived elicitors and induces octadecanoid signaling. PLoS ONE 2012, 7, e30537. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, S.D.; Novak, N.G.; Ju, C.J.; Cooke, J.E. Potato, Solanum tuberosum, defense against Colorado potato beetle, Leptinotarsa decemlineata (Say): Microarray gene expression profiling of potato by Colorado potato beetle regurgitant treatment of wounded leaves. J. Chem. Ecol. 2008, 34, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Duceppe, M.-O.; Cloutier, C.; Michaud, D. Wounding, insect chewing and phloem sap feeding differentially alter the leaf proteome of potato, Solanum tuberosum L. Proteome Sci. 2012, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, S.D.; Novak, N.G.; Blackburn, M.B. Inhibition of proteinase inhibitor transcripts by Leptinotarsa decemlineata regurgitant in Solanum lycopersicum. J. Chem. Ecol. 2007, 33, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.H.; Rosa, C.; Scully, E.D.; Peiffer, M.; Tooker, J.F.; Hoover, K.; Luthe, D.S.; Felton, G.W. Herbivore exploits orally secreted bacteria to suppress plant defenses. Proc. Natl. Acad. Sci. USA 2013, 110, 15728–15733. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Baldwin, I.T. Independently silencing two photosynthetic proteins in Nicotiana attenuata has different effects on herbivore resistance. Plant Physiol. 2008, 148, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Bilgin, D.D.; Zavala, J.A.; Zhu, J.; Clough, S.J.; Ort, D.R.; DeLucia, E.H. Biotic stress globally downregulates photosynthesis genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef] [PubMed]
- Gruden, K.; Popovic, T.; Cimerman, N.; Krizaj, I.; Strukelj, B. Diverse enzymatic specificities of digestive proteases, “intestains”, enable Colorado potato beetle larvae to counteract the potato defence mechanism. Biol. Chem. 2003, 384, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Petek, M.; Turnšek, N.; Gašparič, M.B.; Novak, M.P.; Gruden, K.; Slapar, N.; Popovič, T.; Štrukelj, B.; Gruden, K.; Štrukelj, B.; et al. A complex of genes involved in adaptation of Leptinotarsa decemlineata larvae to induced potato defense. Arch. Insect Biochem. Physiol. 2012, 79, 153–181. [Google Scholar] [CrossRef] [PubMed]
- Ortego, F.; Novillo, C.; Sanchez-Serrano, J.J.; Castanera, P. Physiological response of Colorado potato beetle and beet armyworm larvae to depletion of wound-inducible proteinase inhibitors in transgenic potato plants. J. Insect Physiol. 2001, 47, 1291–1300. [Google Scholar] [CrossRef]
- Wegorek, W.; Dubniak, H. Reaction of potato leaves to the eggs of the Colorado beetle (Leptinotarsa decemlineata Say). Biul. Inst. Ochr. Rosl. 1972, 52, 409–413. [Google Scholar]
- Balbyshev, N.F.; Lorenzen, J.H. Hypersensitivity and egg drop: A novel mechanism of host plant resistance to Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1997, 90, 652–657. [Google Scholar] [CrossRef]
- NASS (National Agricultural Statistics Service), 2010, Acreage. Available online: http://usda.mannlib.cornell.edu/usda/nass/Acre/2010s/2010/Acre-06-30-2010.pdf (accessed on 17 May 2016).
- Mendelsohn, M.; Kough, J.; Vaituzis, Z.; Matthews, K. Are Bt crops safe? Nat. Biotechnol. 2003, 21, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Christou, P.; Capell, T.; Kohli, A.; Gatehouse, J.A.; Gatehouse, A.M. Recent developments and future prospects in insect pest control in transgenic crops. Trends Plant Sci. 2006, 11, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Perlak, F.J.; Stone, T.B.; Muskopf, Y.M.; Peterson, L.J.; Parker, G.B.; McPherson, S.A.; Wyman, J.; Love, S.; Reed, G.; Biever, D. Genetically improved potatoes: Protection from damage by Colorado potato beetles. Plant Mol. Biol. 1993, 22, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Gould, F.; Follett, P.; Nault, B.; Kennedy, G.G. Resistance management strategies for transgenic potato plants. In Advances in Potato Pest Biology and Management; Zehnder, G.W., Jansson, R.K., Powelson, M.L., Raman, K.V., Eds.; APS Press: St. Paul, MN, USA, 1994; pp. 255–277. [Google Scholar]
- Whalon, M.E.; Miller, D.L.; Hollingworth, R.M.; Grafius, E.J.; Miller, J.R. Selection of a Colorado potato beetle (Coleoptera: Chrysomelidae) strain resistant to Bacillus thuringiensis. J. Econ. Entomol. 1993, 86, 226–233. [Google Scholar] [CrossRef]
- Raharadja, U.; Whalon, M.E. Inheritance of resistance to Bacillus thuringiensis subsp. tenebrionis CryIIIA δ-endotoxin in Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1995, 88, 21–26. [Google Scholar]
- Whalon, M.E.; Ferro, D.N. Bt-potato resistance management. In Now or Never: Serious New Plans to Save a Natural Pest Control; Mellon, M., Rissler, J., Eds.; UCS: Cambridge, MA, USA, 1998; pp. 107–136. [Google Scholar]
- Loseva, O.; Ibrahim, M.; Candas, M.; Koller, C.N.; Bauer, L.S.; Bulla, L.A., Jr. Changes in protease activity and Cry3Aa toxin binding in the Colorado potato beetle: Implications for insect resistance to Bacillus thuringiensis toxins. Insect Biochem. Mol. Biol. 2002, 32, 567–577. [Google Scholar] [CrossRef]
- Hoffmann, J.A.; Reichhard, J.-M.; Hetru, C. Innate immunity in higher insects. Curr. Opin. Immunol. 1996, 8, 8–13. [Google Scholar] [CrossRef]
- Gunning, R.V.; Dang, H.T.; Kemp, F.C.; Nicholson, I.C.; Moores, G.D. New resistance mechanism in Helicoverpa armigera threatens transgenic crops expressing Bacillus thuringiensis Cry1Ac toxin. Appl. Environ. Microbiol. 2005, 71, 2558–2563. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Roberts, H.; Sarjan, M.; Featherstone, N.; Lahnstein, J.; Akhurst, R.; Schmidt, O. Is the mature endotoxin Cry1Ac from Bacillus thuringiensis inactivated by a coagulation reaction in the gut lumen of resistant Helicoverpa armigera larvae? Insect Biochem. Mol. Biol. 2005, 35, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Hoy, C.W.; Head, G. Correlation between behavioral and physiological responses to transgenic potatoes containing Bacillus thuringiensis delta-endotoxin in Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1995, 88, 480–486. [Google Scholar] [CrossRef]
- Alyokhin, A.V.; Ferro, D.N. Modifications in flight and oviposition of Bt-resistant and Bt-susceptible Colorado potato beetles as a result of exposure to Bacillus thuringiensis subsp. tenebrionis Cry3A toxin. Entomol. Exp. Appl. 1999, 90, 93–101. [Google Scholar] [CrossRef]
- Guo, W.; Wang, Z.; Luo, X.; Jin, X.; Chang, J.; He, J.; Tu, E.; Tian, Y.; Si, H.; Wu, J. Development of selectable marker-free transgenic potato plants expressing Cry3A against the Colorado potato beetle (Leptinotarsa decemlineata Say). Pest Manag. Sci. 2016, 72, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Hoy, C.W. Colorado potato beetle resistance management strategies for transgenic potatoes. Am. J. Potato Res. 1999, 76, 215–219. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Mickel, C.E.; Standish, J. Susceptibility of Processed Soy Flour and Soy Grits in Storage to Attack by Tribolium castaneum. Univ. Minn. Agric. Exp. Stn. Tech. Bull. 1947, 178, 1–20. [Google Scholar]
- Hilder, V.A.; Gatehouse, A.M.R.; Sheerman, S.E.; Baker, R.F.; Boulter, D. A novel mechanism of insect resistance engineered into tobacco. Nature 1987, 330, 160–163. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; Freire, M.G.M. Insect digestive enzymes as a target for pest control. Invertebr. Surviv. J. 2011, 8, 190–198. [Google Scholar]
- Broadway, R.M.; Duffey, S.S. Plant proteinase inhibitors: Mechanism of action and effect on the growth and digestive physiology of larval Heliothis zea and Spodoptera exiqua. J. Insect Physiol. 1986, 32, 827–833. [Google Scholar] [CrossRef]
- Ninković, S.; Miljuš-Đukić, J.; Lazarević, J. Proteinase inhibitors and the potential of their application in insect pest management. Ecology 2001, 36, 1–38. [Google Scholar]
- Zhu-Salzman, K.; Zeng, R. Insect response to plant defensive protease inhibitors. Annu. Rev. Entomol. 2015, 60, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Thie, N.M.R.; Houseman, J.G. Identification of cathepsin B, D, and H in the larval midgut of Colorado potato beetle, Leptinorarsa decemlineata Say (Coleoptera: Chrysomelidae). Insect Biochem. 1990, 20, 313–318. [Google Scholar] [CrossRef]
- Novillo, C.; Castanera, P.; Ortego, F. Characterisation and distribution of chymotrypsin and other digestive proteinases in Colorado potato beetle larvae. Arch. Insect Biochem. Physiol. 1997, 36, 181–201. [Google Scholar] [CrossRef]
- Bolter, C.J.; Latoszek-Green, M. Effect of chronic ingestion of the cysteine proteinase inhibitor, E-64, on Colorado potato beetle gut proteinases. Entomol. Exp. Appl. 1997, 83, 295–303. [Google Scholar] [CrossRef]
- Michaud, D.; Nguyen-Quoc, B.; Yelle, S. Selective inhibition of Colorado potato beetle cathepsin H by oryzacystatins I and II. FEBS Lett. 1993, 331, 173–176. [Google Scholar] [CrossRef]
- Leplé, J.-C.; Bonadé-Bottino, M.; Augustin, S.; Pilate, G.; Dumanois Lê Tân, V.; Delplanque, A.; Cornu, D.; Jouanin, L. Toxicity to Chrysomela tremulae (Coleoptera: Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor. Mol. Breed. 1995, 1, 319–328. [Google Scholar]
- Ninković, S.; Miljuš-Đukić, J.; Radović, S.; Maksimović, V.; Lazarević, J.; Vinterhalter, B.; Nešković, M.; Smigocki, A. Phytodecta fornicata Brüggemann resistance mediated by oryzacystatin II proteinase inhibitor transgene. Plant Cell Tissue Organ Cult. 2007, 91, 289–294. [Google Scholar] [CrossRef]
- Cloutier, C.; Fournier, M.; Jean, C.; Yelle, S.; Michaud, D. Growth compensation and faster development of Colorado potato beetle (Coleoptera: Chrysomelidae) feeding on potato foliage expressing oryzacystatin I. Arch. Insect Biochem. Physiol. 1999, 40, 69–79. [Google Scholar] [CrossRef]
- Cingel, A.; Savić, J.; Vinterhalter, B.; Vinterhalter, D.; Kostić, M.; Šešlija-Jovanović, D.; Smigocki, A.; Ninković, S. Growth and development of Colorado potato beetle larvae, Leptinotarsa decemlineata, on potato plants expressing the oryzacystatin II proteinase inhibitor. Transgenic Res. 2015, 24, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Cingel, A.; Savić, J.; Lazarević, J.; Ćosić, T.; Raspor, M.; Smigocki, A.; Ninković, S. Co-expression of the proteinase inhibitors oryzacystatin I and oryzacystatin II in transgenic potato alters Colorado potato beetle larval development. Insect Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Cingel, A.; Savić, J.; Ćosić, T.; Zdravković-Korać, S.; Momčilović, I.; Smigocki, A.; Ninković, S. Pyramiding rice cystatin OCI and OCII genes in transgenic potato (Solanum tuberosum L.) for resistance to Colorado potato beetle (Leptinotarsa decemlineata Say). Euphytica 2014, 198, 425–438. [Google Scholar] [CrossRef]
- Álvarez-Alfageme, F.; Martínez, M.; Pascual-Ruiz, S.; Castañera, P.; Diaz, I.; Ortego, F. Effects of potato plants expressing a barley cystatin on the predatory bug Podisus maculiventris via herbivorous prey feeding on the plant. Transgenic Res. 2007, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rasoolizadeh, A.; Goulet, M.-C.; Sainsbury, F.; Cloutier, C.; Michaud, D. Single substitutions to closely related amino acids contribute to the functional diversification of an insect-inducible, positively selected plant cystatin. FEBS J. 2016, 283, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Bown, D.P.; Wilkinson, H.S.; Gatehouse, J.A. Regulation of expression of genes encoding digestive proteases in the gut of a polyphagous lepidopteran larva in response to dietary protease inhibitors. Physiol. Entomol. 2004, 29, 278–290. [Google Scholar] [CrossRef]
- Spit, J.; Zels, S.; Dillen, S.; Holtof, M.; Wynant, N.; Vanden Broeck, J. Effects of different dietary conditions on the expression of trypsin and chymotrypsin-like protease genes in the digestive system of the migratory locust, Locusta migratoria. Insect Mol. Biol. 2014, 48, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Brunelle, F.; Nguyen-Quoc, B.; Cloutier, C.; Michaud, D. Protein hydrolysis by Colorado potato beetle (Leptinotarsa decemlineata Say: Chrysomelidae) digestive proteases: The catalytic role of cathepsin D. Arch. Insect Biochem. Physiol. 1999, 42, 88–98. [Google Scholar] [CrossRef]
- Brunelle, F.; Cloutier, C.; Michaud, D. Colorado potato beetles compensate for tomato cathepsin D inhibitor expressed in transgenic potato. Arch. Insect Biochem. Physiol. 2004, 55, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Bolter, C.J.; Jongsma, M.A. Colorado potato beetles (Leptinotarsa decemlineata) adapt to proteinase inhibitors induced in potato leaves by methyl jasmonate. J. Insect Physiol. 1995, 41, 1071–1078. [Google Scholar] [CrossRef]
- Overney, S.; Fawe, A.; Yelle, S.; Michaud, D. Diet-related plasticity of the digestive proteolytic system in larvae of the Colorado potato beetle (Leptinotarsa decemlineata Say). Arch. Insect Biochem. Physiol. 1997, 36, 241–250. [Google Scholar] [CrossRef]
- Rivard, D.; Cloutier, C.; Michaud, D. Colorado potato beetles show differential digestive compensatory responses to host plants expressing distinct sets of defense proteins. Arch. Insect Biochem. Physiol. 2004, 55, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Vorster, J.; Rasoolizadeh, A.; Goulet, M.-C.; Cloutier, C.; Sainsbury, F.; Michaud, D. Positive selection of digestive Cys proteases in herbivorous Coleoptera. Insect Biochem. Mol. Biol. 2015, 65, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Gruden, K.; Kuipers, A.G.; Guncar, G.; Slapar, N.; Strukelj, B.; Jongsma, M.A. Molecular basis of Colorado potato beetle adaptation to potato plant defence at the level of digestive cysteine proteinases. Insect Biochem. Mol. Biol. 2004, 34, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, F.; Rhéaume, A.-J.; Goulet, M.-C.; Vorster, J.; Michaud, D. Discrimination of differentially inhibited cysteine proteases by activity-based profiling using cystatin variants with tailored specificities. J. Proteome Res. 2012, 11, 5983–5993. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, F.; Benchabane, M.; Goulet, M.-C.; Michaud, D. Multimodal protein constructs for herbivore insect control. Toxins 2012, 4, 455–475. [Google Scholar] [CrossRef] [PubMed]
- Brunelle, F.; Girard, C.; Cloutier, C.; Michaud, D. A hybrid, broad-spectrum inhibitor of Colorado potato beetle aspartate and cysteine digestive proteinases. Insect Biochem. Physiol. 2005, 60, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Kutas, J.; Kondrák, M.; Szenthe, B.; Patthy, A.; Bánfalvi, Z.; Nádasy, M.; Gráf, L.; Asbóth, B. Colorado potato beetle larvae on potato plants expressing a locust proteinase inhibitor. Commun. Agric. Appl. Biol. Sci. 2004, 69, 281–287. [Google Scholar] [PubMed]
- Kondrák, M.; Kutas, J.; Szenthe, B.; Patthy, A.; Bánfalvi, Z.; Nádasy, M.; Gráf, L.; Asbóth, B. Inhibition of Colorado potato beetle larvae by a locust proteinase inhibitor peptide expressed in potato. Biotechnol. Lett. 2005, 27, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Outchkourov, N.S.; Rogelj, B.; Strukelj, B.; Jongsma, M.A. Expression of sea anemone equistatin in potato. Effects of plant proteases on heterologous protein production. Plant Physiol. 2003, 133, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Gruden, K.; Strukelj, B.; Popovic, T.; Lenarcic, B.; Bevec, T.; Brzin, J.; Kregar, I.; Herzog-Velikonja, J.; Stiekema, W.J.; Bosch, D.; et al. The cysteine protease activity of Colorado potato beetle (Leptinotarsa decemlineata Say) guts, which is insensitive to potato protease inhibitors, is inhibited by thyroglobulin type-1 domain inhibitors. Insect Biochem. Mol. Biol. 1998, 28, 549–560. [Google Scholar] [CrossRef]
- Šmid, I.; Gruden, K.; Buh Gašparič, M.; Koruza, K.; Petek, M.; Pohleven, J.; Brzin, J.; Kos, J.; Zel, J.; Sabotič, J. Inhibition of the growth of Colorado potato beetle larvae by macrocypins, protease inhibitors from the parasol mushroom. J. Agric. Food Chem. 2013, 61, 12499–12509. [Google Scholar] [CrossRef] [PubMed]
- Šmid, I.; Rotter, A.; Gruden, K.; Brzin, J.; Gašparič, M.B.; Kos, J.; Žel, J.; Sabotic, J. Clitocypin, a fungal cysteine protease inhibitor, exerts its insecticidal effect on Colorado potato beetle larvae by inhibiting their digestive cysteine proteases. Pestic. Biochem. Physiol. 2015, 122, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Abe, K. Cystatin-based control of insects, with special reference to oryzacystatin. In Recombinant Protease Inhibitors in Plants; Michaud, D., Ed.; Landes Bioscience: Georgetown, TX, USA, 2000; pp. 27–42. [Google Scholar]
- Franca, F.H.; Plaisted, R.L.; Roush, R.T.; Via, S.; Tingey, W.M. Selection response of the Colorado potato beetle for adaptation to the resistant potato, Solanum berthaultii. Entomol. Exp. Appl. 1994, 73, 101–109. [Google Scholar] [CrossRef]
- Voytas, D.F. Plant genome engineering with sequence-specific nucleases. Annu. Rev. Plant Biol. 2013, 64, 327–350. [Google Scholar] [CrossRef] [PubMed]
- The Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Congiu, L.; Lindström, L.; Piiroinen, S.; Vidotto, M.; Grapputo, A. Sequencing, de novo assembly and annotation of the Colorado potato beetle, Leptinotarsa decemlineata, transcriptome. PLoS ONE 2014, 9, e86012. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Liu, X.P.; Wan, P.J.; Shi, X.Q.; Guo, W.C.; Li, G.Q. The P450 enzyme shade mediates the hydroxylation of ecdysone to 20-hydroxyecdysone in the Colorado potato beetle, Leptinotarsa decemlineata. Insect Mol. Biol. 2014, 23, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.-Y.; Li, Q.; Zhou, L.-T.; Meng, Q.-W.; Lü, F.-G.; Guo, W.-C.; Li, G.-Q. Knockdown of juvenile hormone acidmethyl transferase severely affects the performance of Leptinotarsa decemlineata (Say) larvae and adults. Pest Manag. Sci. 2016, 72, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.B.; Ferro, D.N.; Porter, A.H. Invasions on large and small scales: Management of a well-established crop pest, the Colorado potato beetle. Biol. Invasions 2001, 3, 295–306. [Google Scholar] [CrossRef]
- Voss, R.H.; Ferro, D.N.; Logan, J.A. Role of reproductive diapause in the population dynamics of the Colorado potato beetle (Coleoptera: Chrysomelidae) in western Massachusetts. Environ. Entomol. 1988, 17, 863–871. [Google Scholar] [CrossRef]
- Matthews, D.L.; Michalak, P.S.; MacRae, R.J. The effect of traditional insect repellent plants on insect numbers in a mixed planting system. In Proceedings of the 4th International Federation of Organic Agricultural Movements Conference, Resource Conserving, Environmentally Sound Agricultural Alternatives, Cambridge, MA, USA, 18–20 August 1982.
- Martel, J.W.; Alford, A.R.; Dickens, J.C. Evaluation of a novel host plant volatile-based attracticide for management of Colorado potato beetle, Leptinotarsa decemlineata (Say). Crop Prot. 2007, 26, 822–827. [Google Scholar] [CrossRef]
- Brust, G.E. Natural enemies in straw-mulch reduce Colorado potato beetle populations and damage in potato. Biol. Control 1994, 4, 163–169. [Google Scholar] [CrossRef]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cingel, A.; Savić, J.; Lazarević, J.; Ćosić, T.; Raspor, M.; Smigocki, A.; Ninković, S. Extraordinary Adaptive Plasticity of Colorado Potato Beetle: “Ten-Striped Spearman” in the Era of Biotechnological Warfare. Int. J. Mol. Sci. 2016, 17, 1538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091538
Cingel A, Savić J, Lazarević J, Ćosić T, Raspor M, Smigocki A, Ninković S. Extraordinary Adaptive Plasticity of Colorado Potato Beetle: “Ten-Striped Spearman” in the Era of Biotechnological Warfare. International Journal of Molecular Sciences. 2016; 17(9):1538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091538
Chicago/Turabian StyleCingel, Aleksandar, Jelena Savić, Jelica Lazarević, Tatjana Ćosić, Martin Raspor, Ann Smigocki, and Slavica Ninković. 2016. "Extraordinary Adaptive Plasticity of Colorado Potato Beetle: “Ten-Striped Spearman” in the Era of Biotechnological Warfare" International Journal of Molecular Sciences 17, no. 9: 1538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091538