Transcriptome Sequencing Reveals the Differentially Expressed lncRNAs and mRNAs Involved in Cryoinjuries in Frozen-Thawed Giant Panda (Ailuropoda melanoleuca) Sperm


 ,
,  ,
,
Abstract
:
1. Introduction
2. Result
2.1. Sperm Quality before and after Cryopreservation
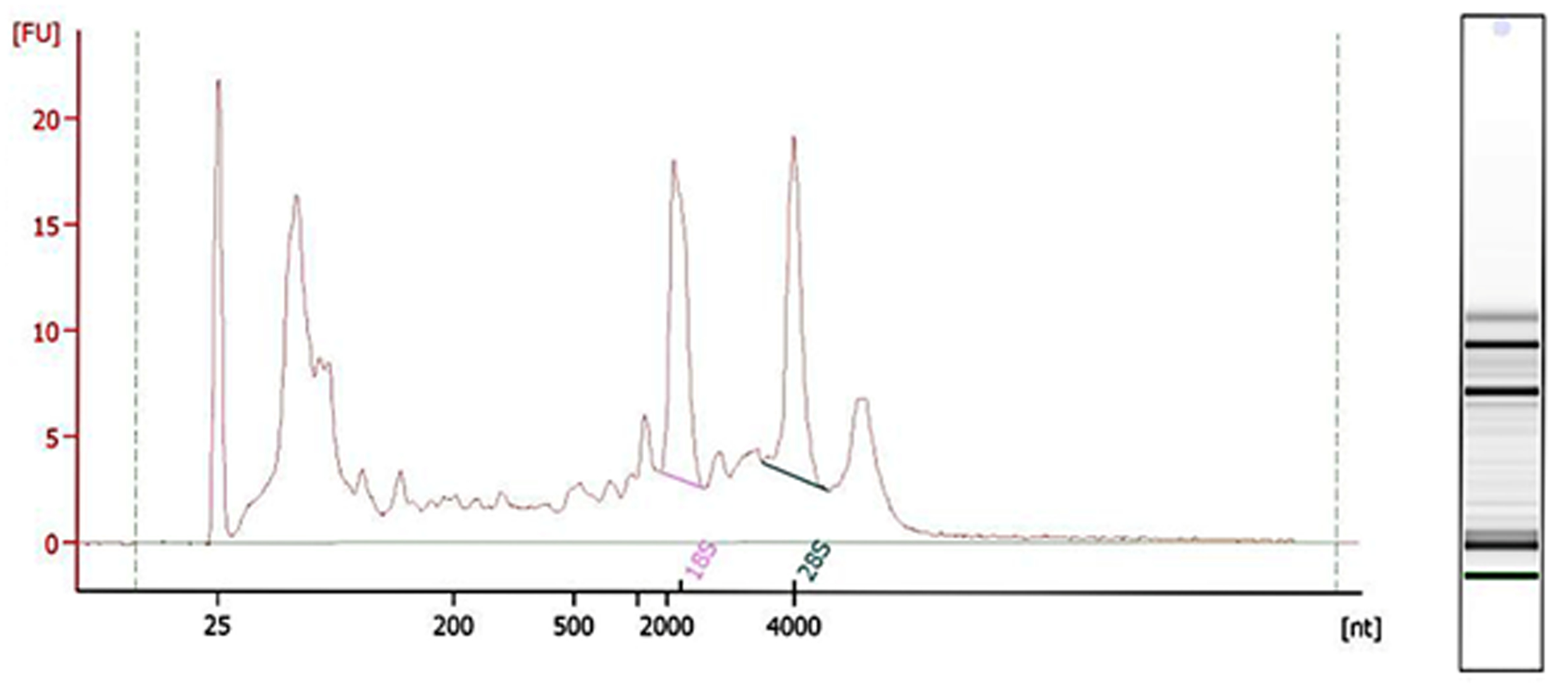
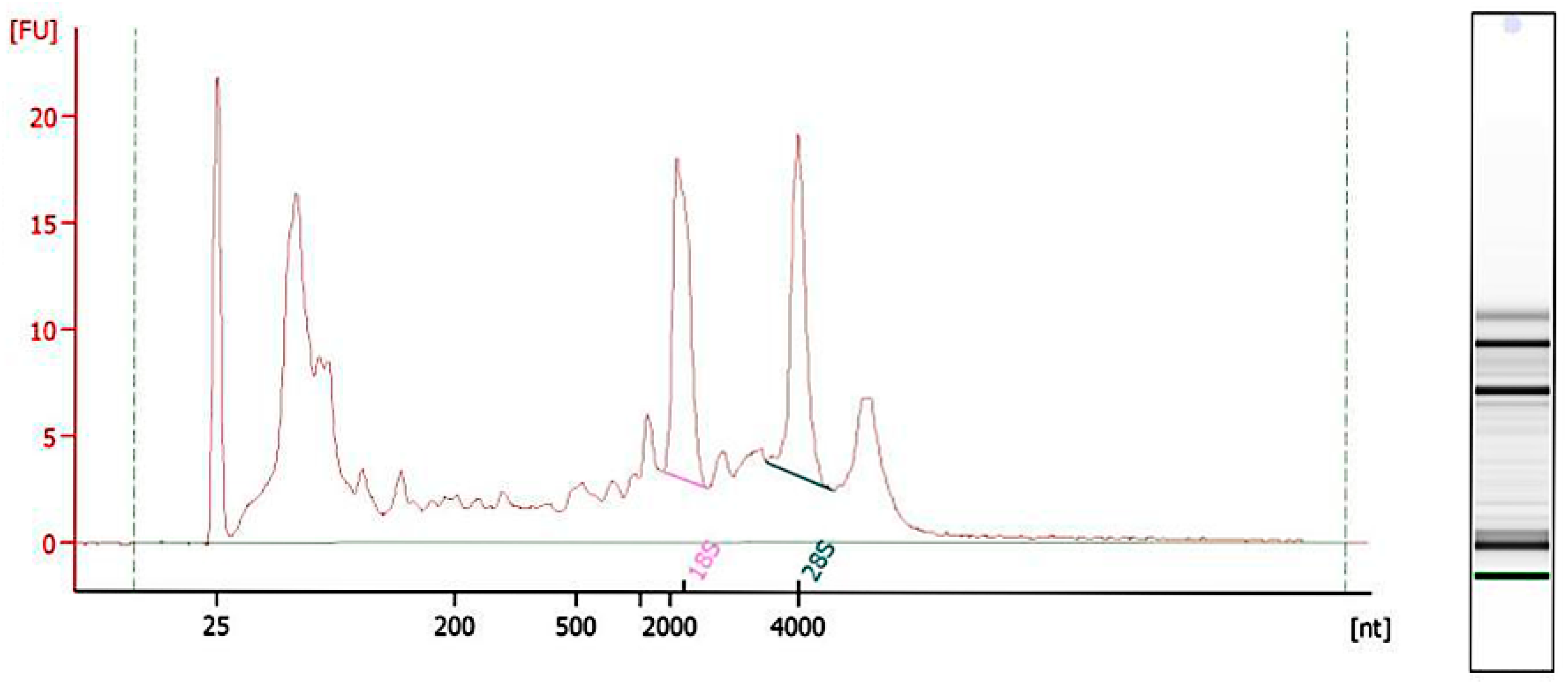
2.2. RNA Quality Inspection
2.3. RNA Sequencing Roundup
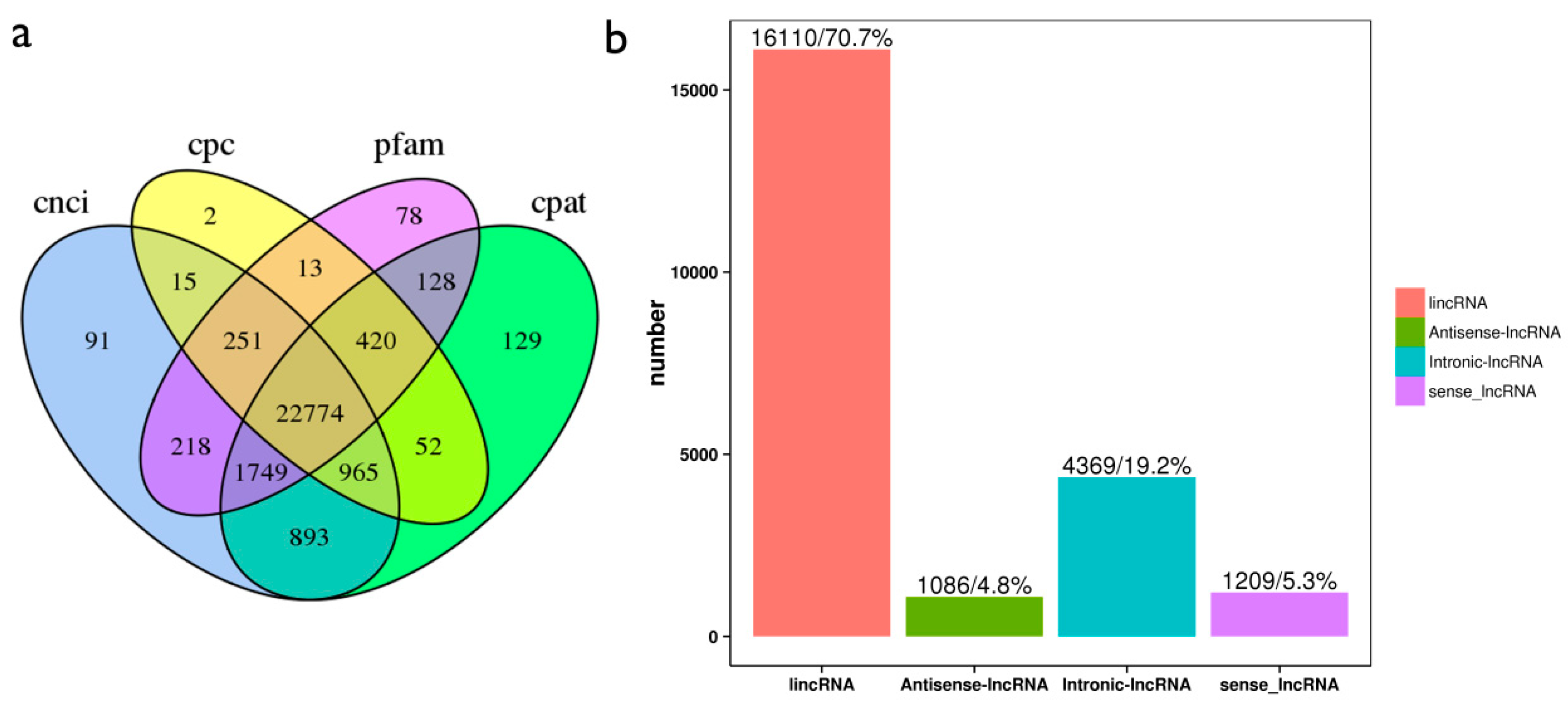
2.4. Identification of lncRNAs and mRNA
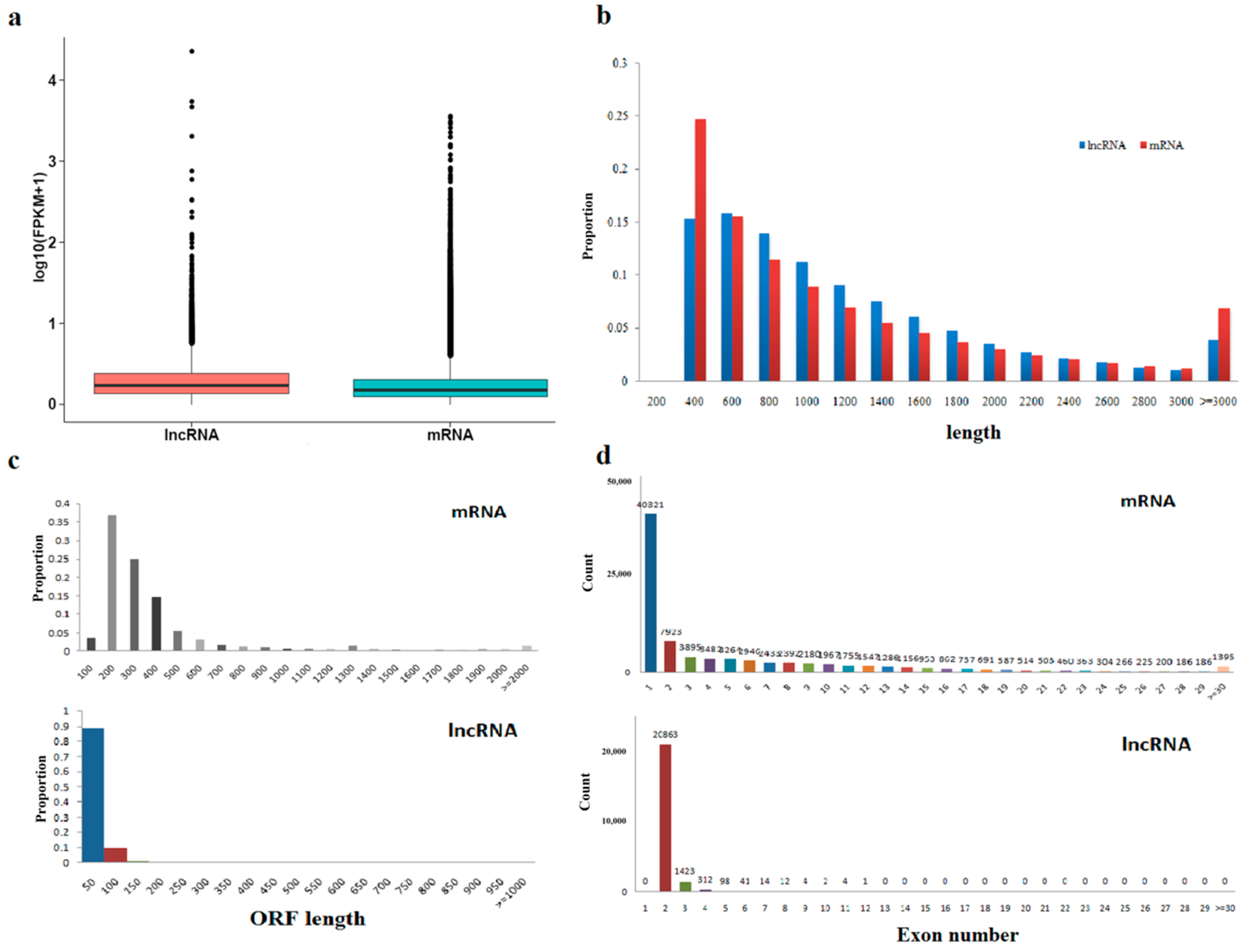
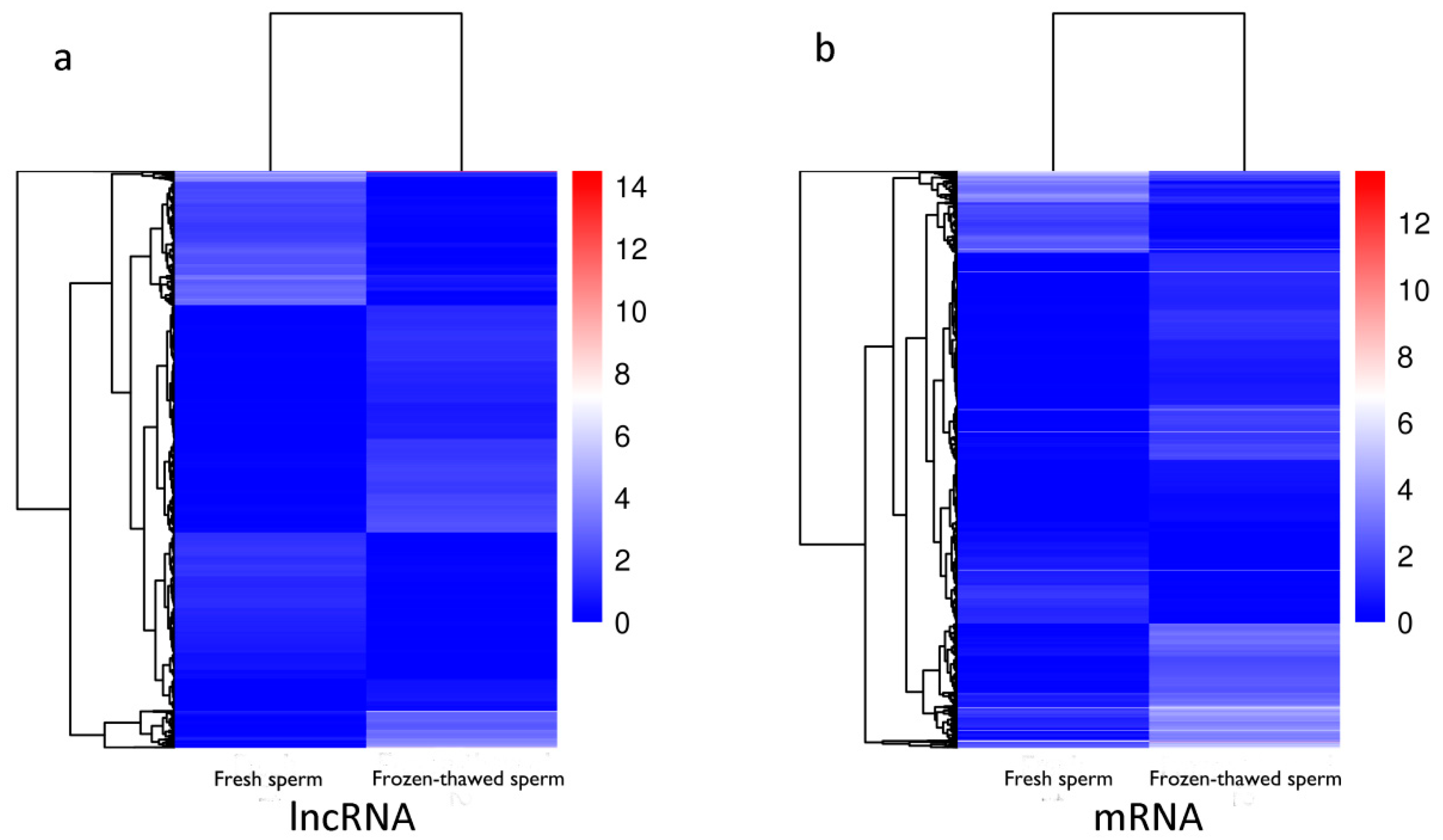
2.5. Characteristic Comparison of lncRNAs and mRNAs
2.6. Differential Expression Analysis
2.7. Target Genes Prediction of cis- and trans-LncRNAs
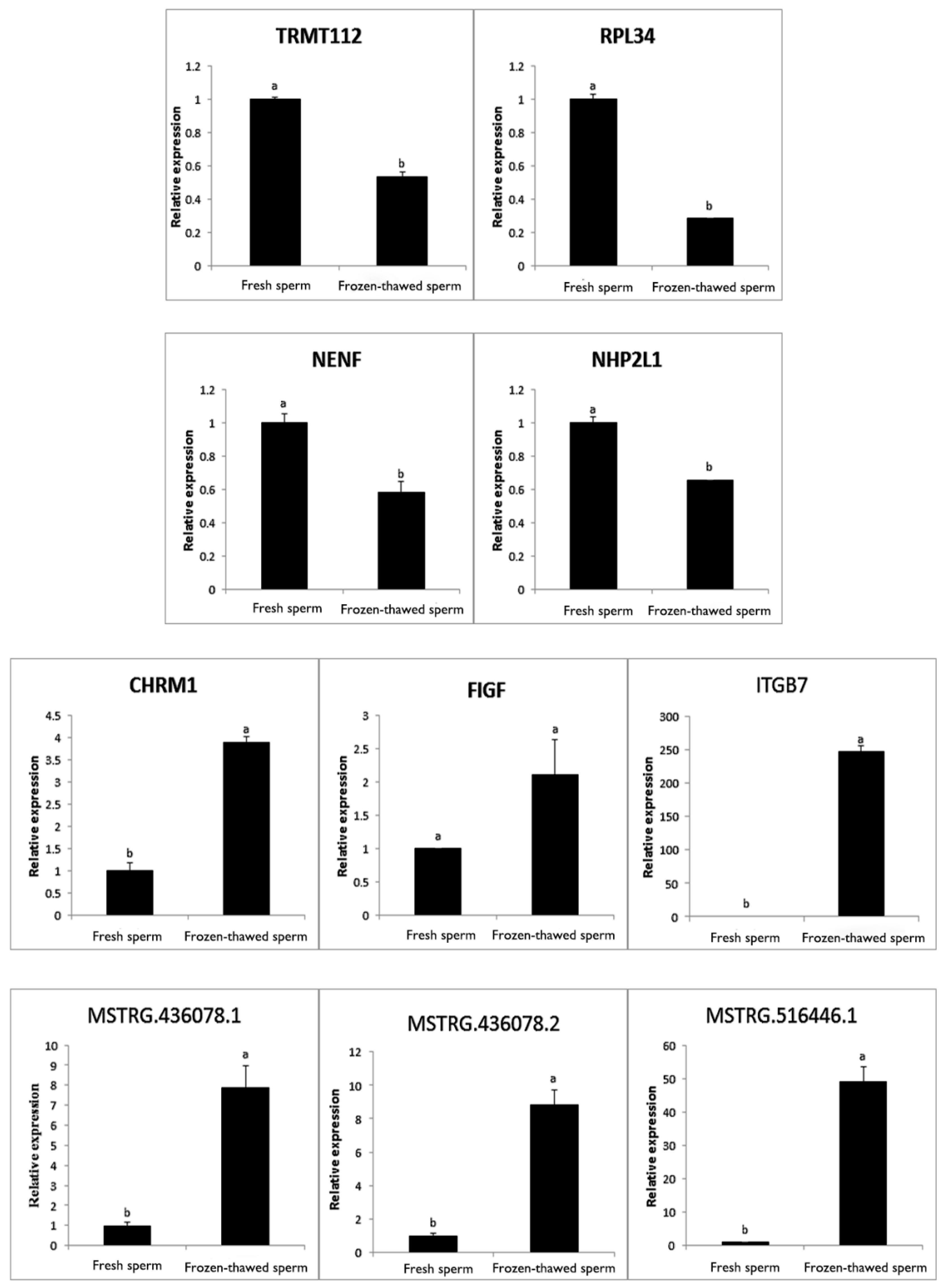
2.8. qRT-PCR Validation
2.9. Functional Enrichment Analysis of GO and KEGG
2.10. Co-Expression of LncRNAs and mRNA
3. Discussion
4. Materials and Methods
4.1. Animal Ethics Statement
4.2. Sperm Collection and Cryopreservation
4.3. Total RNA Extraction, Library Preparation, and Sequencing
4.4. Quality Analysis, Mapping, and Transcriptome Assembly
4.5. Identification and Expression Analysis of lncRNA and mRNA
4.6. Differential Expression Analysis
4.7. Target Gene Prediction
4.8. qRT-PCR Validation
4.9. GO and KEGG Enrichment Analyses
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feng, W.H. The artificial breeding and protecrion of giant panda. Explor. Nat. 1992, 11, 27–32. [Google Scholar]
- Spindler, R.E.; Huang, Y.; Howard, J.G.; Wang, P.; Zhang, H.; Zhang, G.; Wildt, D.E. Acrosomal integrity and capacitation are not influenced by sperm cryopreservation in the giant panda. Reproduction 2004, 127, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, W.H.; Wang, P.Y.; Wang, C.R. Study on semen character of giant panda I. Observation of semen quality. Acta Theriol. Sinica 1991, 11, 1–8. [Google Scholar]
- Huang, Y.; Li, D.; Zhang, H.; Du, J.; Zhang, G.; Wang, P.; Howard, J.; Spindler, R. Studies on spermatozoa in giant pandas. Chin. J. Vet. Med. 2001, 37, 9–11. [Google Scholar]
- Hou, R.; Wang, J.S.; Zhang, H.M.; Lan, J.C.; Wang, C.D.; Yu, J.Q.; Huang, X.M. Development and application of new method for in giant panda semen cryopreservation. Chin. J. Vet. Sci. 2005, 25, 320–322. [Google Scholar]
- Zhou, Y.M.; Wu, D.; Tang, C.X.; Qu, C.M. Effects of adding vitamin b12,BSA and heparin in medium for frozen sperm quality of giant panda. Sichuan J. Zool. 2007, 26, 669–671. [Google Scholar]
- Huang, Y.; Li, D.S.; Zhang, H.M.; Du, J.; Zhang, G.Q.; Wang, P.Y.; Howard, J.; Spindler, R. Study for freezing semen of giant pandas. Chin. J. Zool. 2001, 36, 25–29. [Google Scholar]
- Huang, Y.; Wang, Y.P.; Spindler Rebecca, E.; JoGayle, H.; Zhang, H.; David, E.W. Study on the influence of post-thaw motility on giant panda cryopreserved sperm by thawing rate and pentyoxyfilline in vitro. Acta Theriol. Sinica 2004, 24, 286–292. [Google Scholar]
- Santiago-Moreno, J.; Esteso, M.C.; Pradiee, J.; Castano, C.; Toledano-Diaz, A.; O’Brien, E.; Lopez-Sebastian, A.; Martinez-Nevado, E.; Delclaux, M.; Fernandez-Moran, J.; et al. Giant panda (Ailuropoda melanoleuca) sperm morphometry and function after repeated freezing and thawing. Andrologia 2016, 48, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Pantano, L.; Jodar, M.; Bak, M.; Ballesca, J.L.; Tommerup, N.; Oliva, R.; Vavouri, T. The small RNA content of human sperm reveals pseudogene-derived piRNAs complementary to protein-coding genes. RNA 2015, 21, 1085–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NAZ, R.K. Effect of actinomycin d and cycloheximide on human sperm function. Arch. Androl. 1998, 41, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Sciamanna, I.; Barberi, L.; Martire, A.; Pittoggi, C.; Beraldi, R.; Giordano, R.; Rosa Magnano, A.; Hogdson, C.; Spadafora, C. Sperm endogenous reverse transcriptase as mediator of new genetic information. Biochem. Biophys. Res. Commun. 2003, 312, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Gur, Y.; Breitbart, H. Mammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomes. Genes Dev. 2006, 20, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gur, Y.; Breitbart, H. Protein synthesis in sperm: Dialog between mitochondria and cytoplasm. Mol. Cell. Endocrinol. 2008, 282, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, X.; Li, H.X.; Cui, Q.W.; Yu, J.; Wang, G.L. Delivery of CatSper2 siRNA into rat sperms by electroporation repressed Ca2+ influx during sperm hyperactivation. Agric. Sci. China 2011, 10, 1958–1967. [Google Scholar] [CrossRef]
- Yu, J.; Jiang, X.Q.; Zhou, S.; Wang, G.I. RNA interference-mediated downregulation of sAC gene inhibits sperm hyperactivation in male rats (Rattus norvegicus). J. Integr. Agric. 2014, 13, 394–401. [Google Scholar] [CrossRef]
- Wichman, L.; Somasundaram, S.; Breindel, C.; Valerio, D.M.; McCarrey, J.R.; Hodges, C.A.; Khalil, A.M. Dynamic expression of long noncoding RNAs reveals their potential roles in spermatogenesis and fertility. Biol. Reprod. 2017, 97, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dai, D.; Chang, Y.; Li, Y.; Zhang, M.; Zhou, G.; Peng, Z.; Zeng, C. Cryopreservation of boar sperm induces differential microRNAs expression. Cryobiology 2017, 76, 24–33. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Fan, H.; Liu, Y.F.; Pan, Y.Y.; Liu, Y.K.; Mo, F.F.; Gu, Y.J.; Sun, Y.L.; Zhao, D.D.; Yu, N.; et al. The difference in expression of long noncoding RNAs in rat semen induced by high-fat diet was associated with metabolic pathways. Peer J. 2017, 5, e3518. [Google Scholar] [CrossRef] [PubMed]
- Kornienko, A.E.; Guenzl, P.M.; Barlow, D.P.; Pauler, F.M. Gene regulation by the act of long non-coding RNA transcription. BMC Biol. 2013, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Yang, Y.W.; Liu, B.; Sanyal, A.; Corces-Zimmerman, R.; Chen, Y.; Lajoie, B.R.; Protacio, A.; Flynn, R.A.; Gupta, R.A.; et al. A long noncoding RNA maintains active chromatin to coordinate homeotic gene expression. Nature 2011, 472, 120–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loewer, S.; Cabili, M.N.; Guttman, M.; Loh, Y.H.; Thomas, K.; Park, I.H.; Garber, M.; Curran, M.; Onder, T.; Agarwal, S.; et al. Large intergenic non-coding RNA-RoR modulates reprogramming of human induced pluripotent stem cells. Nat. Genet. 2010, 42, 1113–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Morales, D.R.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, T.; Wang, Y.L.; Lin, M.F.; Koegel, A.K.; Kotake, Y.; Grant, G.D.; Horlings, H.M.; Shah, N.; Umbricht, C.; Wang, P.; et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nat. Genet. 2011, 43, 621–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttman, M.; Donaghey, J.; Carey, B.W.; Garber, M.; Grenier, J.K.; Munson, G.; Young, G.; Lucas, A.B.; Ach, R.; Bruhn, L.; et al. LincRNAs act in the circuitry controlling pluripotency and differentiation. Nature 2011, 477, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, J.; Wu, J.; Schuster, A.S.; Hennig, G.W.; Yan, W. Expression profiling reveals developmentally regulated lncRNA repertoire in the mouse male germline. Biol. Reprod. 2013, 89, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, M.; Wang, M.; Wu, X.; Geng, L.; Xue, Y.; Wei, X.; Jia, Y.; Wu, X. A long non-coding RNA interacts with Gfra1 and maintains survival of mouse spermatogonial stem cells. Cell Death Dis. 2016, 7, e2140. [Google Scholar] [CrossRef] [PubMed]
- Anguera, M.C.; Ma, W.; Clift, D.; Namekawa, S.; Kelleher, R.J., III; Lee, J.T. Tsx produces a long noncoding RNA and has general functions in the germline, stem cells, and brain. PLoS Genet. 2011, 7, e1002248. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Li, X.K.; Zhang, P.; Wang, J.; Zhu, D.D.; Chen, X.P.; Ye, L.H. Low long non-coding RNA hotair expression is associated with down-regulation of nrf2 in the spermatozoa of patients with asthenozoospermia or oligoasthenozoospermia. Int. J. Clin. Exp. Pathol. 2015, 8, 14198–14205. [Google Scholar] [PubMed]
- Ni, M.J.; Hu, Z.H.; Liu, Q.; Liu, M.F.; Lu, M.H.; Zhang, J.S.; Zhang, L.; Zhang, Y.L. Identification and characterization of a novel non-coding RNA involved in sperm maturation. PLoS ONE 2011, 6, e26053. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.J.; Zhang, T.; An, T.; Zhao, D.D.; Yang, X.Y.; Zhang, D.W.; Zhang, Y.; Mu, Q.Q.; Yu, N.; Ma, X.S.; et al. Differential expression of long noncoding RNAs between sperm samples from diabetic and non-diabetic mice. PLoS ONE 2016, 11, e0154028. [Google Scholar] [CrossRef] [PubMed]
- Card, C.J.; Anderson, E.J.; Zamberlan, S.; Krieger, K.E.; Kaproth, M.; Sartini, B.L. Cryopreserved bovine spermatozoal transcript profile as revealed by high-throughput ribonucleic acid sequencing. Biol. Reprod. 2013, 88. [Google Scholar] [CrossRef] [PubMed]
- Said, T.M.; Gaglani, A.; Agarwal, A. Implication of apoptosis in sperm cryoinjury. Reprod. Biomed. Online 2010, 21, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ye, J.; Yang, C.; Zhang, K.; Li, X.; Luo, L.; Ding, J.; Li, Y.; Cao, H.; Ling, Y.; et al. Screening and evaluating of long noncoding RNAs in the puberty of goats. BMC Genom. 2017, 18, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qin, C.; Cao, G.Q.; Xin, W.F.; Feng, C.Q.; Zhang, W.S. Systematic analysis of long noncoding RNAs in the senescence-accelerated mouse prone 8 brain using RNA sequencing. Mol. Ther. Nucleic Acids 2016, 5, e343. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xue, S.; Liu, X.; Liu, H.; Hu, T.; Qiu, X.; Zhang, J.; Lei, M. Analyses of long non-coding RNA and mRNA profiling using RNA sequencing during the pre-implantation phases in pig endometrium. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Cerolini, S.; Maldjian, A.; Pizzi, F.; Gliozzi, T.M. Changes in sperm quality and lipid composition during cryopreservation of boar semen. Reproduction 2001, 121, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, R.A.; Agarwal, A. Oxidative stress and male infertility: From research bench to clinical practice. J. Androl. 2002, 23, 737–752. [Google Scholar] [PubMed]
- Paasch, U.; Sharma, R.K.; Gupta, A.K.; Grunewald, S.; Mascha, E.J.; Thomas, A.J., Jr.; Glander, H.J.; Agarwal, A. Cryopreservation and thawing is associated with varying extent of activation of apoptotic machinery in subsets of ejaculated human spermatozoa. Biol. Reprod. 2004, 71, 1828–1837. [Google Scholar] [CrossRef] [PubMed]
- Eilish, T.; Donnelly, N.M.; Sheena, E.M.; Lewis, S.E. Cryopreservation of human semen and prepared sperm: Effects on motility parameters and DNA integrity. Fertil. Steril. 2001, 76, 892–900. [Google Scholar]
- Kumar, R.; Atreja, S.K. Effect of incorporation of additives in tris-based egg yolk extender on buffalo (Bubalus bubalis) sperm tyrosine phosphorylation during cryopreservation. Reprod. Domest. Anim. 2012, 47, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Pommer, A.C.; Meyers, S.A. Tyrosine phosphorylation is an indicator of capacitation status in fresh and cryopreserved stallion spermatozoa. Theriogenology 2002, 58, 351–354. [Google Scholar]
- Singh, V.K.; Atreja, S.K.; Kumar, R.; Chhillar, S.; Singh, A.K. Assessment of intracellular ca2+, camp and 1,2-diacylglycerol in cryopreserved buffalo (Bubalus bubalis) spermatozoa on supplementation of taurine and trehalose in the extender. Reprod. Domest. Anim. 2012, 47, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Nynca, J.; Arnold, G.J.; Frohlich, T.; Ciereszko, A. Cryopreservation-induced alterations in protein composition of rainbow trout semen. Proteomics 2015, 15, 2643–2654. [Google Scholar] [CrossRef] [PubMed]
- Ortega Ferrusola, C.; Anel-Lopez, L.; Ortiz-Rodriguez, J.M.; Martin Munoz, P.; Alvarez, M.; de Paz, P.; Masot, J.; Redondo, E.; da Silva, C.B.; Morrell, J.M.; et al. Stallion spermatozoa surviving freezing and thawing experience membrane depolarization and increased intracellular na. Andrology 2017, 5, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Capra, E.; Turri, F.; Lazzari, B.; Cremonesi, P.; Gliozzi, T.M.; Fojadelli, I.; Stella, A.; Pizzi, F. Small RNA sequencing of cryopreserved semen from single bull revealed altered miRNAs and piRNAs expression between high- and low-motile sperm populations. BMC Genom. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Naresh, S.; Atreja, S.K. The protein tyrosine phosphorylation during in vitro capacitation and cryopreservation of mammalian spermatozoa. Cryobiology 2015, 70, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Maquat, L.E. LncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 3′ UTRs via Alu elements. Nature 2011, 470, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Nagano, T.; Mitchell, J.A.; Sanz, L.A.; Pauler, F.M.; Ferguson-Smith, A.C.; Feil, R.; Fraser, P. The air noncoding RNA epigenetically silences transcription by targeting G9a to chromatin. Science 2008, 322, 1717–1720. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.H.; Liu, X.N.; Wang, T.T.; Pan, W.; Tao, Q.F.; Zhou, W.P.; Wang, F.; Sun, S.H. The MBNL3 splicing factor promotes hepatocellular carcinoma by increasing pxn expression through the alternative splicing of lncRNA-PXN-AS1. Nat. Cell Biol. 2017, 19, 820–832. [Google Scholar] [CrossRef] [PubMed]
- Rosales, J.L.; Lee, B.C.; Modarressi, M.; Sarker, K.P.; Lee, K.Y.; Jeong, Y.G.; Oko, R.; Lee, K.Y. Outer dense fibers serve as a functional target for cdk5.P35 in the developing sperm tail. J. Biol. Chem. 2004, 279, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, J.; Liu, X.; Xu, R.; Zhang, Y. Correlation of appearance of metastasis-associated protein1 (Mta1) with spermatogenesis in developing mouse testis. Cell Tissue Res. 2007, 329, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, S.E.; Bajpai, M.; Carr, D.W. Identification and characterization of RHOA-interacting proteins in bovine spermatozoa. Biol. Reprod. 2008, 78, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Zhang, Y.; Li, S.; Sun, W.; Soong, T.W. Tyrosine kinase-independent inhibition by genistein on spermatogenic T-type calcium channels attenuates mouse sperm motility and acrosome reaction. Cell Calcium 2009, 45, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, Y.; Miao, S.; Zhang, C.; Zong, S.; Koide, S.S.; Wang, L. Sperm associated antigen 8 (SPAG8), a novel regulator of activator of crem in testis during spermatogenesis. FEBS Lett. 2010, 584, 2807–2815. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Zhang, J.; Zheng, H.; Xu, C.; Yan, W. Ubqln1 interacts with spem1 and participates in spermiogenesis. Mol. Cell. Endocrinol. 2010, 327, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Marcello, M.R.; Jia, W.; Leary, J.A.; Moore, K.L.; Evans, J.P. Lack of tyrosylprotein sulfotransferase-2 activity results in altered sperm-egg interactions and loss of adam3 and adam6 in epididymal sperm. J. Biol. Chem. 2011, 286, 13060–13070. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shen, C.; Fan, W.; Chen, Y.; Zhang, A.; Feng, Y.; Li, Z.; Kuang, Y.; Wang, Z. Low levels of PRSS37 protein in sperm are associated with many cases of unexplained male infertility. Acta Biochim. Biophys. Sin. 2016, 48, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Kuang, Y.; Liu, J.; Feng, J.; Chen, X.; Wu, W.; Chi, J.; Tang, L.; Wang, Y.; Fei, J.; et al. PRSS37 is required for male fertility in the mouse. Biol. Reprod. 2013, 88. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Guillemain, C.; Victorero, G.; Lepoivre, C.; Bergon, A.; Yammine, M.; Perrin, J.; Sari-Minodier, I.; Boulanger, N.; Rihet, P.; Nguyen, C. Sperm mRNAs and microRNAs as candidate markers for the impact of toxicants on human spermatogenesis: An application to tobacco smoking. Syst. Biol. Reprod. Med. 2015, 61, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Ernesto, J.I.; Weigel Munoz, M.; Battistone, M.A.; Vasen, G.; Martinez-Lopez, P.; Orta, G.; Figueiras-Fierro, D.; de la Vega-Beltran, J.L.; Moreno, I.A.; Guidobaldi, H.A.; et al. CRISP1 as a novel CatSper regulator that modulates sperm motility and orientation during fertilization. J. Cell Biol. 2015, 210, 1213–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, H.L.; Mansell, S.; Alasmari, W.; Brown, S.G.; Wilson, S.M.; Sutton, K.A.; Miller, M.R.; Lishko, P.V.; Barratt, C.L.; Publicover, S.J.; et al. Specific loss of CatSper function is sufficient to compromise fertilizing capacity of human spermatozoa. Hum. Reprod. 2015, 30, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Kumaresan, A.; Kumar, M.; Chhillar, S.; Malik, H.; Kumar, S.; Kaushik, J.K.; Datta, T.K.; Mohanty, A.K. Effect of recombinant and native buffalo OVGP1 on sperm functions and in vitro embryo development: A. comparative study. J. Anim. Sci. Biotechnol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Panigrahi, S.K.; Manterola, M.; Wolgemuth, D.J. Meiotic failure in cyclin A1-deficient mouse spermatocytes triggers apoptosis through intrinsic and extrinsic signaling pathways and 14-3-3 proteins. PLoS ONE 2017, 12, e0173926. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.N.; Ma, Y.; Cress, W.D. RhoBTB2 (DBC2) is a mitotic E2F1 target gene with a novel role in apoptosis. J. Biol. Chem. 2008, 283, 2353–2362. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.K.; Pok, S.; Streit, S.; Ruhe, J.E.; Hart, S.; Lim, K.S.; Loo, H.L.; Aung, M.O.; Lim, S.G.; Ullrich, A. Fibroblast growth factor receptor 4 regulates proliferation, anti-apoptosis and alpha-fetoprotein secretion during hepatocellular carcinoma progression and represents a potential target for therapeutic intervention. J. Hepatol. 2009, 50, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Kai, L.; Samuel, S.K.; Levenson, A.S. Resveratrol enhances p53 acetylation and apoptosis in prostate cancer by inhibiting MTA1/NuRD complex. Int. J. Cancer 2010, 126, 1538–1548. [Google Scholar] [CrossRef] [PubMed]
- Baron, B.W.; Hyjek, E.; Gladstone, B.; Thirman, M.J.; Baron, J.M. PDCD2, a protein whose expression is repressed by BCL6, induces apoptosis in human cells by activation of the caspase cascade. Blood Cell. Mol. Dis. 2010, 45, 169–175. [Google Scholar] [CrossRef] [PubMed]
- N’Guessan, P.; Pouyet, L.; Gosset, G.; Hamlaoui, S.; Seillier, M.; Cano, C.E.; Seux, M.; Stocker, P.; Culcasi, M.; Iovanna, J.L.; et al. Absence of tumor suppressor tumor protein 53-induced nuclear protein 1 (TP53INP1) sensitizes mouse thymocytes and embryonic fibroblasts to redox-driven apoptosis. Antioxid. Redox Signal. 2011, 15, 1639–1653. [Google Scholar] [CrossRef] [PubMed]
- Lo, P.H.; Ko, J.M.; Yu, Z.Y.; Law, S.; Wang, L.D.; Li, J.L.; Srivastava, G.; Tsao, S.W.; Stanbridge, E.J.; Lung, M.L. The lim domain protein, CRIP2, promotes apoptosis in esophageal squamous cell carcinoma. Cancer Lett. 2012, 316, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, T. T-type voltage-activated calcium channel Cav3.1, but not Cav3.2, is involved in the inhibition of proliferation and apoptosis in MCF-7 human breast cancer cells. Int. J. Oncol. 2012, 41, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, Y.H.; Li, M.Q.; Meng, Y.H.; Chen, X.; Shao, J.; Tang, C.L.; Du, M.R.; Jin, L.P.; Li, D.J.; et al. Trophoblasts-derived chemokine CCL24 promotes the roliferation, growth and apoptosis of decidual stromal cells in human early pregnancy. Int. J. Clin. Exp. Pathol. 2013, 6, 1028–1037. [Google Scholar] [PubMed]
- Bruchmann, A.; Roller, C.; Walther, T.V.; Schäfer, G.; Lehmusvaara, S.; Visakorpi, T.; Klocker, H.; Cato, A.C.; Maddalo, D. Bcl-2 associated athanogene 5 (Bag5) is overexpressed in prostate cancer and inhibits ER-stress induced apoptosis. BMC Cancer 2013, 13. [Google Scholar] [CrossRef]
- Maskell, L.J.; Qamar, K.; Babakr, A.A.; Hawkins, T.A.; Heads, R.J.; Budhram-Mahadeo, V.S. Essential but partially redundant roles for POU4F1/Brn-3a and POU4F2/Brn-3b transcription factors in the developing heart. Cell Death. Dis. 2017, 8, e2861. [Google Scholar] [CrossRef] [PubMed]
- Budhram-Mahadeo, V.; Fujita, R.; Bitsi, S.; Sicard, P.; Heads, R. Co-expression of POU4F2/Brn-3b with p53 may be important for controlling expression of pro-apoptotic genes in cardiomyocytes following ischaemic/hypoxic insults. Cell Death. Dis. 2014, 5, e1503. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hu, H.; Shi, X.; Tang, W. Knockdown of S100P by lentiviral-mediated rnai promotes apoptosis and suppresses the colony-formation ability of gastric cancer cells. Oncol. Rep. 2014, 31, 2344–2350. [Google Scholar] [CrossRef] [PubMed]
- Ke, K.; Shen, J.; Song, Y.; Cao, M.; Lu, H.; Liu, C.; Shen, J.; Li, A.; Huang, J.; Ni, H.; et al. CDK5 contributes to neuronal apoptosis via promoting MEF2D phosphorylation in rat model of intracerebral hemorrhage. J. Mol. Neurosci. 2015, 56, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Keenan, M.M.; Liu, B.; Tang, X.; Wu, J.; Cyr, D.; Stevens, R.D.; Ilkayeva, O.; Huang, Z.; Tollini, L.A.; Murphy, S.K.; et al. ACLY and ACC1 regulate hypoxia-induced apoptosis by modulating etv4 via α-ketoglutarate. PLoS Genet. 2015, 11, e1005599. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.; Wan, B.; Peng, J.; Yang, Y.; Lv, D.; Zhou, X.; Shu, F.; Li, F.; Zhong, L.; Wu, H.; et al. PRPS2 expression correlates with sertoli-cell only syndrome and inhibits the apoptosis of tm4 sertoli cells. J. Urol. 2015, 194, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Parmigiani, A.; Yang, C.; Budanov, A.V. Sestrin2 facilitates death receptor-induced apoptosis in lung adenocarcinoma cells through regulation of XIAP degradation. Cell Cycle 2015, 14, 3231–3241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.Y.; Cao, Z.H.; Hu, X.B.; Li, G.Q.; Dong, S.F.; Xu, G.L.; Zhang, K.Q. Light/IFN-γ triggers β cells apoptosis via NF-κB/Bcl2-dependent mitochondrial pathway. J. Cell. Mol. Med. 2016, 20, 1861–1871. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jia, G.; Shi, B.; Ge, G.; Duan, H.; Yang, Y. PRSS8 is downregulated and suppresses tumour growth and metastases in hepatocellular carcinoma. Cell. Physiol. Biochem. 2016, 40, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Wang, L.; Zhong, C.; Qi, R.; Li, Y. AHSA1 regulates proliferation, apoptosis, migration, and invasion of osteosarcoma. Biomed. Pharmacother. 2016, 77, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Jourd’heuil, F.L.; Xu, H.; Reilly, T.; McKellar, K.; Alaoui, C.E.; Steppich, J.; Liu, Y.F.; Zhao, W.; Ginnan, R.; David Conti, R.L.-S.; et al. The hemoglobin homolog cytoglobin in smooth muscle inhibits apoptosis and regulates vascular remodeling. Arterioscle. Thromb. Vasc. Biol. 2017, 37, 1944–1955. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Wang, S.; Ren, B.; Wang, J.; Chen, J.; Lu, J.; Zhan, S.; Fu, Y.; Huang, L.; Tan, J. CHOP favors endoplasmic reticulum stress-induced apoptosis in hepatocellular carcinoma cells via inhibition of autophagy. PLoS ONE 2017, 12, e0183680. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Mei, X.; Wang, L.; Yang, X. RhoA inhibits apoptosis and increases proliferation of cultured SPCA1 lung cancer cells. Mol. Med. Rep. 2017, 15, 3963–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savino, A.M.; Sarno, J.; Trentin, L.; Vieri, M.; Fazio, G.; Bardini, M.; Bugarin, C.; Fossati, G.; Davis, K.L.; Gaipa, G.; et al. The histone deacetylase inhibitor givinostat (ITF2357) exhibits potent anti-tumor activity against CRLF2-rearranged BCP-ALL. Leukemia 2017, 31, 2365–2375. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, D.S.; Zhang, H.M.; Du, J.; Zhang, G.Q.; Wang, P.Y.; Howard, J.G.; Spindler, R.; Durrant, B.; Olson, M.A. Electroeljaculation and semen cryopreservation in giant pandas. J. Sichuan Teach. Coll. 2000, 21, 238–243. [Google Scholar]
- Agarwal, A.; Sharma, R.K. Automation is the key to standardized semen analysis using the automated SQA-V sperm quality analyzer. Fertil. Steril. 2007, 87, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; He, L.; Peng, W.; Ding, L.; Tang, K.; Fang, D.; Zhang, Y. Selection of optimal reference genes for quantitative RT-PCR studies of boar spermatozoa cryopreservation. Cryobiology 2014, 68, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, W.; Zeng, P.; Wang, J.; Geng, B.; Yang, J.; Cui, Q. LncTar: A tool for predicting the RNA targets of long noncoding RNAs. Brief. Bioinform. 2015, 16, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| lncRNA | log2FC | Target mRNA | log2FC | Description |
|---|---|---|---|---|
| MSTRG.122368.1 | 3.14 | CDK5 | 2.76 | Regulating of sperm tail development [51] |
| MSTRG.655028.1 | −2.86 | MTA1 | −3.33 | Crucial for spermatogenesis [52] |
| MSTRG.332212.2 | 3.86 | RHOA | −3.02 | Involved in capacitation and the acrosome reaction [53] |
| MSTRG.531884.1 | 3.93 | CACNA1G(Cav3.1) | 2.74 | Regulate male fertility in mice [54] |
| MSTRG.264076.1 | 3.81 | SPAG8 | 3.18 | Cell division during spermatogenesis [55] |
| MSTRG.480606.2 | 4.12 | SPEM1 | -2.53 | Spermatogenesis-essential proteins [56] |
| MSTRG.408558.1 | 3.43 | TPST2 | 3.01 | Male infertility, sperm motility defects [57] |
| MSTRG.369918.1 | 5.54 | PRSS37 | −3.28 | Prss37 deletion markedly decreased fertilization rate [58,59] |
| MSTRG.1467.1 | 2.81 | CNGB1 | 4.88 | Differentially represented between smokers and non-smokers’ sperm [60] |
| MSTRG.169617.1 | 4.46 | CATSPER1 | −2.91 | Involved in hyperactivation and essential for fertility [61], functional failure of CatSper is sufficient to compromise fertility of human sperm [62] |
| MSTRG.597193.1 | 3.97 | OVGP1 | 2.66 | Sustaining the sperm functions, include motile, membrane intact, proportion of capacitated and acrosome- reacted [63] |
| MSTRG.651082.1 | 3.38 | CCNA1 | −3.08 | Essential for spermatogenesis in the mouse. Ccna1- deficient spermatocytes arrest at late meiotic prophase and undergo apoptosis [64] |
| MSTRG.651082.3 | −3.30 |
| Gene Name | Log2FC | Function | Ref. | lncRNA | Log2FC |
|---|---|---|---|---|---|
| RHOBTB2 | 2.87 | + | [65] | MSTRG.531884.1 | 3.93 |
| FGFR4 | 3.37 | + | [66] | MSTRG.628795.1 | 4.88 |
| MTA1 | −3.33 | − | [67] | MSTRG.655028.1 | −2.86 |
| PDCD2 | −2.72 | + | [68] | MSTRG.509816.1 | 4.02 |
| TP53INP1 | −2.91 | + | [69] | MSTRG.561458.1 | −3.23 |
| CRIP2 | −3.74 | + | [70] | MSTRG.655028.1 | −2.86 |
| CACNA1G | 2.74 | + | [71] | MSTRG.531884.1 | 3.93 |
| CCL24 | 5.23 | + | [72] | MSTRG.444753.1 | 4.88 |
| BAG5 | −2.70 | − | [73] | MSTRG.263206.1 | 4.14 |
| POU4F1 | 4.17 | + | [74,75] | MSTRG.306867.1 | −3.32 |
| S100P | −2.52 | − | [76] | MSTRG.552283.1 | 3.52 |
| CDK5 | 2.76 | + | [77] | MSTRG.122368.1 | 3.14 |
| ETV4 | 2.60 | + | [78] | MSTRG.298973.1 | 4.73 |
| PRPS2 | −2.66 | − | [79] | MSTRG.101520.1 | −5.39 |
| MSTRG.101521.1 | −4.27 | ||||
| SESN2 | 5.16 | + | [80] | MSTRG.33577.3 | 3.73 |
| TNFSF14 (LIGHT) | 5.66 | + | [81] | MSTRG.609960.1 | −3.08 |
| PRSS8 | 4.58 | + | [82] | MSTRG.493468.1 | 4.16 |
| AHSA1 | −2.66 | − | [83] | MSTRG.555075.1 | 5.64 |
| CYGB | 5.05 | − | [84] | MSTRG.301513.1 | −5.91 |
| DDIT3 (CHOP) | −2.70 | + | [85] | MSTRG.141461.2 | 5.18 |
| RHOA | −3.02 | − | [86] | MSTRG.332212.2 | 3.86 |
| CRLF2 | 3.02 | − | [87] | MSTRG.343934.1 | −4.49 |
| Gene Name | Primer Sequence (5′–3′) | Amplicon (bp) |
|---|---|---|
| ITGB7 | F: AGGTCTCATCCCCCGAGAAG | 168 |
| R: CGTACACAGGGTTCAAAGGC | ||
| MSTRG.436078.1 | F: CAGGCTTCCTCCTCTCTCCA | 143 |
| R: CCACCAGATCTCAAGGACAGC | ||
| MSTRG.436078.2 | F: GCCTGTCTCATTGCTCAAGGT | 114 |
| R: GGACTATTCTGGTAGCTGTGTCCA | ||
| CHRM1 | F: GCAAGTGGCTTTCATTGGGA | 133 |
| R: CAGGCTGAGCAGGAAGTAGT | ||
| MSTRG.516446.1 | F: GGAGAATTACGGTGGGATGAC | 149 |
| R: AAGAAAACACTAACGCAGAAAGG | ||
| FIGF(VEGFD) | F: AAGGAGAAGAGGGCTGCCTA | 117 |
| R: GACAGCAACTTGGCAAAGCA | ||
| NENF | F: TGGCAGTGAAAGGAGTGGTGTT | 151 |
| R: CCCGTAGTGTCATGGGTGAGGT | ||
| RPL34 | F: AAACTAGGCTGTCCCGAACC | 119 |
| R: AGCACGAACTCCTCGAAGTC | ||
| NHP2L1 | F: CGGAAAGGAGCCAATGAAG | 205 |
| R: CAGAACAGGCGATGACAGG | ||
| TRMT112 | F: TGGCGCGTATGATACCCAAG | 209 |
| R: GCGACTGATGGGGAACAGAT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, M.-X.; Li, Y.; Zhang, Y.; Liang, K.; Ren, Y.-N.; Zhang, M.; Zhou, G.-B.; Zhou, Y.-M.; Wu, K.; Wang, C.-D.; et al. Transcriptome Sequencing Reveals the Differentially Expressed lncRNAs and mRNAs Involved in Cryoinjuries in Frozen-Thawed Giant Panda (Ailuropoda melanoleuca) Sperm. Int. J. Mol. Sci. 2018, 19, 3066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103066
Ran M-X, Li Y, Zhang Y, Liang K, Ren Y-N, Zhang M, Zhou G-B, Zhou Y-M, Wu K, Wang C-D, et al. Transcriptome Sequencing Reveals the Differentially Expressed lncRNAs and mRNAs Involved in Cryoinjuries in Frozen-Thawed Giant Panda (Ailuropoda melanoleuca) Sperm. International Journal of Molecular Sciences. 2018; 19(10):3066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103066
Chicago/Turabian StyleRan, Ming-Xia, Yuan Li, Yan Zhang, Kai Liang, Ying-Nan Ren, Ming Zhang, Guang-Bin Zhou, Ying-Min Zhou, Kai Wu, Cheng-Dong Wang, and et al. 2018. "Transcriptome Sequencing Reveals the Differentially Expressed lncRNAs and mRNAs Involved in Cryoinjuries in Frozen-Thawed Giant Panda (Ailuropoda melanoleuca) Sperm" International Journal of Molecular Sciences 19, no. 10: 3066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103066