Ciprofloxacin Enhances TRAIL-Induced Apoptosis in Lung Cancer Cells by Upregulating the Expression and Protein Stability of Death Receptors through CHOP Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Ciprofloxacin Potentiated TRAIL-Induced Apoptosis in Human Lung Cancer Cells
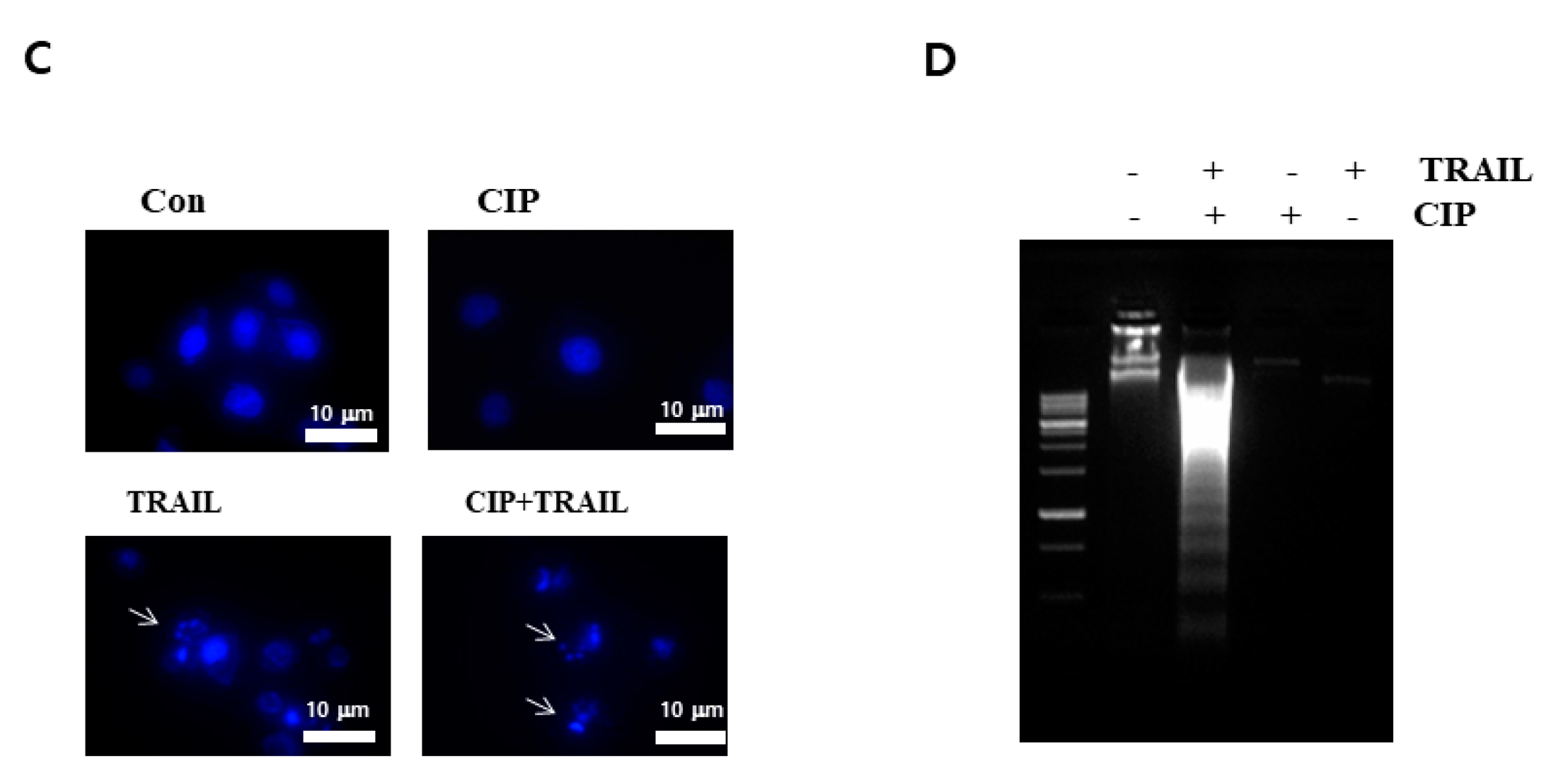
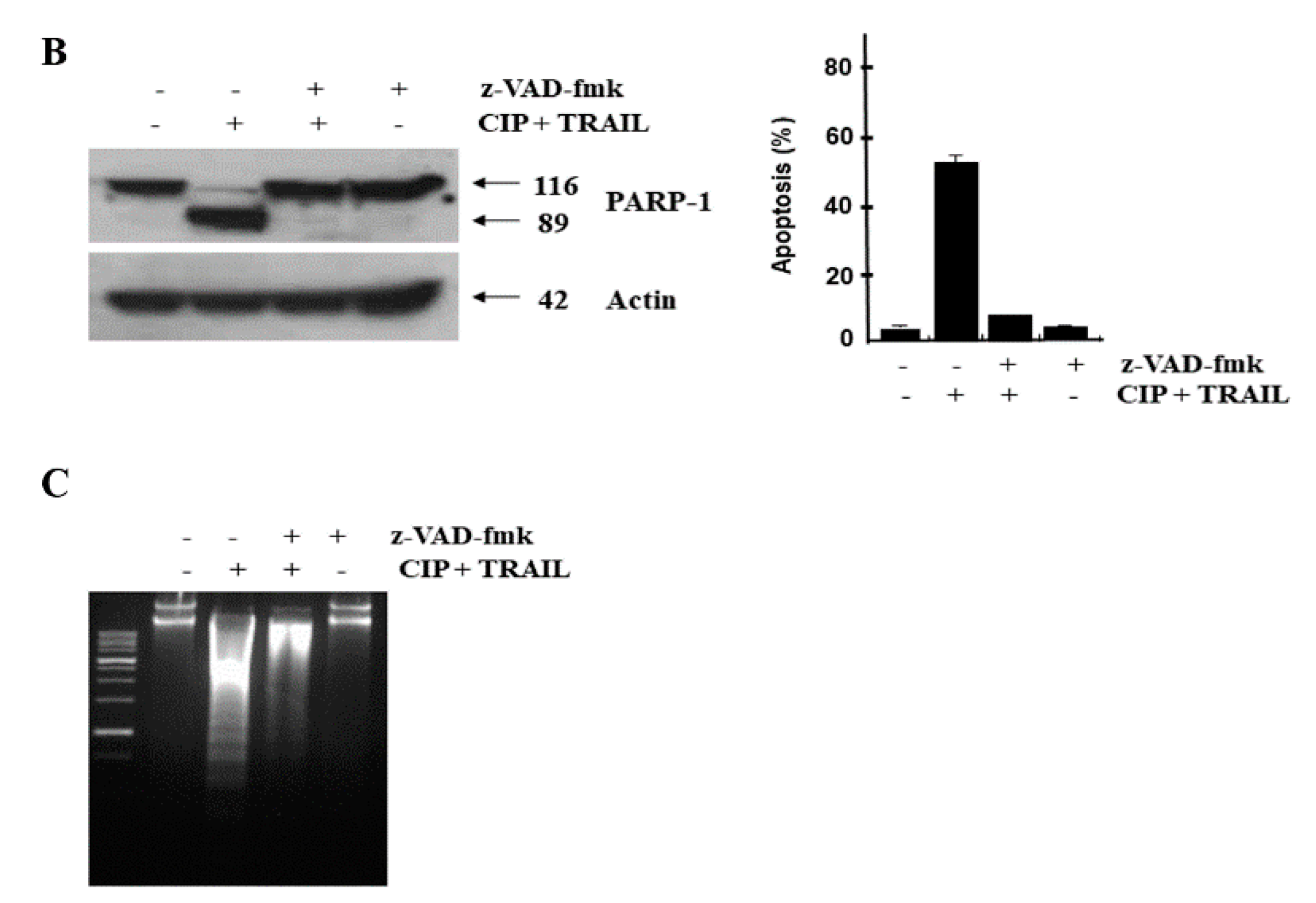
2.2. CIP Sensitized TRAIL-Induced Apoptosis through Caspase Pathway
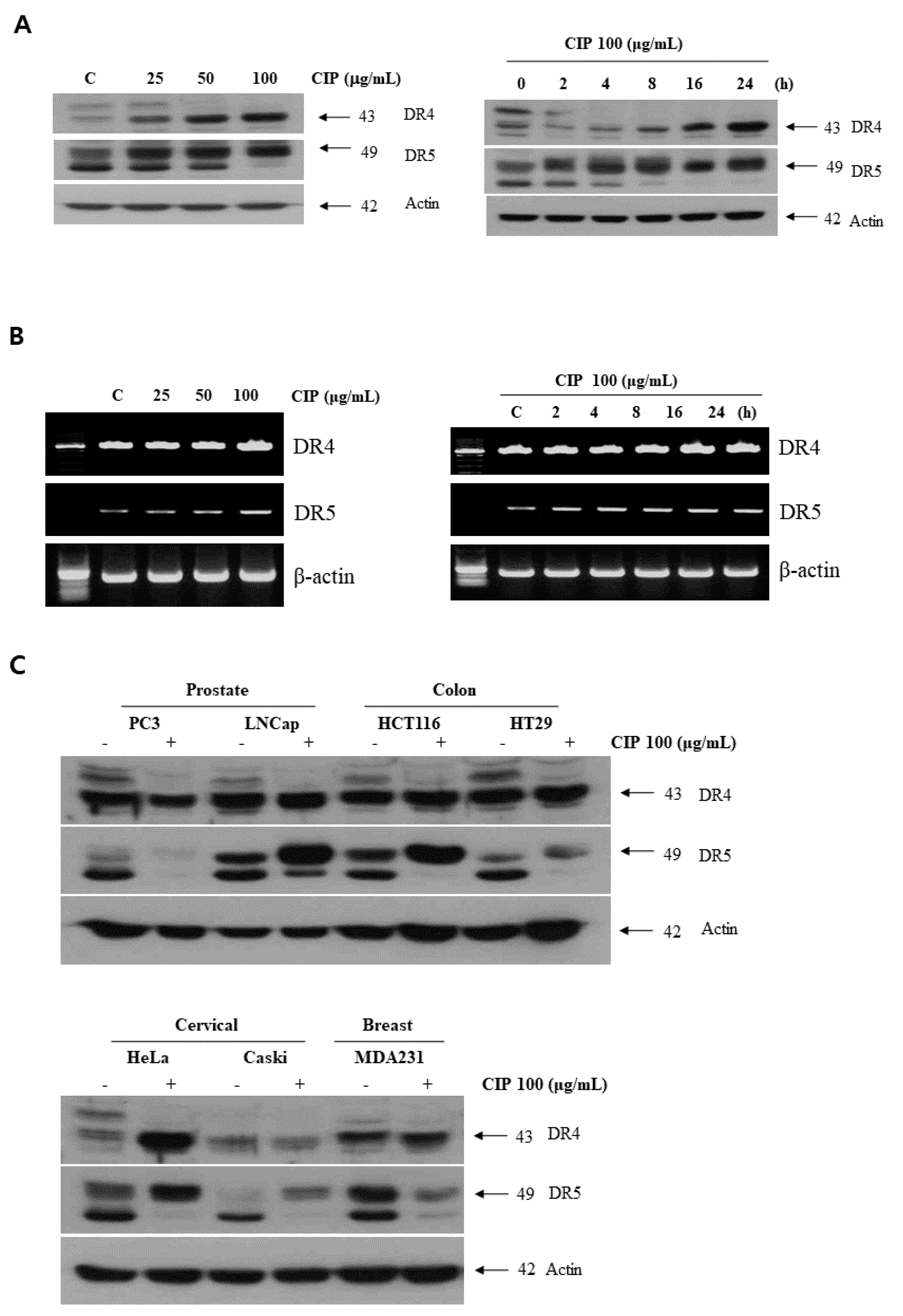
2.3. CIP Upregulated Death Receptors Expression in Various Cancer Cells
2.4. Death Receptor Was Required by CIP to Enhance TRAIL-Induced Apoptosis
2.5. CHOP Mediated CIP-Induced Death Receptors Upregulation and Sensitization to TRAIL
2.6. CIP Sustained Death Receptor Protein Stability
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Chemical Treatments
4.3. Cellular Viability Assay
4.4. Flow Cytometric Assay of DNA Content
4.5. Analysis of Cytochrome C Release
4.6. Western Blot Analysis
4.7. DNA Fragmentation and 4’,6’-Diamidino-2-Phenylindole (DAPI) Staining Assay
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TRAIL | Tumor necrosis factor (TNF)-related apoptosis-inducing ligand |
| FADD | Fas-associated protein with death domain |
| DISC | Death-inducing signaling complex |
| FQ | Fluoroquinolone |
| CHOP | CCAAT/enhancer-binding protein homologous protein |
| CHX | Cycloheximide |
| PARP | Poly (ADP-ribose) polymerase |
| PI | Propidium iodide |
| NAC | N-acetyl-l-cyteine |
| ROS | Reactive oxygen species |
References
- Sheridan, J.P.; Marsters, S.A.; Pitti, R.M.; Gurney, A.; Skubatch, M.; Baldwin, D.; Ramakrishnan, L.; Gray, C.L.; Baker, K.; Wood, W.I.; et al. Control of TRAIL-induced apoptosis by a family of signaling and decoy receptors. Science 1997, 277, 818–821. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Pai, R.C.; Fong, S.; Leung, S.; Lawrence, D.A.; Marsters, S.A.; Blackie, C.; Chang, L.; McMurtrey, A.E.; Hebert, A.; et al. Safety and antitumor activity of recombinant soluble Apo2 ligand. J. Clin. Investig. 1999, 104, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walczak, H.; Miller, R.E.; Ariail, K.; Gliniak, B.; Griffith, T.S.; Kubin, M.; Chin, W.; Jones, J.; Woodward, A.; Le, T.; et al. Tumoricidal activity of tumor necrosis factor-related apoptosis-inducing ligand in vivo. Nat. Med. 1999, 5, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S. Apoptosis by death factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; An, J.Y.; Kwon, Y.T.; Lee, Y.J. Evidence for two modes of development of acquired tumor necrosis factor-related apoptosis-inducing ligand resistance. Involvement of Bcl-xL. J. Biol. Chem. 2007, 282, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fang, B. Mechanisms of resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther. 2005, 12, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Maksimovic-Ivanic, D.; Stosic-Grujicic, S.; Nicoletti, F.; Mijatovic, S. Resistance to TRAIL and how to surmount it. Immunol. Res. 2012, 52, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Drlica, K. Mechanism of fluoroquinolone action. Curr. Opin. Microbiol. 1999, 2, 504–508. [Google Scholar] [CrossRef]
- Shen, L.L.; Baranowski, J.; Pernet, A.G. Mechanism of inhibition of DNA gyrase by quinolone antibacterials: Specificity and cooperativity of drug binding to DNA. Biochemistry 1989, 28, 3879–3885. [Google Scholar] [CrossRef] [PubMed]
- Dalhoff, A.; Shalit, I. Immunomodulatory effects of quinolones. Lancet Infect. Dis. 2003, 3, 359–371. [Google Scholar] [CrossRef]
- Dalhoff, A. Immunomodulatory activities of fluoroquinolones. Infection 2005, 33, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Shoenfeld, Y.; Sherer, Y.; Fishman, P. Interleukin-3 and pregnancy loss in antiphospholipid syndrome. Scand. J. Rheumatol. Suppl. 1998, 107, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Savion, S.; Blank, M.; Shepshelovich, J.; Fishman, P.; Shoenfeld, Y.; Toder, V. Ciprofloxacin affects pregnancy loss in CBA/JxDBA/2J mice possibly via elevation of interleukin-3 and granulocyte macrophage-colony stimulating factor production. Am. J. Reprod. Immunol. 2000, 44, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Blank, M.; George, J.; Fishman, P.; Levy, Y.; Toder, V.; Savion, S.; Barak, V.; Koike, T.; Shoenfeld, Y. Ciprofloxacin immunomodulation of experimental antiphospholipid syndrome associated with elevation of interleukin-3 and granulocyte-macrophage colony-stimulating factor expression. Arthritis Rheum. 1998, 41, 224–232. [Google Scholar] [CrossRef]
- Stein, R.B.; Hanauer, S.B. Medical therapy for inflammatory bowel disease. Gastroenterol. Clin. N. Am. 1999, 28, 297–321. [Google Scholar] [CrossRef]
- Rath, H.C.; Schultz, M.; Freitag, R.; Dieleman, L.A.; Li, F.; Linde, H.J.; Scholmerich, J.; Sartor, R.B. Different subsets of enteric bacteria induce and perpetuate experimental colitis in rats and mice. Infect. Immun. 2001, 69, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.J.; Keft, A.F. A review on the strategies for the development and application of new anti-arthritic agents. Immunopharmacol. Immunotoxicol. 1995, 17, 607–663. [Google Scholar] [CrossRef] [PubMed]
- Breban, M.; Fournier, C.; Gougerot-Pocidalo, M.A.; Muffat-Joly, M.; Pocidalo, J.J. Protective effects of ciprofloxacin against type II collagen induced arthritis in rats. J. Rheumatol. 1992, 19, 216–222. [Google Scholar] [PubMed]
- Gurbay, A.; Osman, M.; Favier, A.; Hincal, F. Ciprofloxacin-Induced Cytotoxicity and Apoptosis in HeLa Cells. Toxicol. Mech. Methods 2005, 15, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Herold, C.; Ocker, M.; Ganslmayer, M.; Gerauer, H.; Hahn, E.G.; Schuppan, D. Ciprofloxacin induces apoptosis and inhibits proliferation of human colorectal carcinoma cells. Br. J. Cancer 2002, 86, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Mondal, E.R.; Das, S.K.; Mukherjee, P. Comparative evaluation of antiproliferative activity and induction of apoptosis by some fluoroquinolones with a human non-small cell lung cancer cell line in culture. Asian Pac. J. Cancer Prev. 2004, 5, 196–204. [Google Scholar] [PubMed]
- Reuveni, D.; Halperin, D.; Shalit, I.; Priel, E.; Fabian, I. Quinolones as enhancers of camptothecin-induced cytotoxic and anti-topoisomerase I. effects. Biochem. Pharmacol. 2008, 75, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Reuveni, D.; Halperin, D.; Fabian, I.; Tsarfaty, G.; Askenasy, N.; Shalit, I. Moxifloxacin increases anti-tumor and anti-angiogenic activity of irinotecan in human xenograft tumors. Biochem. Pharmacol. 2010, 79, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; Szczepanski, M.J.; Kim, S.Y.; Kim, J.H.; An, J.Y.; Kwon, Y.T.; Alcala, M.A., Jr.; Bartlett, D.L.; Lee, Y.J. c-Cbl-mediated degradation of TRAIL receptors is responsible for the development of the early phase of TRAIL resistance. Cell Signal. 2010, 22, 553–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranha, O.; Grignon, R.; Fernandes, N.; McDonnell, T.J.; Wood, D.P.J.; Sarkar, F.H. Suppression of human prostate cancer cell growth by ciprofloxacin is associated with cell cycle arrest and apoptosis. Int. J. Oncol. 2003, 22, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Jun, Y.T.; Kim, H.J.; Song, M.J.; Lim, J.H.; Lee, D.G.; Han, K.J.; Choi, S.M.; Yoo, J.H.; Shin, W.S.; Choi, J.H. In vitro effects of ciprofloxacin and roxithromycin on apoptosis of jurkat T lymphocytes. Antimicrob. Agents Chemother. 2003, 47, 1161–1164. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.W.; Claire, D.C.; Weissig, V.; Rowe, T.C. Delayed cytotoxicity and cleavage of mitochondrial DNA in ciprofloxacin-treated mammalian cells. Mol. Pharmacol. 1996, 50, 1178–1188. [Google Scholar] [PubMed]
- Aranha, O.; Wood, D.P., Jr.; Sarkar, F.H. Ciprofloxacin mediated cell growth inhibition, S/G2-M cell cycle arrest, and apoptosis in a human transitional cell carcinoma of the bladder cell line. Clin. Cancer Res. 2000, 6, 891–900. [Google Scholar] [PubMed]
- Dida, F.; Li, Y.; Iwao, A.; Deguchi, T.; Azuma, E.; Komada, Y. Resistance to TRAIL induced apoptosis caused by constitutional phosphorylation of Akt and PTEN in acute lymphoblastic leukemia cells. Exp. Hematol. 2008, 36, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- Reuss, D.E.; Mucha, J.; Hagenlocher, C.; Ehemann, V.; Kluwe, L.; Manutner, V.; von Deimling, A. Sensitivity of malignant peripheral nerve sheath tumor cells to TRAIL is augmented by loss of NF1 through modulation of MYC/MAD and is potentiated by curcumin through induction of ROS. PLoS ONE 2013, 8, e57152. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Ye, J.; Alonso-Basanta, M.; Hahn, S.M.; Koumenis, C.; Dorsey, J.F. Modulation of CCAAT/enhancer binding protein homologous protein (CHOP) dependent DR5 expression by nelfinavir sensitizes glioblastoma multiforme cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). J. Biol. Chem. 2011, 286, 29408–29416. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Wang, H.G. CHOP is involved in endoplasmic reticulum stress-induced apoptosis by enhancing DR5 expression in human carcinoma cells. J. Biol. Chem. 2004, 279, 45495–45502. [Google Scholar] [CrossRef] [PubMed]
- Sung, B.; Prasad, S.; Ravindran, J.; Yadav, V.R.; Aggarwal, B.B. Capsazepine, a TRPV1 antagonist, sensitizes colorectal cancer cells to apoptosis by TRAIL through ROS-JNK-CHOP-mediated upregulation of death receptors. Free Radic. Biol. Med. 2012, 53, 1977–1987. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, E.J.; Yoon, Y.J.; Heo, J.; Lee, T.H.; Kim, Y.-H. Ciprofloxacin Enhances TRAIL-Induced Apoptosis in Lung Cancer Cells by Upregulating the Expression and Protein Stability of Death Receptors through CHOP Expression. Int. J. Mol. Sci. 2018, 19, 3187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103187
Lim EJ, Yoon YJ, Heo J, Lee TH, Kim Y-H. Ciprofloxacin Enhances TRAIL-Induced Apoptosis in Lung Cancer Cells by Upregulating the Expression and Protein Stability of Death Receptors through CHOP Expression. International Journal of Molecular Sciences. 2018; 19(10):3187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103187
Chicago/Turabian StyleLim, Eun Jin, Yu Jeong Yoon, Jeonghoon Heo, Tae Hwa Lee, and Young-Ho Kim. 2018. "Ciprofloxacin Enhances TRAIL-Induced Apoptosis in Lung Cancer Cells by Upregulating the Expression and Protein Stability of Death Receptors through CHOP Expression" International Journal of Molecular Sciences 19, no. 10: 3187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103187