The Novel miRNA N-72 Regulates EGF-Induced Migration of Human Amnion Mesenchymal Stem Cells by Targeting MMP2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
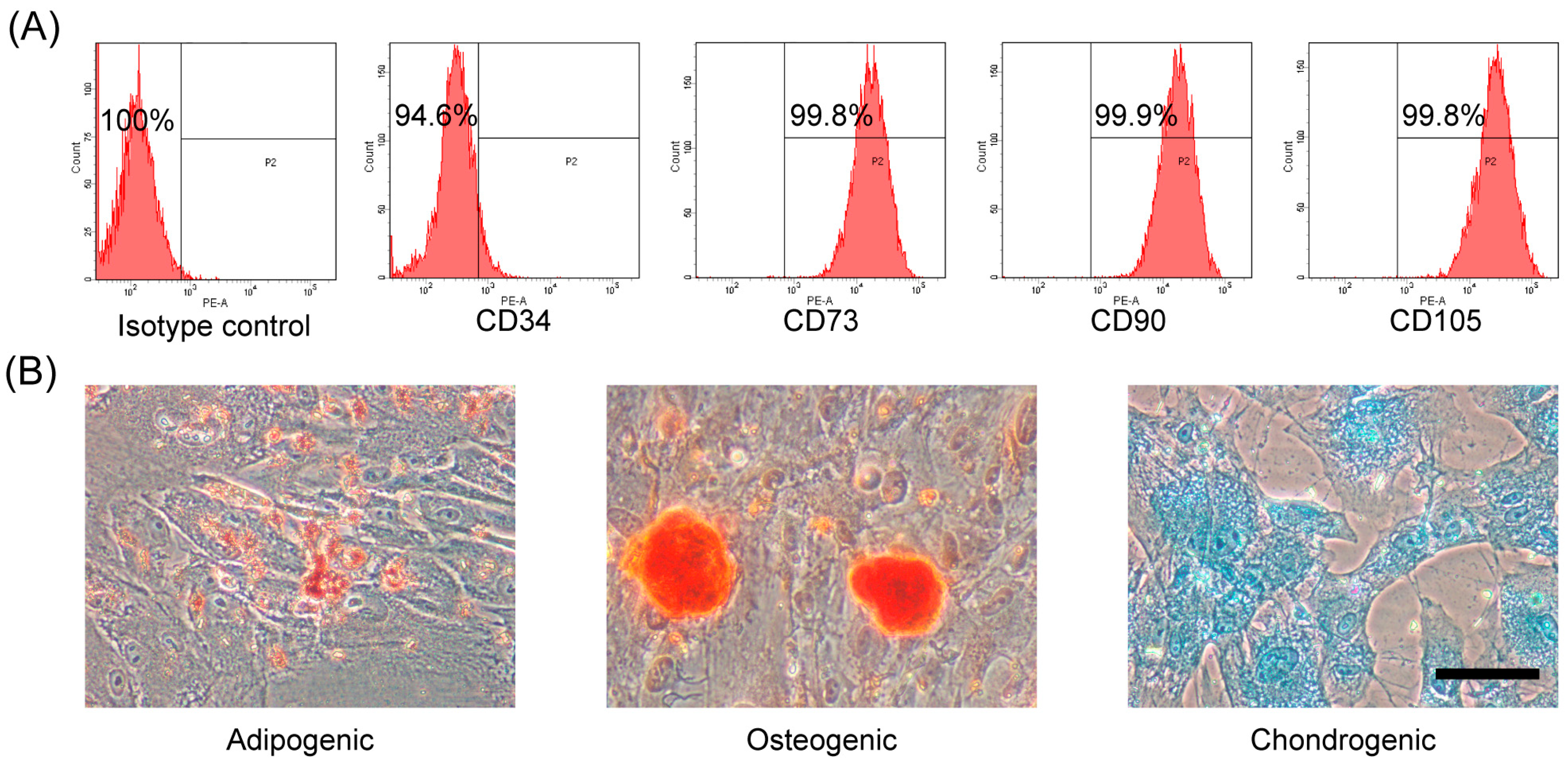
2.1. Characterization and Differentiation Capacity of hAMSCs
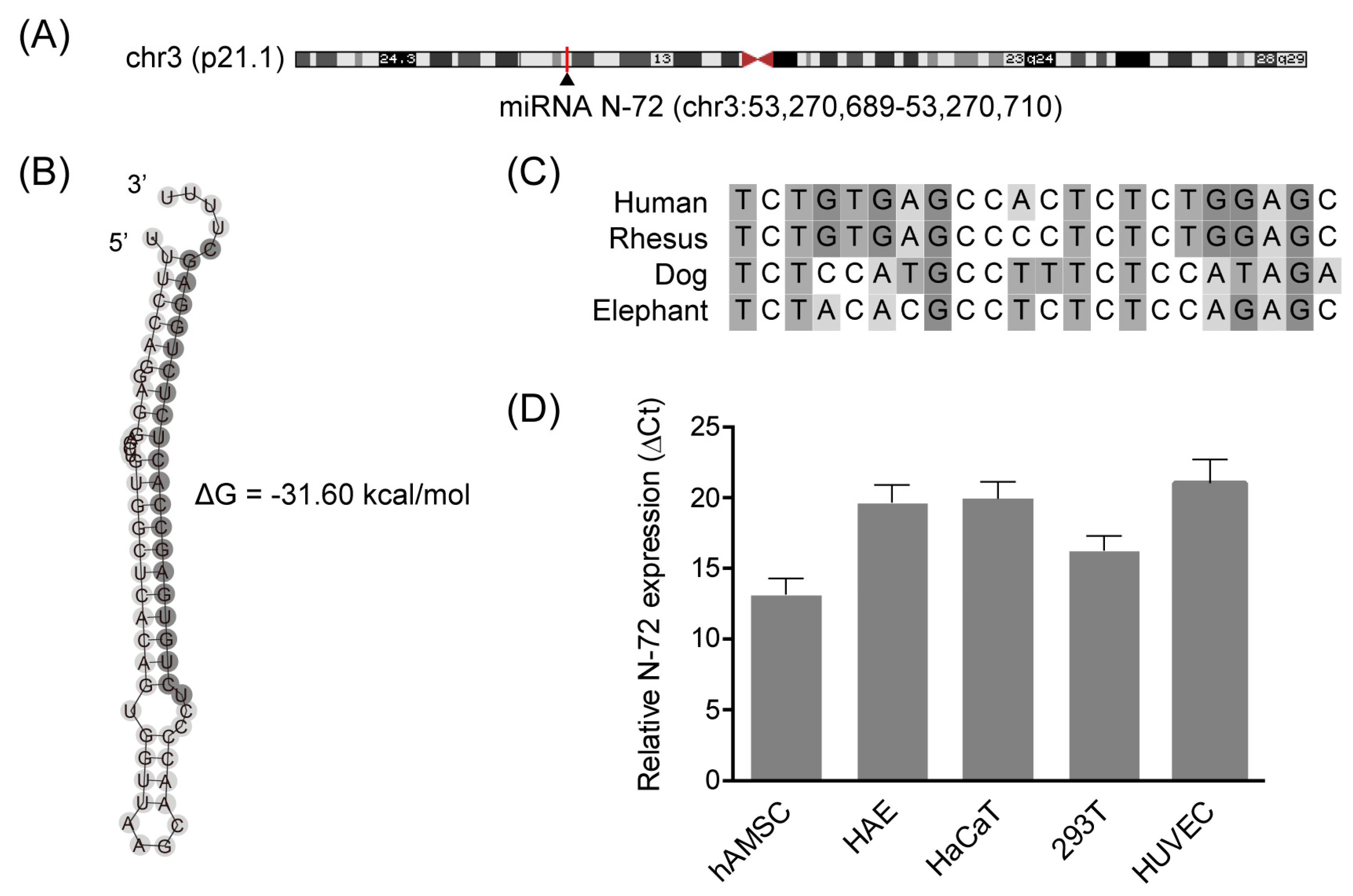
2.2. Discovery of the Novel miRNA N-72
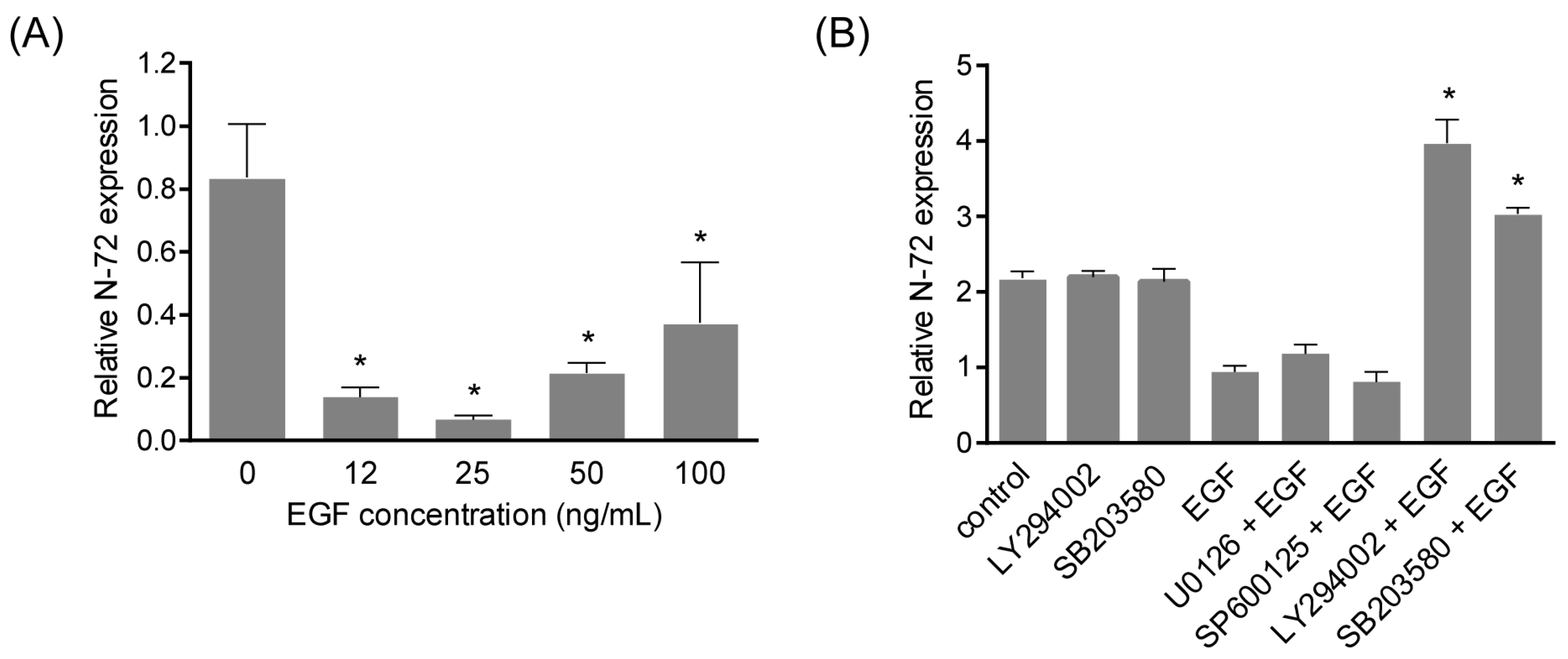
2.3. N-72 Was Downregulated by EGF Treatment
2.4. N-72 Inhibited EGF-Induced Cell Migration
2.5. MMP2 Was a Direct Target of N-72
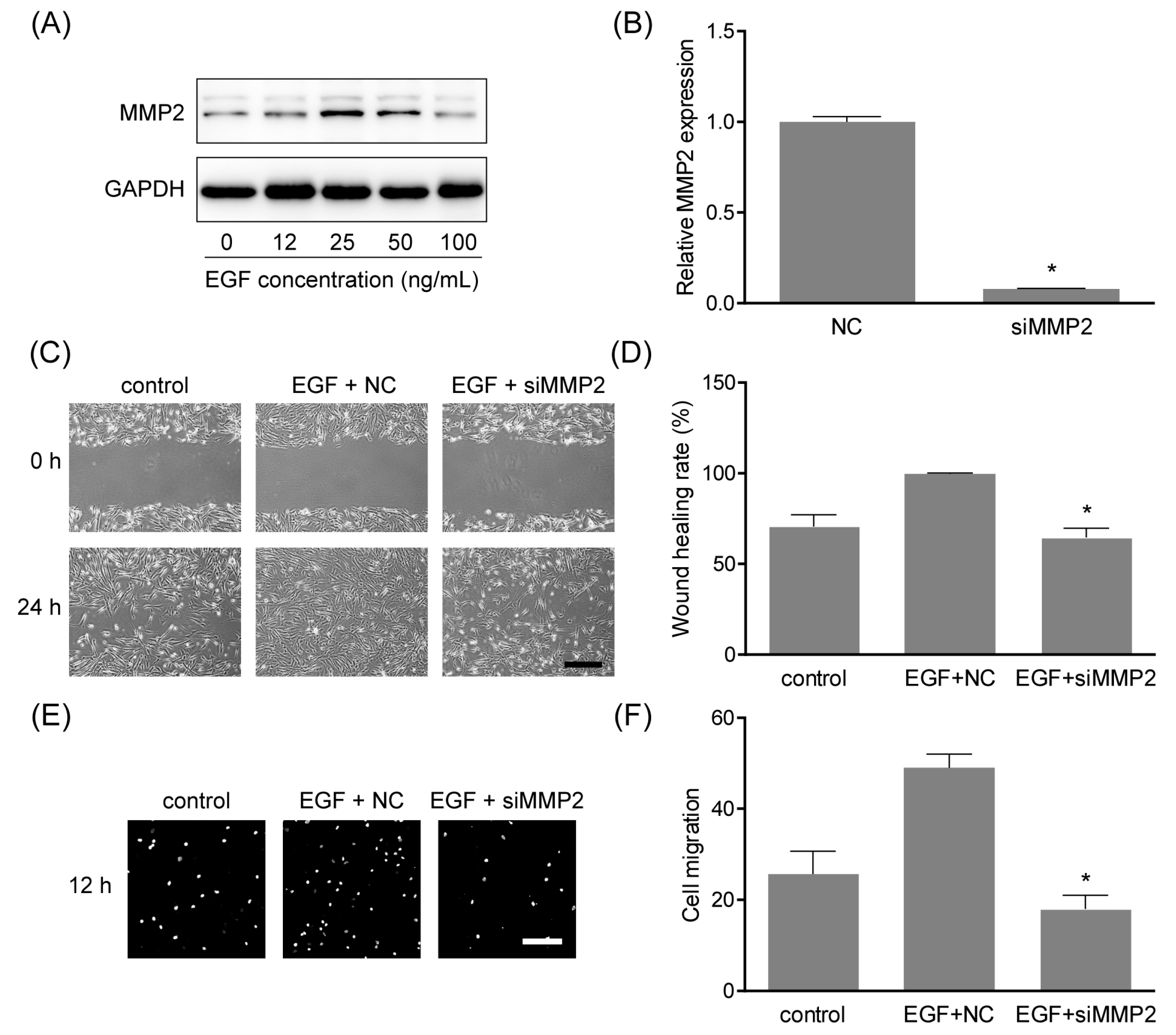
2.6. MMP2 Was Involved in EGF-Induced Cell Migration
3. Discussion
4. Materials and Methods
4.1. hAMSC Isolation and Cell Culture
4.2. Flow Cytometry Analysis
4.3. Trilineage Differentiation in Vitro
4.4. Bioinformatic Analysis
4.5. Quantitative Real-Time PCR
4.6. miRNA and siRNA Transfection
4.7. Transwell Migration Assay
4.8. In Vitro Scratch Assay
4.9. Dual-luciferase Reporter Assay
4.10. Western Blotting
4.11. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Li, C.Y.; Wu, X.Y.; Tong, J.B.; Yang, X.X.; Zhao, J.L.; Zheng, Q.F.; Zhao, G.B.; Ma, Z.J. Comparative analysis of human mesenchymal stem cells from bone marrow and adipose tissue under xeno-free conditions for cell therapy. Stem Cell Res. Ther. 2015, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.B.; Wang, J.; Wang, Z.; Wang, R.; Song, L.; Zhang, T.; Lin, X.W.; Shi, P.; Xin, H.C.; Pang, X.N. Polyethyleneimine-coated Fe3O4 nanoparticles for efficient sirna delivery to human mesenchymal stem cells derived from different tissues. Sci. Adv. Mater. 2015, 7, 1058–1064. [Google Scholar] [CrossRef]
- Le Blanc, K.; Davies, L.C. Mscs-cells with many sides. Cytotherapy 2018, 20, 273–278. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wang, X.; Kang, N.; Xu, J.; Dai, N.; Xu, X.; Zhang, H. Mir-375 inhibits the hepatocyte growth factor-elicited migration of mesenchymal stem cells by downregulating akt signaling. Cell Tissue Res. 2018, 372, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Nakazaki, M.; Sasaki, M.; Kataoka-Sasaki, Y.; Oka, S.; Namioka, T.; Namioka, A.; Onodera, R.; Suzuki, J.; Sasaki, Y.; Nagahama, H.; et al. Intravenous infusion of mesenchymal stem cells inhibits intracranial hemorrhage after recombinant tissue plasminogen activator therapy for transient middle cerebral artery occlusion in rats. J. Neurosurg. 2017, 127, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Cerri, S.; Greco, R.; Levandis, G.; Ghezzi, C.; Mangione, A.S.; Fuzzati-Armentero, M.T.; Bonizzi, A.; Avanzini, M.A.; Maccario, R.; Blandini, F. Intracarotid infusion of mesenchymal stem cells in an animal model of parkinson’s disease, focusing on cell distribution and neuroprotective and behavioral effects. Stem Cells Transl. Med. 2015, 4, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Yang, C.; Yang, P. The promotional effect of mesenchymal stem cell homing on bone tissue regeneration. Curr. Stem Cell Res. Ther. 2017, 12, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Syva, S.H.; Ampon, K.; Lasimbang, H.; Fatimah, S.S. Microenvironmental factors involved in human amnion mesenchymal stem cells fate decisions. J. Tissue Eng. Regen. Med. 2017, 11, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Jonas, S.; Izaurralde, E. Towards a molecular understanding of microrna-mediated gene silencing. Nat. Rev. Genet. 2015, 16, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.T.; Ye, H.; Huang, G.M.; Luo, F.; Liu, Y.W.; Liu, Y.; Yang, X.J.; Shen, J.; Liu, Q.Z.; Zhang, J.P. Long noncoding RNA GAS5 suppresses the migration and invasion of hepatocellular carcinoma cells via miR-21. Tumor Biol. 2016, 37, 2691–2702. [Google Scholar] [CrossRef] [PubMed]
- Han, J.J.; Kim, H.J.; Schafer, S.T.; Paquola, A.; Clemenson, G.D.; Toda, T.; Oh, J.S.; Pankonin, A.R.; Lee, B.S.; Johnston, S.T.; et al. Functional implications of miR-19 in the migration of newborn neurons in the adult brain. Neuron 2016, 91, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chen, P.; Wu, Y.; Wang, K.; Xu, Y.; Chen, H.; Zhang, L.; Wu, R.; Webster, K.A.; Yu, H.; et al. MiR-211/STAT5A signaling modulates migration of mesenchymal stem cells to improve its therapeutic efficacy. Stem Cells 2016, 34, 1846–1858. [Google Scholar] [CrossRef] [PubMed]
- Tome, M.; Lopez-Romero, P.; Albo, C.; Sepulveda, J.C.; Fernandez-Gutierrez, B.; Dopazo, A.; Bernad, A.; Gonzalez, M.A. MiR-335 orchestrates cell proliferation, migration and differentiation in human mesenchymal stem cells. Cell Death Differ. 2011, 18, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, K.; Mohan, S.S.; Raju, R.; Keerthikumar, S.; Kumar, G.S.; Venugopal, A.K.; Telikicherla, D.; Navarro, J.D.; Mathivanan, S.; Pecquet, C.; et al. Netpath: A public resource of curated signal transduction pathways. Genome Biol. 2010, 11, R3. [Google Scholar] [CrossRef] [PubMed]
- Nitzsche, F.; Muller, C.; Lukomska, B.; Jolkkonen, J.; Deten, A.; Boltze, J. Concise review: MSC adhesion cascade-insights into homing and transendothelial migration. Stem Cells 2017, 35, 1446–1460. [Google Scholar] [CrossRef] [PubMed]
- De Becker, A.; Van Hummelen, P.; Bakkus, M.; Vande Broek, I.; De Wever, J.; De Waele, M.; Van Riet, I. Migration of culture-expanded human mesenchymal stem cells through bone marrow endothelium is regulated by matrix metalloproteinase-2 and tissue inhibitor of metalloproteinase-3. Haematologica 2007, 92, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R.I. Microrna biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microrna biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wu, B.; Jia, S.; Zhao, Y.; Hou, R.; Liu, X.; Wang, X.; Chen, L.; Yang, X.; Lei, D.; et al. The mechanically activated p38/mmp-2 signaling pathway promotes bone marrow mesenchymal stem cell migration in rats. Arch. Oral Biol. 2017, 76, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Vaghjiani, V.; Cain, J.E.; Lee, W.; Vaithilingam, V.; Tuch, B.E.; St John, J.C. Modulation of mitochondrial DNA copy number to induce hepatocytic differentiation of human amniotic epithelial cells. Stem Cells Dev. 2017, 26, 1505–1519. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.; Bernhart, S.H.; Honer Zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. Viennarna package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.B.; Wang, J.; Wang, Z.; Zhang, T.; Shi, P.; Wang, X.L.; Zhao, F.; Liu, X.Y.; Lin, X.W.; Pang, X.N. MiR-136 modulates TGF-β1-induced proliferation arrest by targeting PPP2R2A in keratinocytes. Biomed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhang, D.; Chen, M.; Wang, R.; Zhang, T.; Zhao, F.; Lin, X.; Pang, X. The Novel miRNA N-72 Regulates EGF-Induced Migration of Human Amnion Mesenchymal Stem Cells by Targeting MMP2. Int. J. Mol. Sci. 2018, 19, 1363. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051363
Li Y, Zhang D, Chen M, Wang R, Zhang T, Zhao F, Lin X, Pang X. The Novel miRNA N-72 Regulates EGF-Induced Migration of Human Amnion Mesenchymal Stem Cells by Targeting MMP2. International Journal of Molecular Sciences. 2018; 19(5):1363. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051363
Chicago/Turabian StyleLi, Ying, Dianbao Zhang, Meng Chen, Rui Wang, Tao Zhang, Feng Zhao, Xuewen Lin, and Xining Pang. 2018. "The Novel miRNA N-72 Regulates EGF-Induced Migration of Human Amnion Mesenchymal Stem Cells by Targeting MMP2" International Journal of Molecular Sciences 19, no. 5: 1363. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051363