Novel Stenotic Microchannels to Study Thrombus Formation in Shear Gradients: Influence of Shear Forces and Human Platelet-Related Factors

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
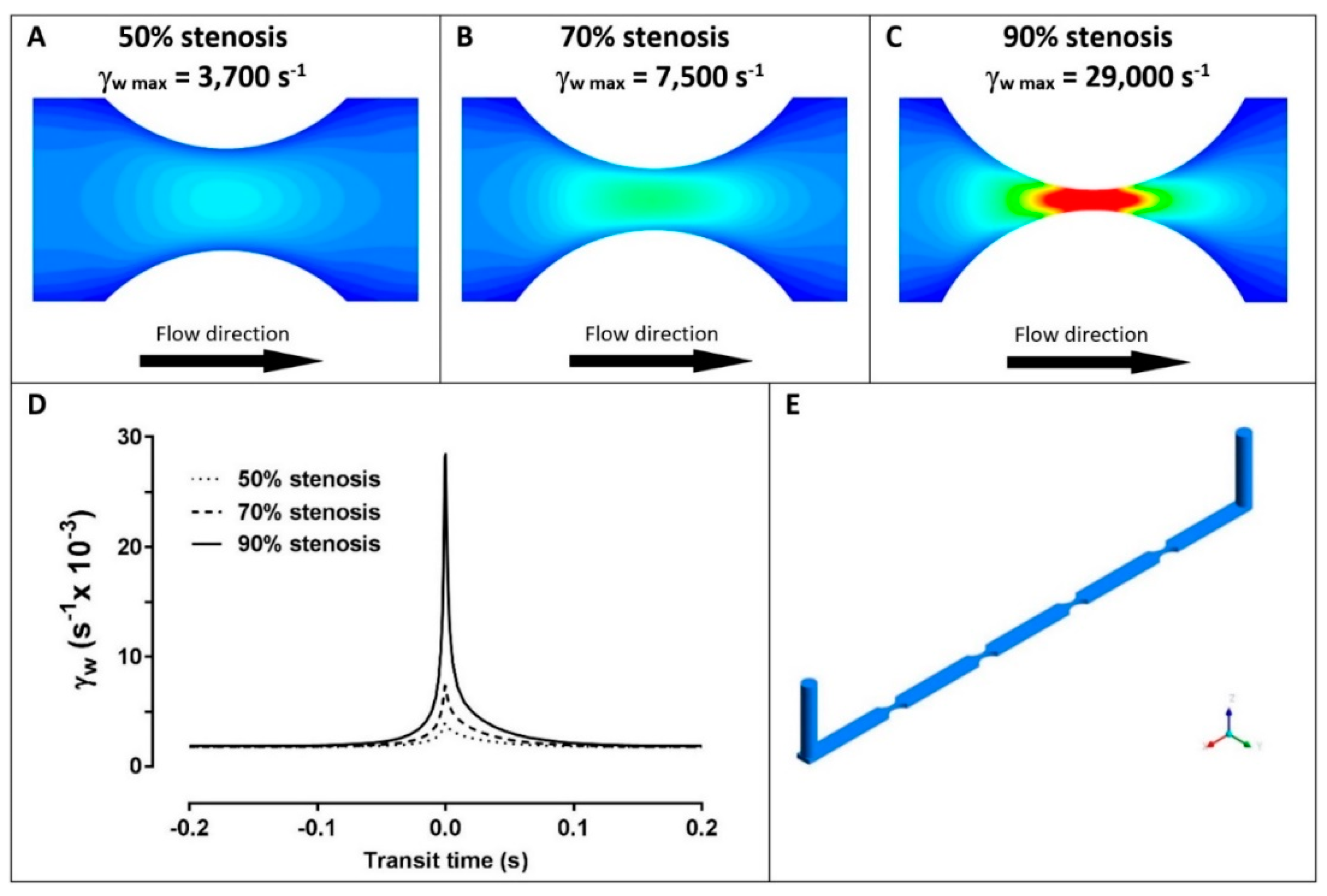
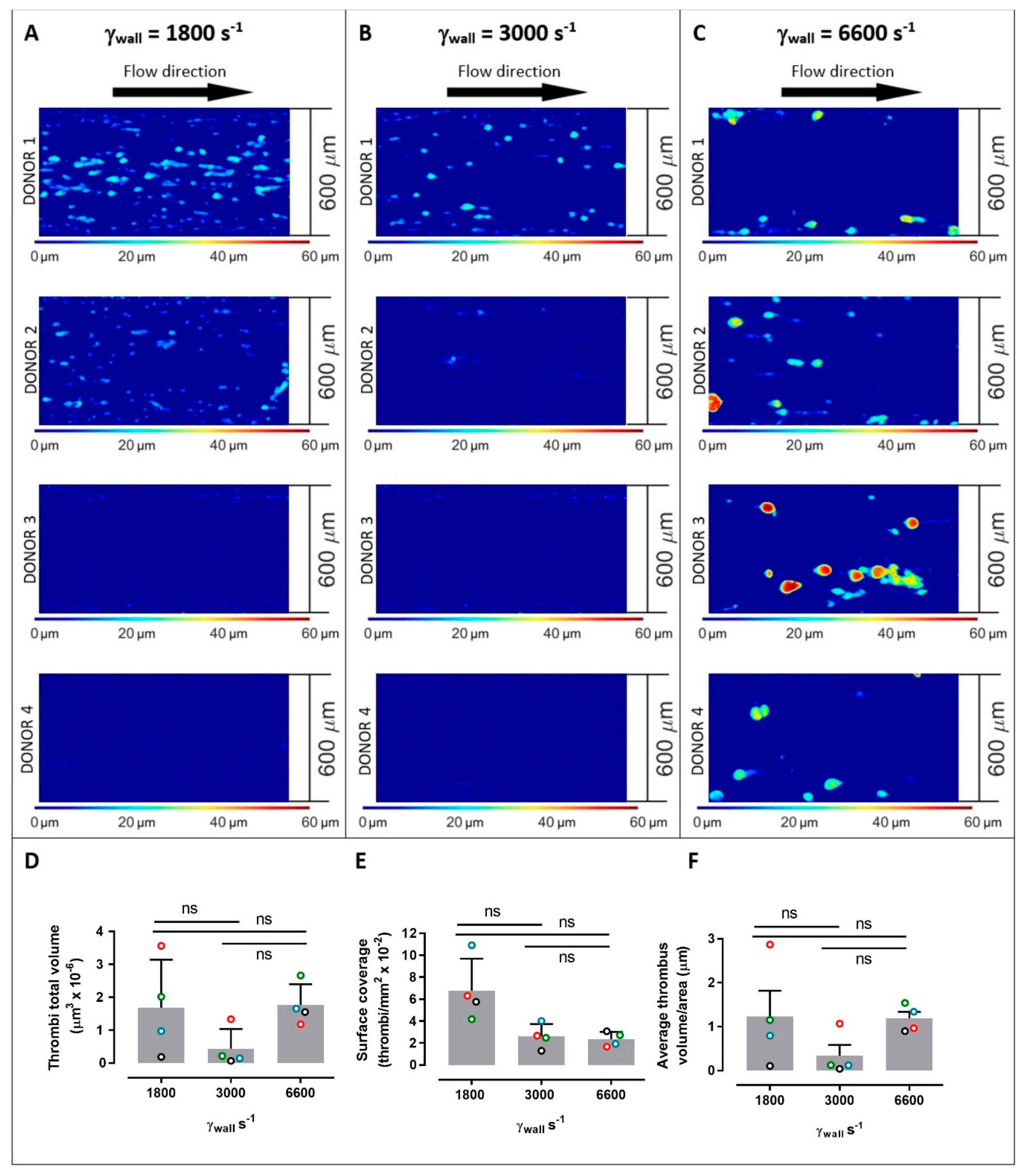
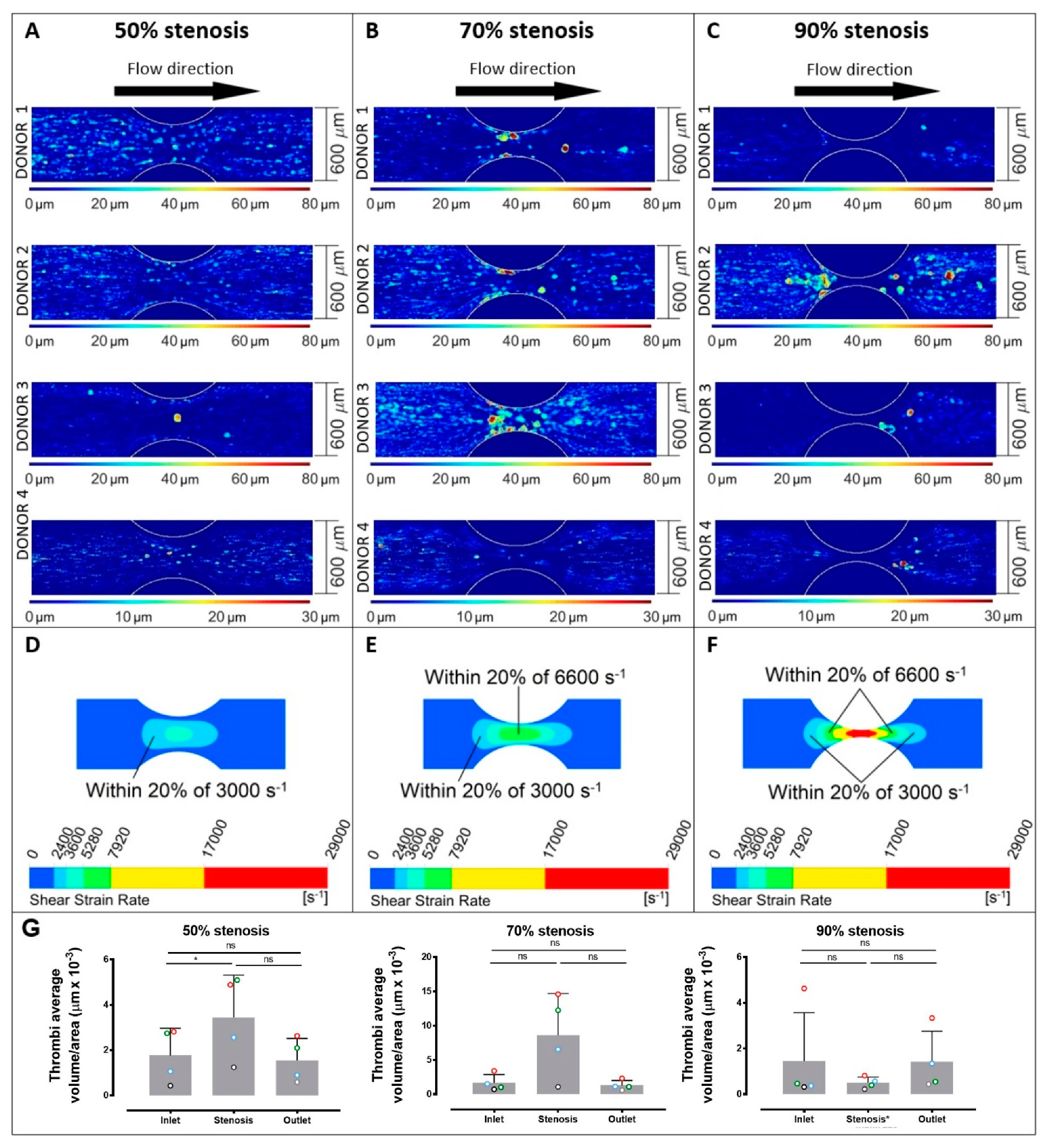
2. Results
3. Discussion
4. Materials and Methods
4.1. Blood Collection and Analysis
4.2. Fabrication of the Stenosed Microchannels
4.3. Calculations of the Stenosis Percentages
4.4. Platelet Thrombus Formation under Flow
4.5. Analysis of Thrombus Formation
4.6. Shear Force Equations and Units
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Al-Tamimi, M.; Tan, C.W.; Qiao, J.; Pennings, G.J.; Javadzadegan, A.; Yong, A.S.C.; Arthur, J.F.; Davis, A.K.; Jing, J.; Mu, F.-T.; et al. Pathological shear triggers shedding of vascular receptors: A novel mechanism for downregulation of platelet glycoprotein (GP)VI in stenosed coronary vessels. Blood 2012, 119, 4311–4320. [Google Scholar] [CrossRef]
- Gardiner, E.E.; Andrews, R.K. Platelet adhesion. In Platelets in Thrombotic and Non-Thrombotic Disorders; Gresele, P., Lopez, J.A., Kleiman, N.S., Page, C.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 309–319. [Google Scholar]
- Maxwell, M.J.; Westein, E.; Nesbitt, W.S.; Giuliano, S.; Dopheide, S.M.; Jackson, S.P. Identification of a 2-stage platelet aggregation process mediating shear-dependent thrombus formation. Blood 2007, 109, 566–576. [Google Scholar] [CrossRef]
- Nesbitt, W.S.; Westein, E.; Tovar-Lopez, F.J.; Tolouei, E.; Mitchell, A.; Fu, J.; Carberry, J.; Fouras, A.; Jackson, S.P. A shear gradient-dependent platelet aggregation mechanism drives thrombus formation. Nat. Med. 2009, 15, 665–673. [Google Scholar] [CrossRef]
- Shi, X.; Yang, J.; Huang, J.; Long, Z.; Ruan, Z.; Xiao, B.; Xi, X. Effects of different shear rates on the attachment and detachment of platelet thrombi. Mol. Med. Rep. 2016, 13, 2447–2456. [Google Scholar] [CrossRef]
- Chopard, B.; de Sousa, D.R.; Lätt, J.; Mountrakis, L.; Dubois, F.; Yourassowsky, C.; Van Antwerpen, P.; Eker, O.; Vanhamme, L.; Perez-Morga, D.; et al. A physical description of the adhesion and aggregation of platelets. R. Soc. Open Sci. 2017, 4, 170219. [Google Scholar] [CrossRef] [Green Version]
- Koltai, K.; Kesmarky, G.; Feher, G.; Tibold, A.; Toth, K. Platelet aggregometry testing: Molecular mechanisms, techniques and clinical implications. Int. J. Mol. Sci. 2017, 18, 1803. [Google Scholar] [CrossRef]
- Li, R.; Grosser, T.; Diamond, S.L. Microfluidic whole blood testing of platelet response to pharmacological agents. Platelets 2017, 28, 457–462. [Google Scholar] [CrossRef]
- Qiu, Y.; Ciciliano, J.; Myers, D.R.; Tran, R.; Lam, W.A. Platelets and physics: How platelets “feel” and respond to their mechanical microenvironment. Blood Rev. 2015, 29, 377–386. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, L.; Slepian, M.J.; Deng, Y.; Bluestein, D. A multiscale biomechanical model of platelets: Correlating with in-vitro results. J. Biomech. 2017, 50, 26–33. [Google Scholar] [CrossRef]
- Slepian, M.J.; Sheriff, J.; Hutchinson, M.; Tran, P.; Bajaj, N.; Garcia, J.G.; Scott Saavedra, S.; Bluestein, D. Shear-mediated platelet activation in the free flow: Perspectives on the emerging spectrum of cell mechanobiological mechanisms mediating cardiovascular implant thrombosis. J. Biomech. 2017, 50, 20–25. [Google Scholar] [CrossRef]
- Pinar, I.P.; Arthur, J.F.; Andrews, R.K.; Gardiner, E.E.; Ryan, K.; Carberry, J. Methods to determine the Lagrangian shear experienced by platelets during thrombus growth. PlosONE 2015, 10, e0144860. [Google Scholar] [CrossRef]
- Gardiner, E.E.; Andrews, R.K. Platelet receptor expression and shedding: GPIb-IX-V and GPVI. Transfusion Med. Rev. 2014, 28, 56–60. [Google Scholar] [CrossRef]
- Andrews, R.K.; Berndt, M.C. Platelet adhesion: A game of catch and release. J. Clin. Invest. 2008, 118, 3009–3011. [Google Scholar] [CrossRef]
- Yago, T.; Lou, J.; Wu, T.; Yang, J.; Miner, J.J.; Coburn, L.; López, J.A.; Cruz, M.A.; Dong, J.F.; McIntire, L.V.; et al. Platelet glycoprotein Ibα forms catch bonds with human WT vWF but not with type 2B von Willebrand disease vWF. J. Clin. Invest. 2008, 118, 3195–3207. [Google Scholar] [CrossRef]
- Arthur, J.F.; Gardiner, E.E.; Matzaris, M.; Taylor, S.G.; Wijeyewickrema, L.; Ozaki, Y.; Kahn, M.L.; Andrews, R.K.; Berndt, M.C. Glycoprotein VI is associated with GPIb-IX-V on the membrane of resting and activated platelets. Thromb. Haemost. 2005, 93, 716–723. [Google Scholar]
- Kruss, S.; Erpenbeck, L.; Amschler, K.; Mundinger, T.A.; Boehm, H.; Helms, H.J.; Friede, T.; Andrews, R.K.; Schön, M.P.; Spatz, J.P. Adhesion maturation of neutrophils on nanoscopically presented platelet glycoprotein Ibα. ACS Nano 2013, 7, 9984–9996. [Google Scholar] [CrossRef]
- Kamada, H.; Imai, Y.; Nakamura, M.; Ishikawa, T.; Yamaguchi, T. Computational study on thrombus formation regulated by platelet glycoprotein and blood flow shear. Microvasc. Res. 2013, 89, 95–106. [Google Scholar] [CrossRef]
- Andrews, R.K.; Gardiner, E.E. Basic mechanisms of platelet receptor shedding. Platelets 2017, 28, 319–324. [Google Scholar] [CrossRef]
- Chen, Z.; Mondal, N.K.; Ding, J.; Gao, J.; Griffith, B.P.; Wu, Z.J. Shear-induced platelet receptor shedding by non-physiological high shear stress with short exposure time: Glycoprotein Ibα and glycoprotein VI. Thromb. Res. 2015, 135, 692–698. [Google Scholar] [CrossRef]
- Montague, S.J.; Andrews, R.K.; Gardiner, E.E. Mechanisms of receptor shedding in platelets. Blood 2018, 132, 2535–2545. [Google Scholar] [CrossRef] [Green Version]
- Lukito, P.; Wong, A.; Jing, J.; Mado, B.; Arthur, J.F.; Marasco, S.F.; Murphy, D.A.; Pellegrino, V.A.; Bergin, P.J.; Shaw, J.; et al. Mechanical circulatory support is associated with loss of platelet receptors glycoprotein (GP) Ibα and GPVI. J. Thromb. Haemost. 2016, 14, 2253–2260. [Google Scholar] [CrossRef]
- Muthiah, K.; Connor, D.; Ly, K.; Gardiner, E.E.; Andrews, R.K.; Qiao, J.; Rutgers, D.; Robson, D.; Low, J.; Jarvis, S.; et al. Longitudinal changes in haemostatic parameters and reduced pulsatility contribute to non-surgical bleeding in patients with centrifugal continuous flow left ventricular assist devices. J. Heart Lung Transpl. 2016, 35, 743–751. [Google Scholar] [CrossRef]
- Geffen, J.P.V.; Brouns, S.L.N.; Batista, J.; McKinney, H.; Kempster, C.; Nagy, M.; Sivapalaratnam, S.; Baaten, C.C.F.M.J.; Bourry, N.; Frontini, M.; et al. High-throughput elucidation of thrombus formation reveals sources of platelet function variability. Haematologica. 2018. [Google Scholar] [CrossRef]
- Chan, P.; Cheng, S.H.; Poon, T.C. Automated segmentation in confocal images using a density clustering method. J. Electron Imaging 2007, 16, 043003. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lui, M.; Gardiner, E.E.; Arthur, J.F.; Pinar, I.; Lee, W.M.; Ryan, K.; Carberry, J.; Andrews, R.K. Novel Stenotic Microchannels to Study Thrombus Formation in Shear Gradients: Influence of Shear Forces and Human Platelet-Related Factors. Int. J. Mol. Sci. 2019, 20, 2967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122967
Lui M, Gardiner EE, Arthur JF, Pinar I, Lee WM, Ryan K, Carberry J, Andrews RK. Novel Stenotic Microchannels to Study Thrombus Formation in Shear Gradients: Influence of Shear Forces and Human Platelet-Related Factors. International Journal of Molecular Sciences. 2019; 20(12):2967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122967
Chicago/Turabian StyleLui, Mathew, Elizabeth E. Gardiner, Jane F. Arthur, Isaac Pinar, Woei Ming Lee, Kris Ryan, Josie Carberry, and Robert K. Andrews. 2019. "Novel Stenotic Microchannels to Study Thrombus Formation in Shear Gradients: Influence of Shear Forces and Human Platelet-Related Factors" International Journal of Molecular Sciences 20, no. 12: 2967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122967