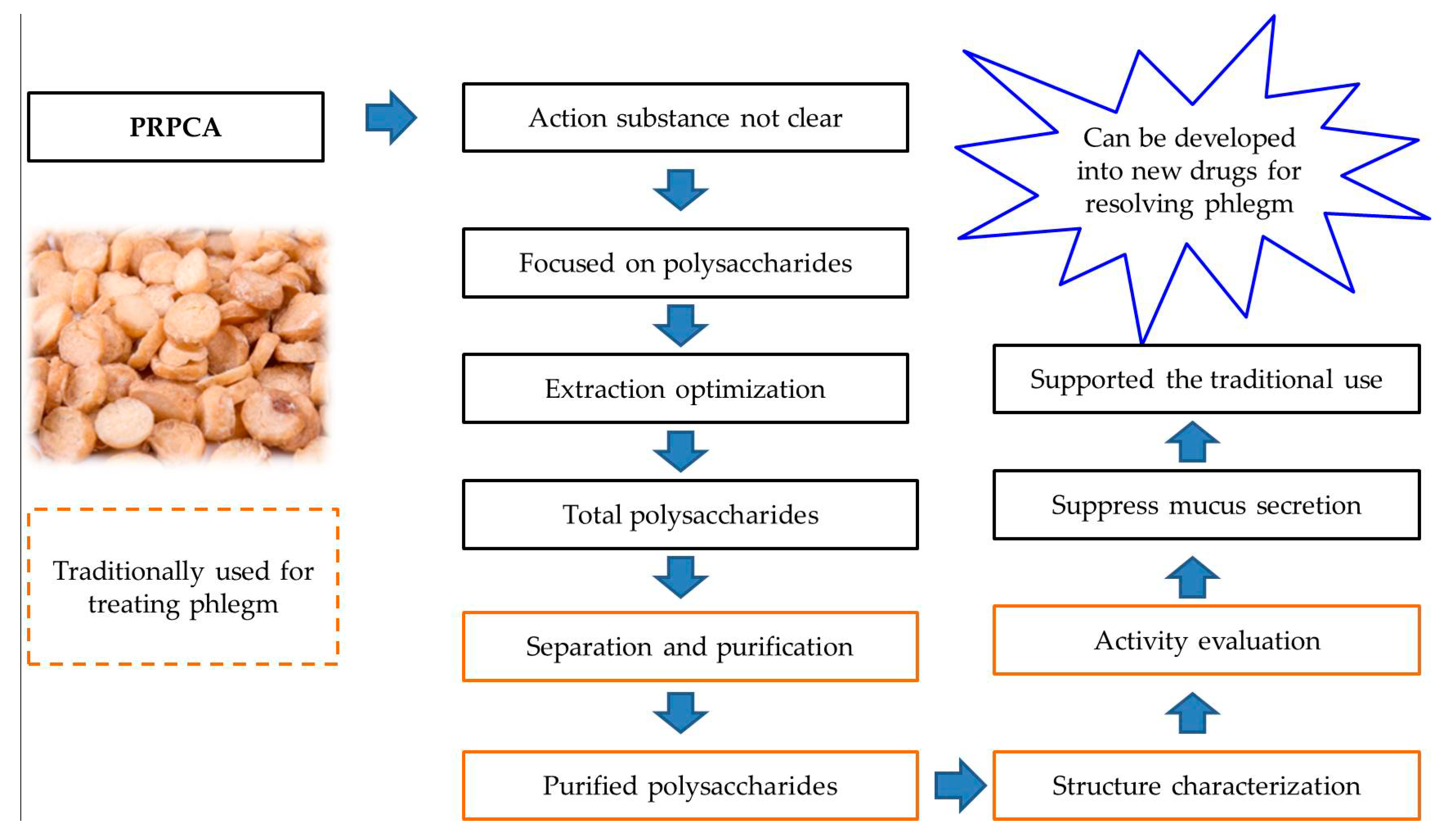
Purification, Characterization of Two Polysaccharides from Pinelliae Rhizoma Praeparatum Cum Alumine and Their Anti-Inflammatory Effects on Mucus Secretion of Airway Epithelium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
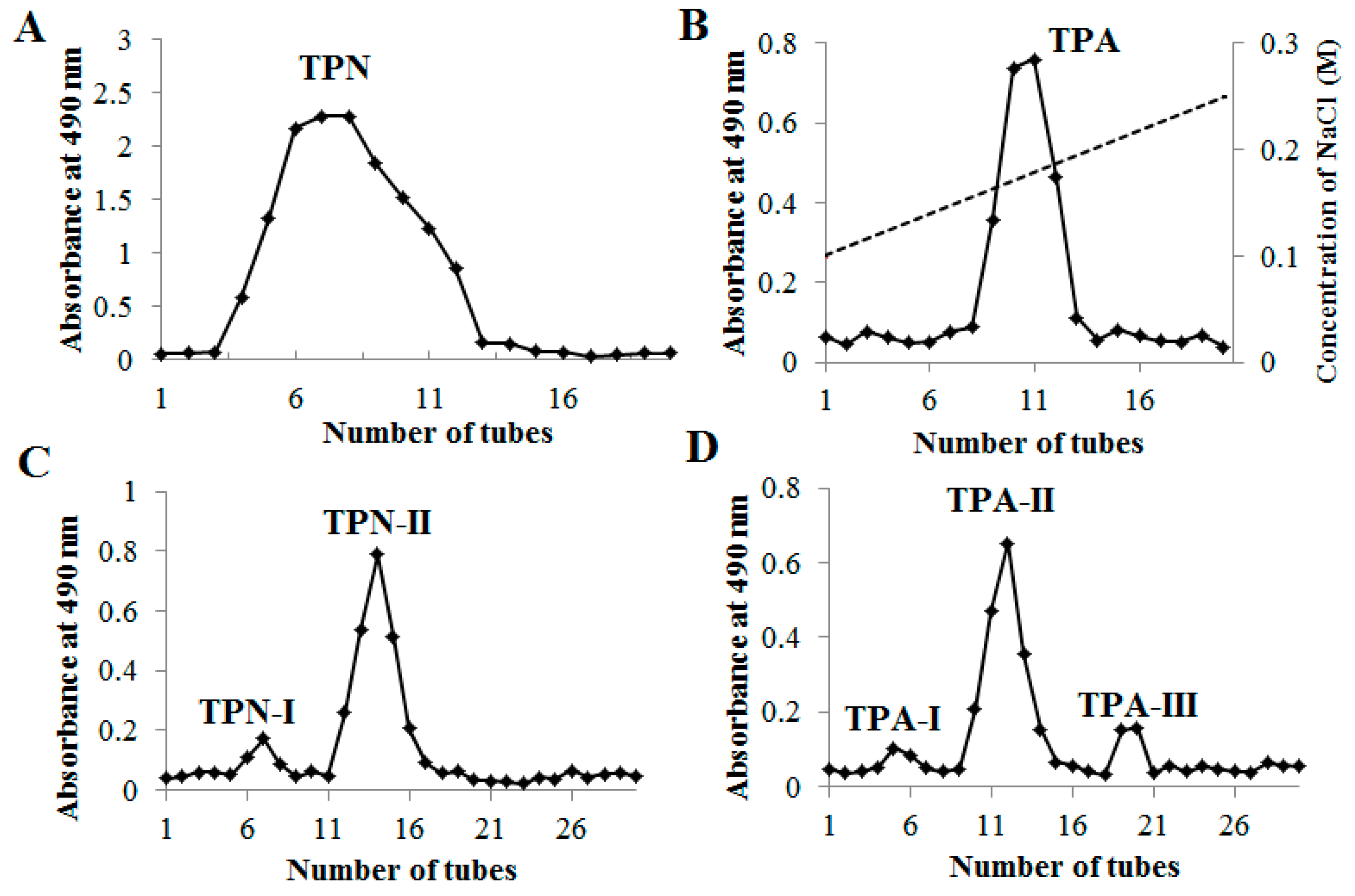
2.1. Purification of Polysaccharides from PRPCA
2.2. Morphological Properties
2.3. The Characterization of TPN-II and TPA-II
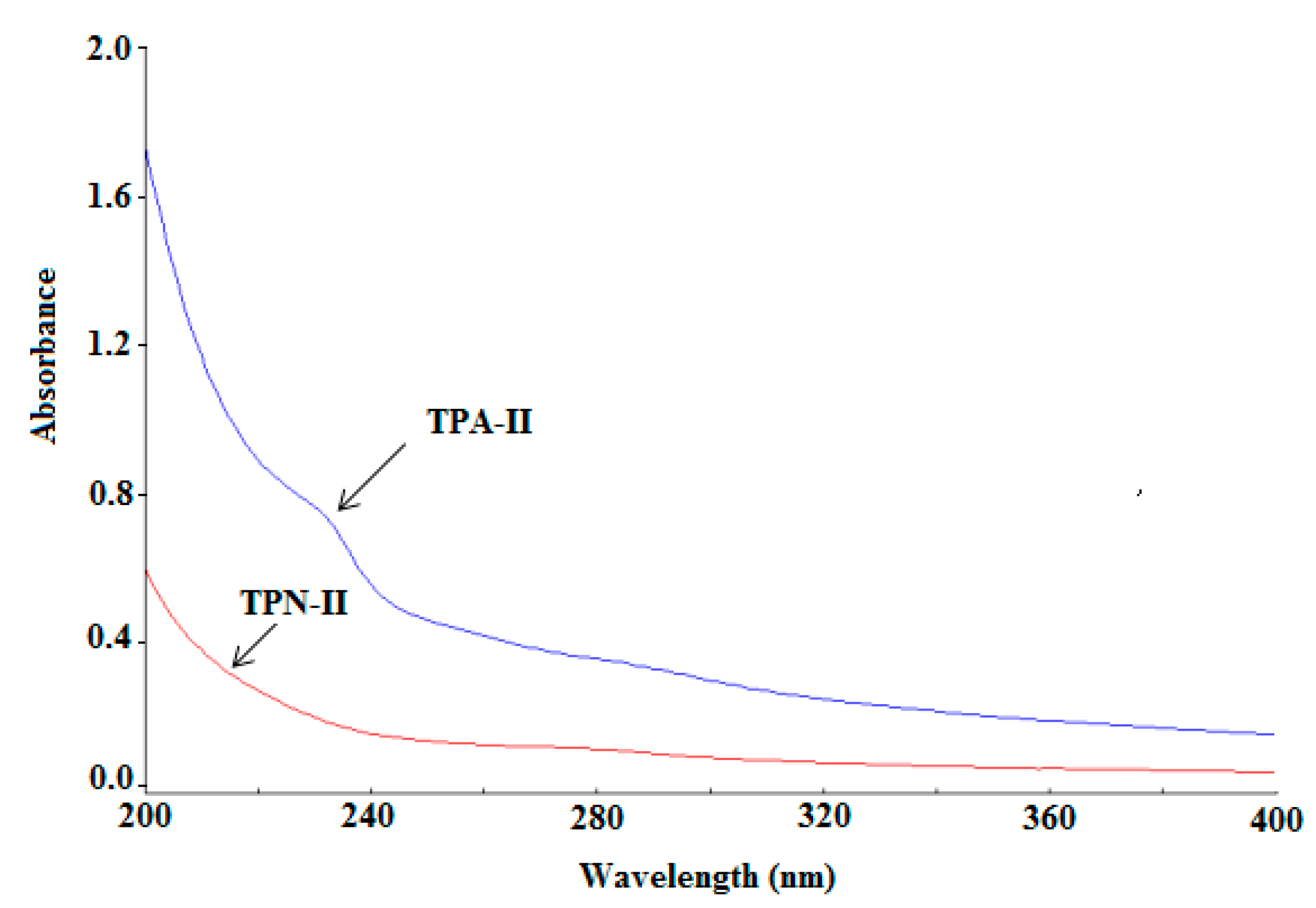
2.3.1. Ultraviolet (UV) Analysis
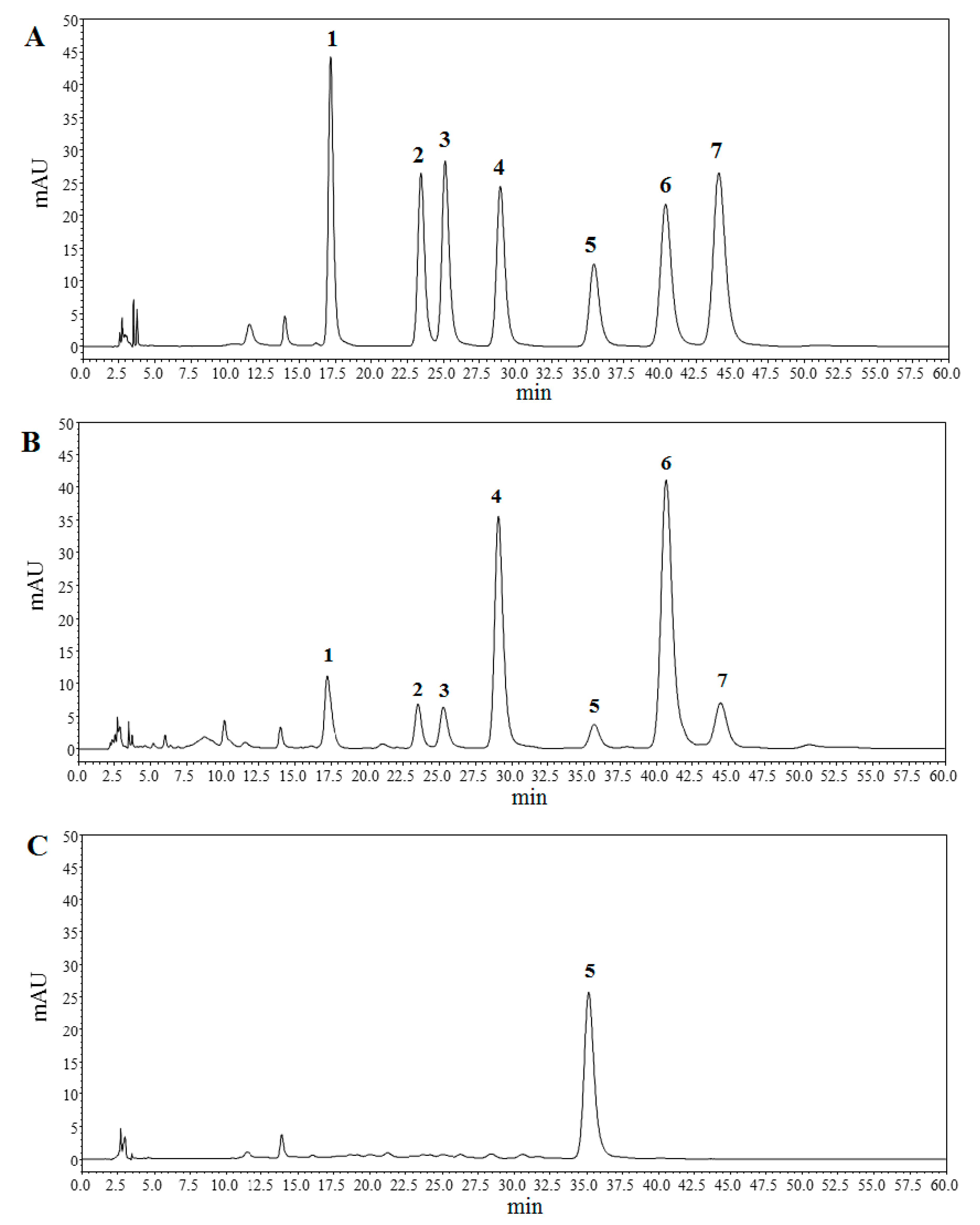
2.3.2. Molecular Weight and Monosaccharide Composition
2.4. Anti-Inflammatory Effects on Mucus Secretion
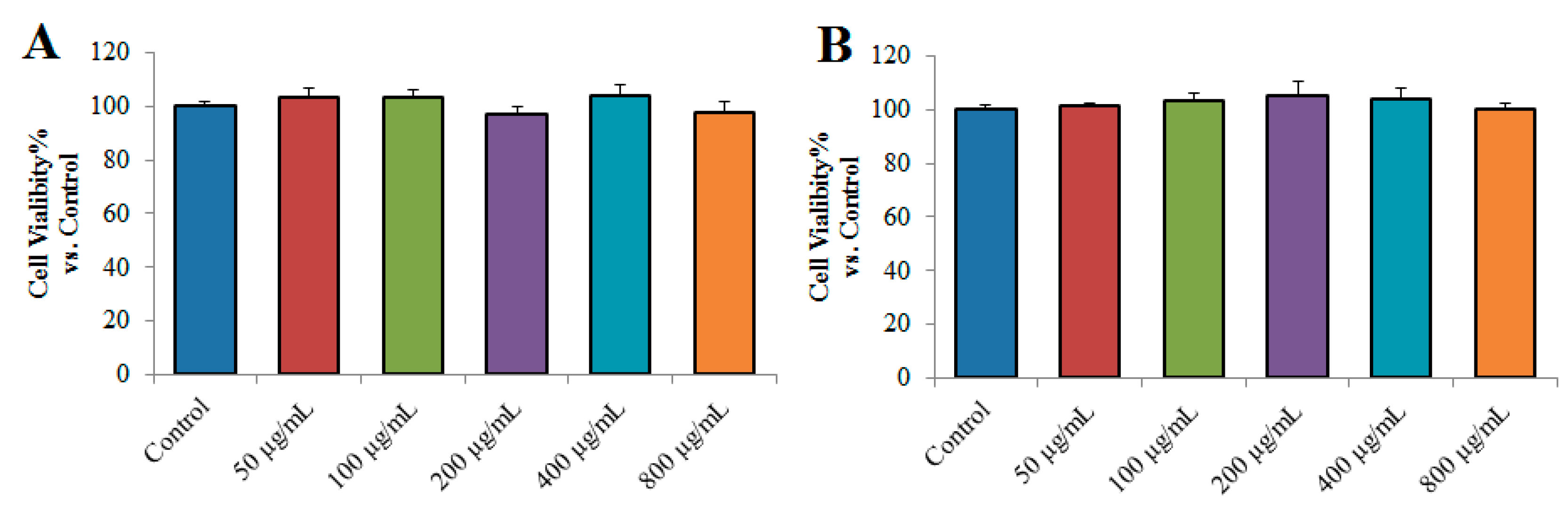
2.4.1. Effects of TPN-II and TPA-II on the Viability of NCI-H292 Cells
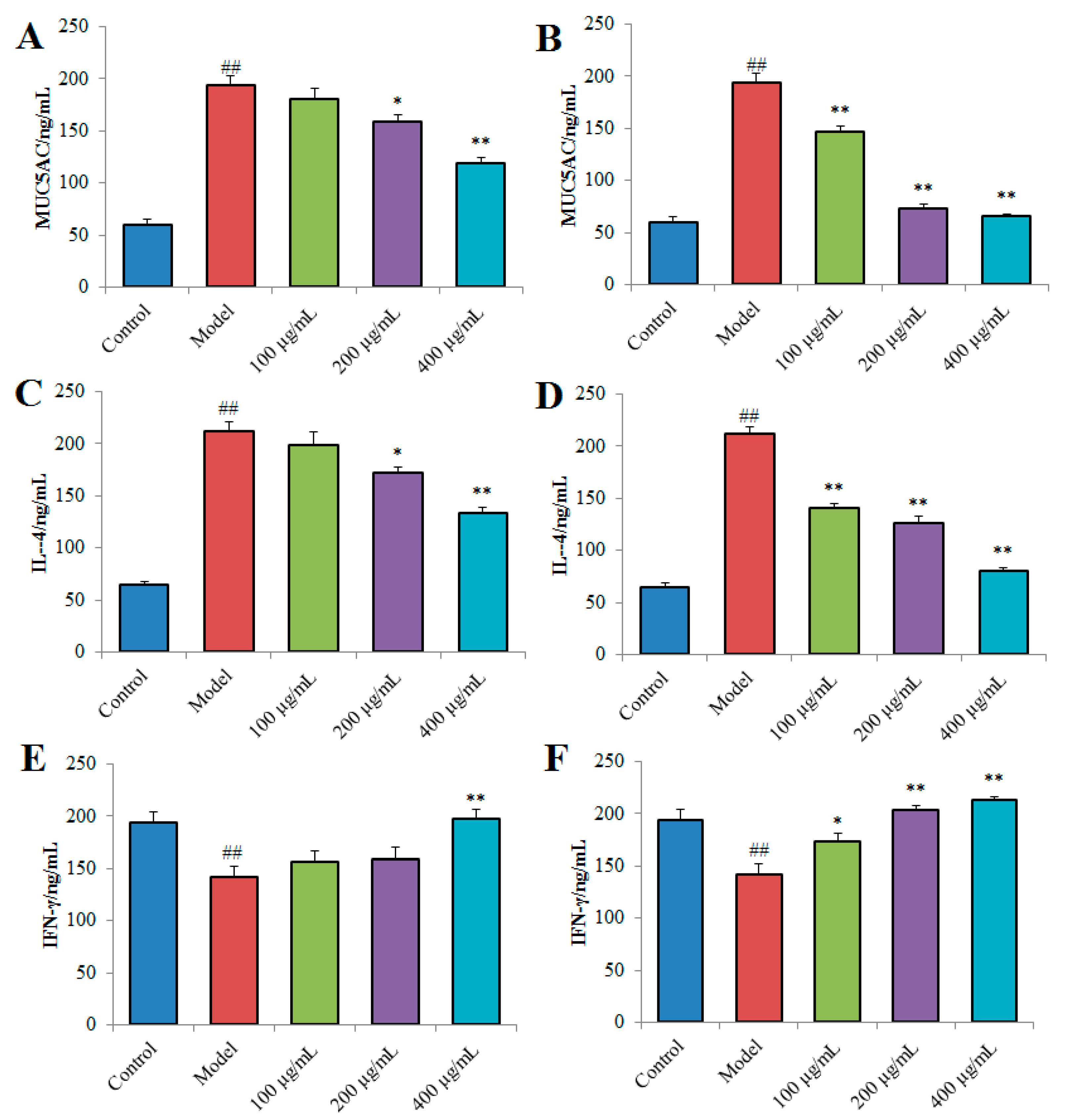
2.4.2. Effects of TPN-II and TPA-II on Overproduction of MUC5AC, interleukin-4 (IL-4) and interferon-γ (IFN-γ) in NCI-H292 cells
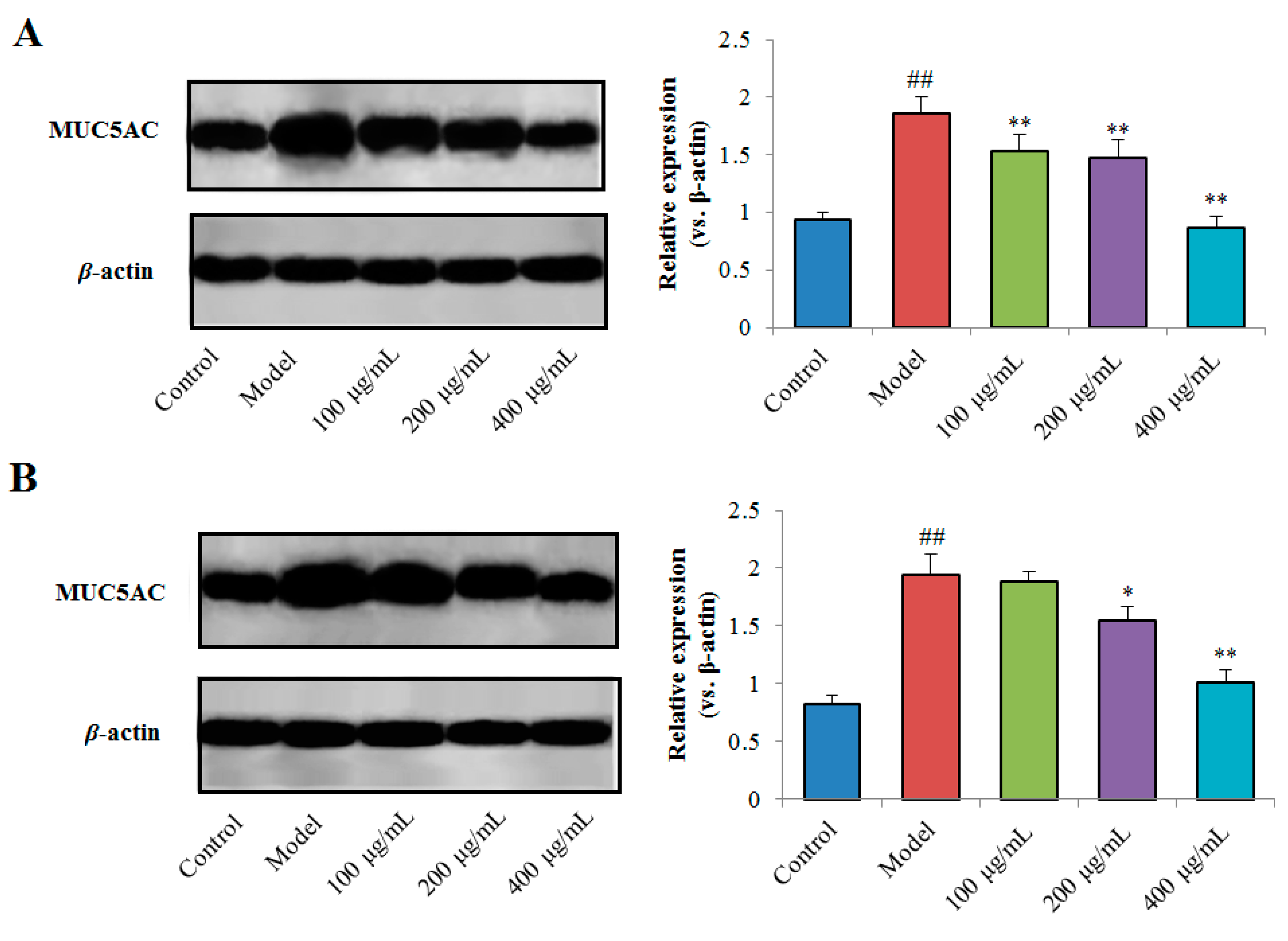
2.4.3. Effects of TPN-II and TPA-II on Expression of MUC5AC Protein
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Extraction and Purification of Polysaccharides from PRPCA
4.3. Characterization Analysis of TPN-II and TPA-II
4.3.1. SEM Analysis
4.3.2. UV and FT-IR Spectrometric Analysis
4.3.3. Molecular Weight Determination and Monosaccharide Composition Analysis
4.3.4. NMR Spectroscopy
4.4. Anti-Inflammatory Effects on Mucus Secretion of NCI-H292 Cells
4.4.1. Cell Culture and Treatments
4.4.2. ELISA for MUC5AC, IL-4 and IFN-γ
4.4.3. Western Blot Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Ara | Arabinose |
| D2O | Deuteroxide |
| DEAE-52 | Diethylaminoethyl Cellulose 52 |
| ELISA | Enzyme-linked immunosorbent assay |
| Gal | Galactose |
| FT-IR | Fourier transform-infrared spectroscopy |
| GalA | Galacturonic acid |
| Glu | Glucose |
| GluA | Glucuronic acid |
| HPGPC | High performance gel permeation chromatography |
| HPLC | High performance liquid chromatography |
| IFN-γ | Interferon-γ |
| IgE | Immunoglobulin E |
| IL-4 | Interleukin-4 |
| LD50 | Lethal dose for 50% |
| LPS | Lipopolysaccharide |
| Man | Mannose |
| MUC2 | Mucin-2 |
| MUC5AC | Mucin-5 subtype AC |
| NMR | Nuclear magnetic resonance |
| PRPCA | Pinelliae Rhizoma Praeparatum cum Alumine |
| PVDF | Polyvinylidene fluoride |
| Rha | Rhamnose |
| S.D. | Standard deviation |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SPSS | Statistical Product and Service Solutions |
| SEM | Scanning electron microscopy |
| Th1 | Helper T 1 |
| Th2 | Helper T 2 |
| TNF-α | Tumor necrosis factor-α |
| TPA | Acidic fraction of total polysaccharides |
| TPN | Neutral fraction of total polysaccharides |
| UV | Ultraviolet |
| WPMP | Polysaccharides of Polygonum multiflorum |
References
- He, X.; Wang, X.; Fang, J.; Chang, Y.; Ning, N.; Guo, H.; Huang, L.; Huang, X.; Zhao, Z. Structures, biological activities, and industrial applications of the polysaccharides from Hericium erinaceus (Lion’s Mane) mushroom: A review. Int. J. Biol. Macromol. 2017, 97, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Minzanova, S.T.; Mironov, V.F.; Arkhipova, D.M.; Khabibullina, A.V.; Mironova, L.G.; Zakirova, Y.M.; Milyukov, V.A. Biological Activity and Pharmacological Application of Pectic Polysaccharides: A Review. Polymers 2018, 10, 1407. [Google Scholar] [CrossRef] [PubMed]
- Shang, H.; Li, R.; Wu, H.; Sun, Z. Polysaccharides from Trifolium repens L. extracted by different methods and extraction condition optimization. Sci. Rep. 2019, 9, 6353. [Google Scholar] [CrossRef] [PubMed]
- Ayeka, P.A.; Bian, Y.; Githaiga, P.M.; Zhao, Y. The immunomodulatory activities of licorice polysaccharides (Glycyrrhiza uralensis Fisch.) in CT 26 tumor-bearing mice. BMC Complement. Altern. Med. 2017, 17, 536. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-C.; Zhu, Z.-Y.; Liu, Y.-L.; Sun, H.-Q. Comparisons of the anti-tumor activity of polysaccharides from fermented mycelia and cultivated fruiting bodies of Cordyceps militaris in vitro. Int. J. Biol. Macromol. 2019, 130, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Radzki, W.; Sławińska, A.; Skrzypczak, K.; Michalak-Majewska, M. The Impact of Drying of Wild-Growing Mushrooms on the Content and Antioxidant Capacity of Water- Soluble Polysaccharides. Int. J. Med. Mushrooms 2019, 21, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Sun, J.; Liu, J.; Gou, Y.; Zhang, X.; Wu, X.; Sun, R.; Tang, S.; Kan, J.; Qian, C.; et al. Structural characterization and anti-inflammatory activity of alkali-soluble polysaccharides from purple sweet potato. Int. J. Biol. Macromol. 2019, 131, 484–494. [Google Scholar] [CrossRef]
- Xu, J.Y.; Dai, C.; Shan, J.J.; Xie, T.; Xie, H.H.; Wang, M.M.; Yang, G. Determination of the effect of Pinellia ternata (Thunb.) Breit. on nervous system development by proteomics. J. Ethnopharmacol. 2018, 213, 221–229. [Google Scholar] [CrossRef]
- Su, T.; Zhang, W.-W.; Zhang, Y.-M.; Cheng, B.C.-Y.; Fu, X.-Q.; Li, T.; Guo, H.; Li, Y.-X.; Zhu, P.-L.; Cao, H.; et al. Standardization of the manufacturing procedure for Pinelliae Rhizoma Praeparatum cum Zingibere et Alumine. J. Ethnopharmacol. 2016, 193, 663–669. [Google Scholar] [CrossRef]
- Zhang, X.; Cai, Y.; Wang, L.; Liu, H.; Wang, X. Optimization of processing technology of Rhizoma Pinelliae Praeparatum and its anti-tumor effect. Afr. Health Sci. 2015, 15, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-H.; Zhao, Y.-Y.; Cheng, X.-L.; Dai, Z.; Zhou, C.; Bai, X.; Lin, R.-C. General toxicity of Pinellia ternata (Thunb.) Berit. in rat: A metabonomic method for profiling of serum metabolic changes. J. Ethnopharmacol. 2013, 149, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Huang, B.; Wang, G.; Zhang, C. The ethnobotanical, phytochemical and pharmacological profile of the genus Pinellia. Fitoterapia 2014, 93, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Ye, W.H.; Wu, Z.L.; Gao, X.S. The acute, subacute and cumulative toxicity studies on mice of Pinellia ternata and its processed products. Chin. Tradit. Pat. Med. 1988, 10, 18–20. [Google Scholar]
- Liu, Y.J.; Mo, X.L.; Tang, X.Z.; Li, J.H.; Hu, M.B.; Yan, D.; Peng, W.; Wu, C.J. Extraction Optimization, Characterization, and Bioactivities of Polysaccharides from Pinelliae Rhizoma Praeparatum Cum Alumine Employing Ultrasound-Assisted Extraction. Molecules 2017, 22, 965. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-Y.; Huang, X.-X.; Zhang, M.-Y.; Zhou, L.; Li, D.-Q.; Cheng, Z.-Y.; Li, L.-Z.; Peng, Y.; Song, S.-J. Chemical constituents from the tubers of Pinellia ternata (Araceae) and their chemotaxonomic interest. Biochem. Syst. Ecol. 2015, 62, 236–240. [Google Scholar] [CrossRef]
- Liu, F.; Zhu, Z.-Y.; Sun, X.; Gao, H.; Zhang, Y.-M. The preparation of three selenium-containing Cordyceps militaris polysaccharides: Characterization and anti-tumor activities. Int. J. Biol. Macromol. 2017, 99, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.-Y.; Dong, F.; Liu, X.; Lv, Q.; Yang, Y.; Liu, F.; Chen, L.; Wang, T.; Wang, Z.; Zhang, Y. Effects of extraction methods on the yield, chemical structure and anti-tumor activity of polysaccharides from Cordyceps gunnii mycelia. Carbohydr. Polym. 2016, 140, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Nie, C.Z.P.; Zhu, P.L.; Ma, S.P.; Wang, M.C.; Hu, Y.D. Purification, characterization and immunomodulatory activity of polysaccharides from stem lettuce. Carbohydr. Polym. 2018, 188, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.; Li, J.; Chen, Y.; Zhang, L. The structures and biological functions of polysaccharides from traditional Chinese herbs. Prog. Mol. Biol. Transl. Sci. 2019, 163, 423–444. [Google Scholar]
- Iwashita, J.; Yamamoto, T.; Sasaki, Y.; Abe, T. MUC5AC production is downregulated in NCI-H292 lung cancer cells cultured on type-IV collagen. Mol. Cell. Biochem. 2010, 337, 65–75. [Google Scholar] [CrossRef]
- Kaliner, M.; Shelhamer, J.H.; Borson, B.; Nadel, J.; Patow, C.; Marom, Z. Human respiratory mucus. Am. Rev. Respir. Dis. 1986, 134, 612–621. [Google Scholar] [CrossRef]
- Kuperman, D.A.; Huang, X.; Koth, L.L.; Chang, G.H.; Dolganov, G.M.; Zhu, Z.; Elias, J.A.; Sheppard, D.; Erle, D.J. Direct effects of interleukin-13 on epithelial cells cause airway hyperreactivity and mucus overproduction in asthma. Nat. Med. 2002, 8, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, H.; Mukae, H.; Sakamoto, N.; Amenomori, M.; Kitazaki, T.; Imamura, Y.; Fujita, H.; Ishii, H.; Nakayama, S.; Yanagihara, K.; et al. Different effects of telithromycin on MUC5AC production induced by human neutrophil peptide-1 or lipopolysaccharide in NCI-H292 cells compared with azithromycin and clarithromycin. J. Antimicrob. Chemother. 2009, 63, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Han, P.; Yu, L.; Chen, Y.; Ye, B.; Qin, L.; Xin, H.; Han, T. Anti-allergic rhinitis effects of caffeoylquinic acids from the fruits of Xanthium strumarium in rodent animals via alleviating allergic and inflammatory reactions. Rev. Bras. Farm. 2019, 29, 46–53. [Google Scholar] [CrossRef]
- Zhou, E.S.; Fu, Y.H.; Wei, Z.K.; Yu, Y.Q.; Zhang, X.C.; Yang, Z.T. Thymol attenuates allergic airway inflammation in ovalbumin (OVA)-induced mouse asthma. Fitoterapia 2014, 96, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Chen, L.; Xu, S.; Bao, Q.; Ma, Y.; Guo, L.; Zhang, S.; Huang, X.; Cao, C.; Ruan, L. Allergic disorders and risk of depression: A systematic review and meta-analysis of 51 large-scale studies. Ann. Allergy Asthma Immunol. 2018, 120, 310–317. [Google Scholar] [CrossRef]
- Choy, D.F.; Hart, K.M.; Borthwick, L.A.; Shikotra, A.; Nagarkar, D.R.; Siddiqui, S.; Jia, G.; Ohri, C.M.; Doran, E.; Vannella, K.M.; et al. TH2 and TH17 inflammatory pathways are reciprocally regulated in asthma. Sci. Transl. Med. 2015, 7, 301ra129. [Google Scholar] [CrossRef]
- Troy, N.M.; Hollams, E.M.; Holt, P.G.; Bosco, A. Differential gene network analysis for the identification of asthma-associated therapeutic targets in allergen-specific T-helper memory responses. BMC Med. Genom. 2016, 9, 9. [Google Scholar] [CrossRef]
- Abdelmotelb, A.M.; Rose-Zerilli, M.J.; Barton, S.J.; Holgate, S.T.; Walls, A.F.; Holloway, J.W. Alpha-tryptase gene variation is associated with levels of circulating IgE and lung function in asthma. Clin. Exp. Allergy 2014, 44, 822–830. [Google Scholar] [CrossRef]
- Guo, H.-W.; Yun, C.-X.; Hou, G.-H.; Du, J.; Huang, X.; Lu, Y.; Keller, E.T.; Zhang, J.; Deng, J.-G. Mangiferin Attenuates Th1/Th2 Cytokine Imbalance in an Ovalbumin-Induced Asthmatic Mouse Model. PLoS ONE 2014, 9, e100394. [Google Scholar] [CrossRef]
- Yu, Y.; Shen, M.; Wang, Z.; Wang, Y.; Xie, M.; Xie, J.-H. Sulfated polysaccharide from Cyclocarya paliurus enhances the immunomodulatory activity of macrophages. Carbohydr. Polym. 2017, 174, 669–676. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, Y.; Lv, J.; Cheng, M.; Wu, Y.; Cao, K.; Zhang, X.; Mou, X.; Fan, Q. Structure characterization of two functional polysaccharides from Polygonum multiflorum and its immunomodulatory. Int. J. Biol. Macromol. 2018, 113, 195–204. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, X.; Wang, F.; Xu, J.; Tang, X.; Li, N. Structural characterization and antioxidant activity of polysaccharide from ginger. Int. J. Biol. Macromol. 2018, 111, 862–869. [Google Scholar] [CrossRef]
- Fan, W.; Zhang, S.; Hao, P.; Zheng, P.; Liu, J.; Zhao, X. Structure characterization of three polysaccharides and a comparative study of their immunomodulatory activities on chicken macrophage. Carbohydr. Polym. 2016, 153, 631–640. [Google Scholar] [CrossRef]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.-I.; Lee, Y.C. Carbohydrate analysis by a phenol–sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef]
- Kolsi, R.B.A.; Fakhfakh, J.; Krichen, F.; Jribi, I.; Chiarore, A.; Patti, F.P.; Blecker, C.; Allouche, N.; Belghith, H.; Belghith, K. Structural characterization and functional properties of antihypertensive Cymodocea nodosa sulfated polysaccharide. Carbohydr. Polym. 2016, 151, 511–522. [Google Scholar] [CrossRef]
- Zhang, T.-T.; Lu, C.-L.; Jiang, J.-G.; Wang, M.; Wang, D.-M.; Zhu, W. Bioactivities and extraction optimization of crude polysaccharides from the fruits and leaves of Rubus chingii Hu. Carbohydr. Polym. 2015, 130, 307–315. [Google Scholar] [CrossRef]
- Ben Romdhane, M.; Haddar, A.; Ghazala, I.; Ben Jeddou, K.; Helbert, C.B.; Ellouz-Chaabouni, S. Optimization of polysaccharides extraction from watermelon rinds: Structure, functional and biological activities. Food Chem. 2017, 216, 355–364. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Xu, L.; Wu, Q.; Wang, Q.; Kong, W.; Liang, J.; Yao, J.; Zhang, J. Synthesis and structural features of phosphorylated Artemisia sphaerocephala polysaccharide. Carbohydr. Polym. 2018, 181, 19–26. [Google Scholar] [CrossRef]
- He, P.; Zhang, A.; Zhang, F.; Linhardt, R.J.; Sun, P. Structure and bioactivity of a polysaccharide containing uronic acid from Polyporus umbellatus sclerotia. Carbohydr. Polym. 2016, 152, 222–230. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, W.; Yu, J.; Zou, S.; Wang, J.; Yao, W.; Gao, X. Characterization and hypoglycemic effect of a polysaccharide extracted from the fruit of Lycium barbarum L. Carbohydr. Polym. 2013, 98, 8–16. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.; Chen, J.; Wang, S. A polysaccharide from Pinellia ternata inhibits cell proliferation and metastasis in human cholangiocarcinoma cells by targeting of Cdc42 and 67 kDa Laminin Receptor (LR). Int. J. Biol. Macromol. 2016, 93, 520–525. [Google Scholar] [CrossRef]
- Li, Q.; Yang, S.; Yang, S.; Xin, F.; Wang, M. Anti-inflammatory activity of phlomisoside F isolated from Phlomis younghusbandii Mukerjee. Int. Immunopharmacol. 2015, 28, 724–730. [Google Scholar] [CrossRef]
- Iwashita, J.; Ito, Y.; Yokoo, M.; Takahashi, S.; Murata, J. Akt induces down regulation of MUC5AC production in NCI-H292 human airway epithelial cells cultured on extracellular matrix. Biosci. Biotechnol. Biochem. 2014, 78, 212–221. [Google Scholar] [CrossRef] [Green Version]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, M.; Liu, Y.; Wang, L.; Wang, J.; Li, L.; Wu, C. Purification, Characterization of Two Polysaccharides from Pinelliae Rhizoma Praeparatum Cum Alumine and Their Anti-Inflammatory Effects on Mucus Secretion of Airway Epithelium. Int. J. Mol. Sci. 2019, 20, 3553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143553
Hu M, Liu Y, Wang L, Wang J, Li L, Wu C. Purification, Characterization of Two Polysaccharides from Pinelliae Rhizoma Praeparatum Cum Alumine and Their Anti-Inflammatory Effects on Mucus Secretion of Airway Epithelium. International Journal of Molecular Sciences. 2019; 20(14):3553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143553
Chicago/Turabian StyleHu, Meibian, Yujie Liu, Li Wang, Jiaolong Wang, Lin Li, and Chunjie Wu. 2019. "Purification, Characterization of Two Polysaccharides from Pinelliae Rhizoma Praeparatum Cum Alumine and Their Anti-Inflammatory Effects on Mucus Secretion of Airway Epithelium" International Journal of Molecular Sciences 20, no. 14: 3553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143553