Genome-Wide Analysis of Carboxylesterases (COEs) in the Whitefly, Bemisia tabaci (Gennadius)

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results
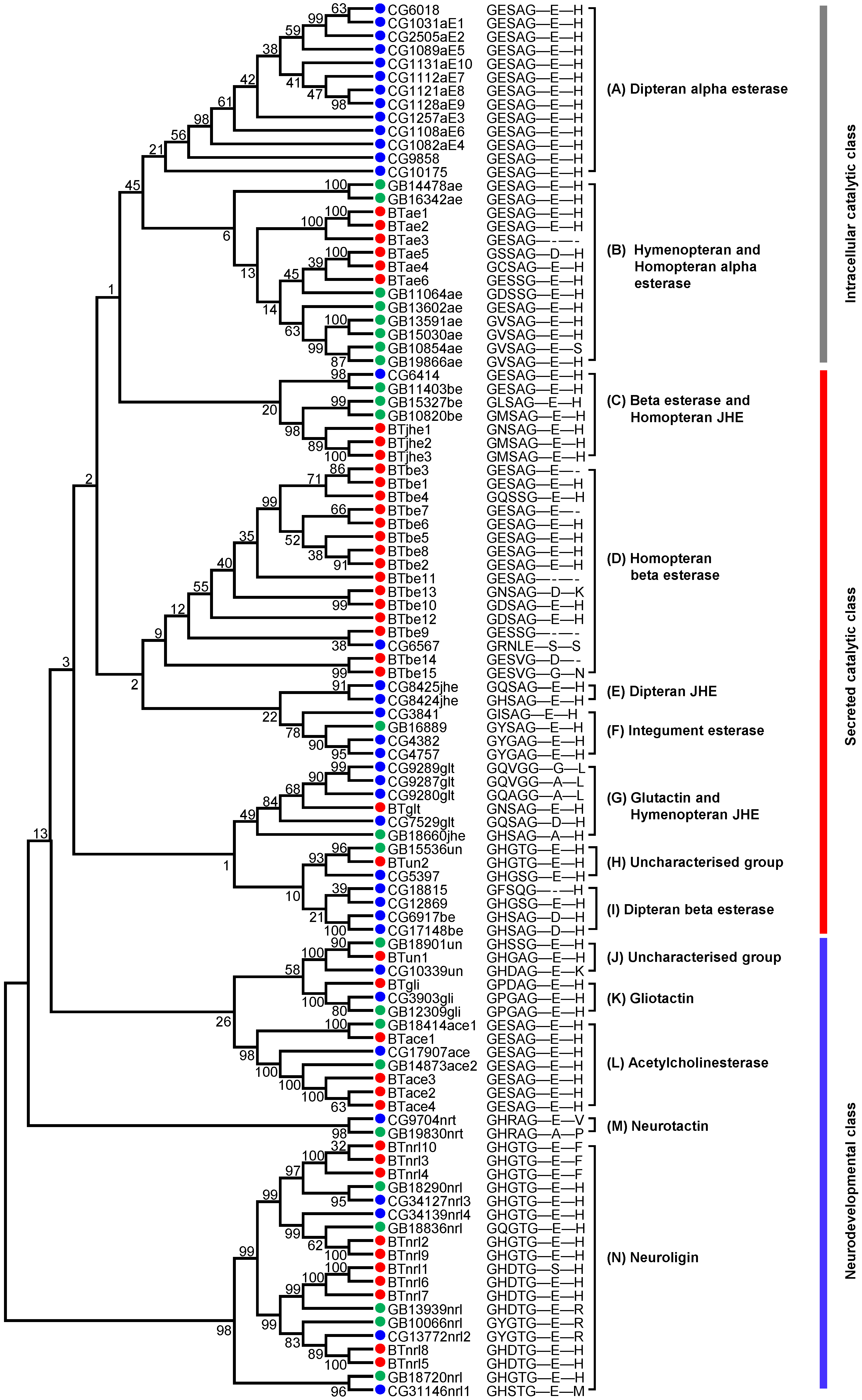
2.1. Identification of COEs in the Genome of B. tabaci MED
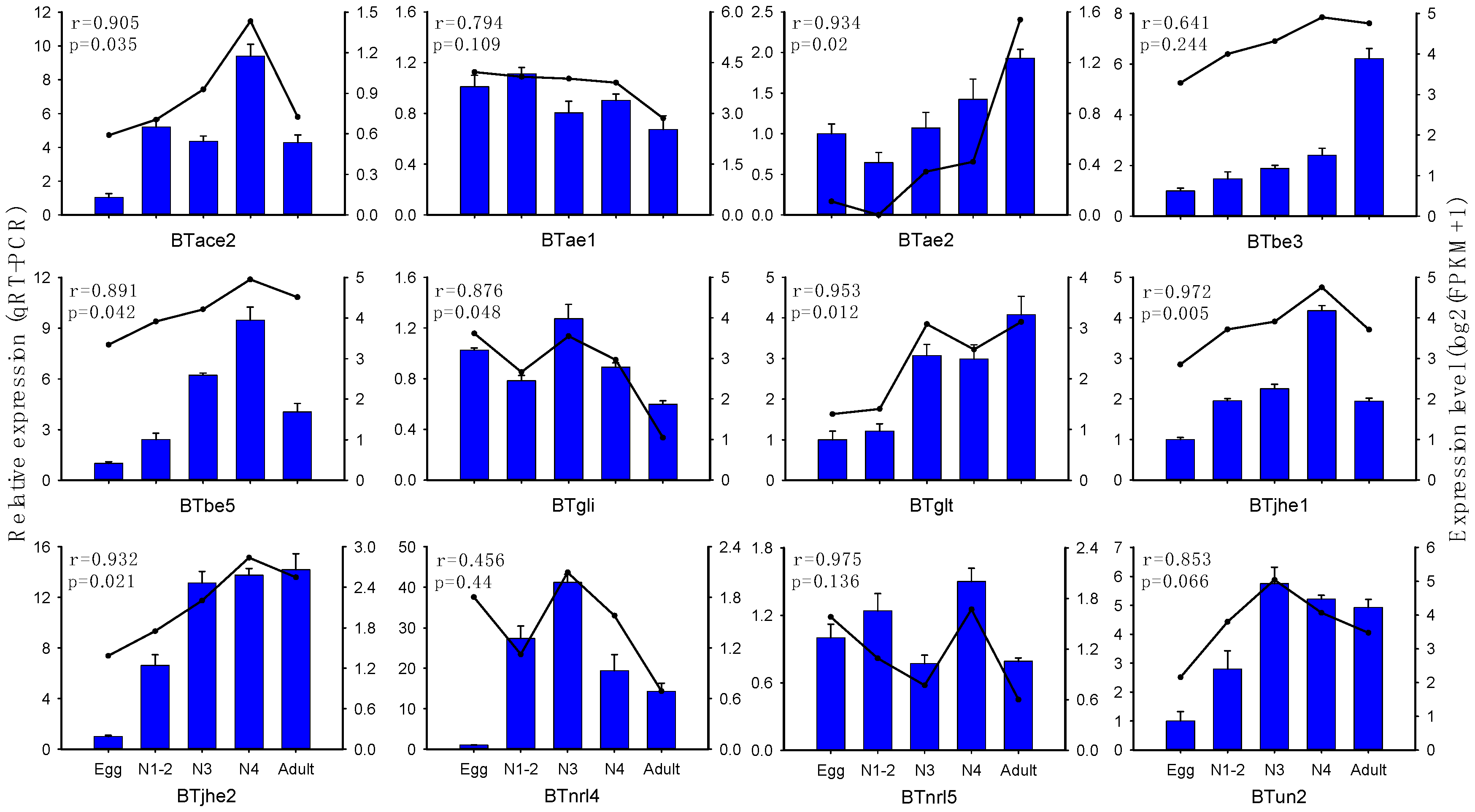
2.2. Expression Profiling of B. tabaci MED COE Genes
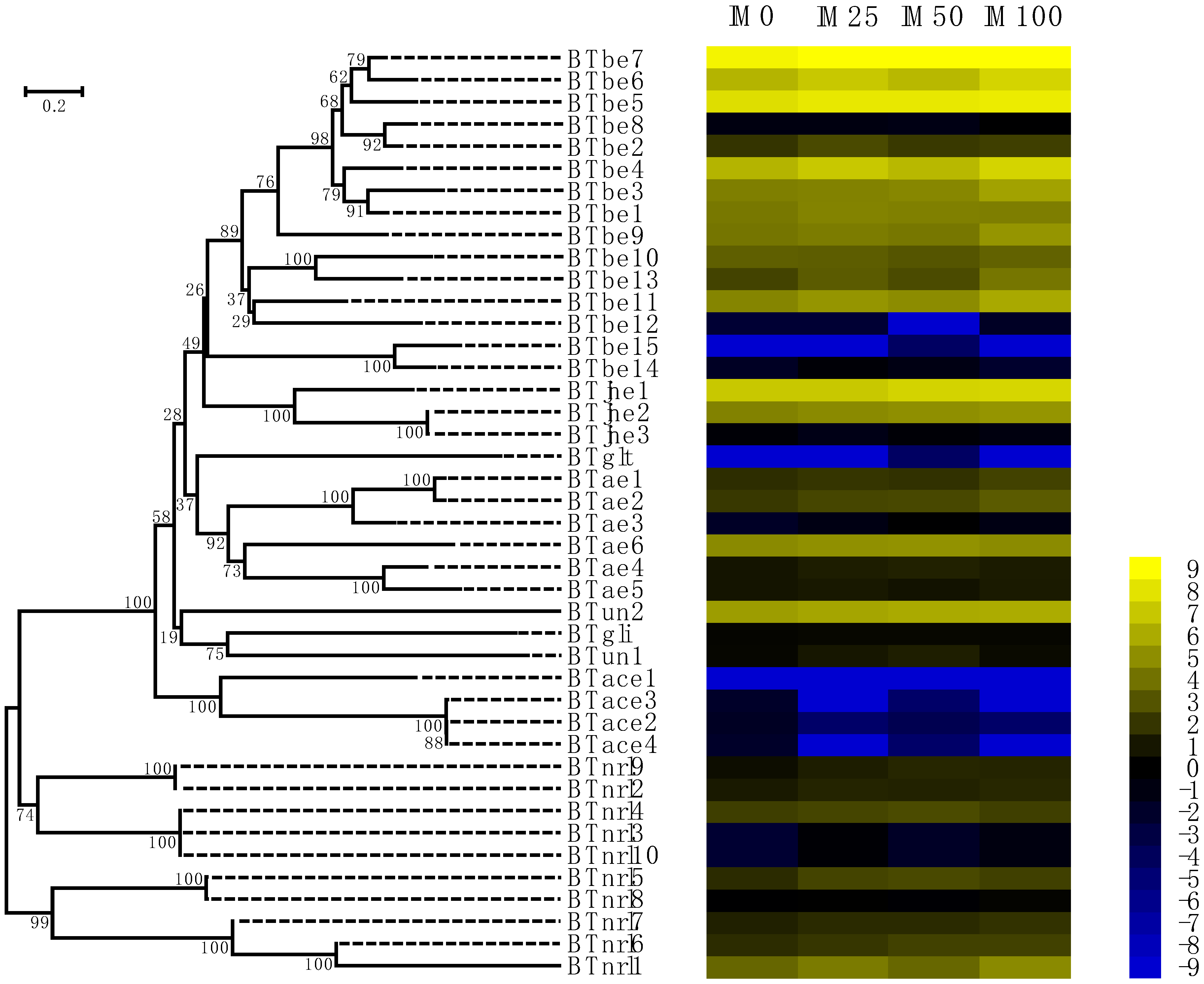
2.3. Responses of COE Expression to Imidacloprid
2.4. COEs Involved in the Imidacloprid Resistance of B. tabaci MED
3. Discussion
4. Materials and Methods
4.1. Insect Strain
4.2. De Novo Identification of COE Genes
4.3. Phylogenetic Analysis of B. tabaci MED COE Genes
4.4. RNA-Seq Analysis
4.5. RNA Isolation and cDNA Synthesis
4.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
4.7. dsRNA Synthesis and RNAi Assays
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| COE | Carboxylesterase |
| RNAi | RNA interference |
| FPKM | Fragments per kilobase of transcript per million fragments mapped |
| NJ | Neighbor-joining |
| JTT | Jones–Taylor–Thornton |
| dsRNA | Double-strand RNA |
| EGFP | Enhanced green fluorescent protein |
| JH | Juvenile hormone |
| JHE | Juvenile hormone esterase |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| AAAP | Amino acid/auxin permease |
| P450 | Cytochrome P450 monooxygenase |
| GST | Glutathione S-transferase |
| ABC | ATP-binding cassette transporter |
| mtCOI | Mitochondrial cytochrome oxidase I |
| HMM | Hidden Markov model |
| NCBI | National Center for Biotechnology Information |
| EST | Expressed sequence tag |
References
- Wheelock, C.E.; Shan, G.; Ottea, J. Overview of carboxylesterases and their role in the metabolism of insecticides. J. Pestic Sci. 2005, 30, 75–83. [Google Scholar] [CrossRef]
- Satoh, T.; Hosokawa, M. The mammalian carboxylesterases: From molecules to functions. Annu. Rev. Pharm. Toxicol. 1998, 38, 257–288. [Google Scholar] [CrossRef] [PubMed]
- Montella, I.R.; Schama, R.; Valle, D. The classification of esterases: An important gene family involved in insecticide resistance-a review. Mem. Inst. Oswaldo Cruz. 2012, 107, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.D.G.; Putterill, J.J.; Plummer, K.M.; Newcomb, R.D. The carboxylesterase gene family from Arabidopsis thaliana. J. Mol. Evol. 2003, 57, 487–500. [Google Scholar]
- Bornscheuer, U.T. Microbial carboxylesterases: Classification, properties and application in biocatalysis. Fems. Microbiol. Rev. 2002, 26, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Oakeshott, J.G.; Claudianos, C.; Campbell, P.M.; Newcomb, R.D.; Russell, R.J. Biochemical genetics and genomics of insect esterases. In Insect Pharmacology: Channels, Receptors, Toxins and Enzymes, 1st ed.; Elsevier: London, UK, 2005; pp. 309–381. [Google Scholar]
- Feng, X.; Li, M.; Liu, N. Carboxylesterase genes in pyrethroid resistant house flies, Musca Domest. Insect Biochem. Molec. Biol. 2018, 92, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Alon, M.; Alon, F.; Nauen, R.; Morin, S. Organophosphates’ resistance in the B-biotype of Bemisia tabaci (Hemiptera: Aleyrodidae) is associated with a point mutation in an ace1-type acetylcholinesterase and overexpression of carboxylesterase. Insect Biochem. Molec. Biol. 2008, 38, 940–949. [Google Scholar] [CrossRef]
- Andrews, M.C.; Callaghan, A.; Field, L.M.; Williamson, M.S.; Moores, G.D. Identification of mutations conferring insecticide-insensitive AChE in the cotton-melon aphid, Aphis gossypii Glover. Insect Mol. Biol. 2004, 13, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Dowdy, A.K.; Baker, J.E. Detection of single-base substitution in an esterase gene and its linkage to malathion resistance in the parasitoid Anisopteromalus calandrae (Hymenoptera: Pteromalidae). Pestic. Sci. 1999, 55, 398–404. [Google Scholar] [CrossRef]
- Campbell, P.M.; Newcomb, R.D.; Russell, R.J.; Oakeshott, J.G. Two different amino acid substitutions in the ali-esterase, E3, confer alternative types of organophosphorus insecticide resistance in the sheep blowfly, Lucilia cuprina. Insect Biochem. Molec. Biol. 1998, 28, 139–150. [Google Scholar] [CrossRef]
- Claudianos, C.; Russell, R.J.; Oakeshott, J.G. The same amino acid substitution in orthologous esterases confers organophosphate resistance on the house fly and a blowfly. Insect Biochem. Molec. Biol. 1999, 29, 675–686. [Google Scholar] [CrossRef]
- Small, G.J.; Hemingway, J. Molecular characterization of the amplified carboxylesterase gene associated with organophosphorus insecticide resistance in the brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2000, 9, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.W.; Zhang, J.; Gao, X.W.; Liang, P.; Guo, H.L. Overexpression of carboxylesterase gene associated with organophosphorous insecticide resistance in cotton aphids, Aphis gossypii (Glover). Pestic Biochem. Physiol. 2008, 90, 175–180. [Google Scholar] [CrossRef]
- Wu, S.; Yang, Y.; Yuan, G.; Campbell, P.M.; Teese, M.G.; Russell, R.J.; Oakeshott, J.G.; Wu, Y. Overexpressed esterases in a fenvalerate resistant strain of the cotton bollworm, Helicoverpa armigera. Insect Biochem. Molec. Biol. 2011, 41, 14–21. [Google Scholar] [CrossRef]
- Hawkes, N.J.; Hemingway, J. Analysis of the promoters for the β-esterase genes associated with insecticide resistance in the mosquito Culex quinquefasciatus. Biochim. Biophys. Acta 2002, 1574, 51–62. [Google Scholar] [CrossRef]
- Gong, Y.; Shi, X.; Desneux, N.; Gao, X. Effects of spirotetramat treatments on fecundity and carboxylesterase expression of Aphis gossypii Glover. Ecotoxicology 2016, 25, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Lü, F.G.; Fu, K.Y.; Li, Q.; Guo, W.C.; Ahmat, T.; Li, G.Q. Identification of carboxylesterase genes and their expression profiles in the Colorado potato beetle Leptinotarsa decemlineata treated with fipronil and cyhalothrin. Pestic Biochem. Physiol. 2015, 122, 86–95. [Google Scholar] [CrossRef]
- Poupardin, R.; Reynaud, S.; Strode, C.; Ranson, H.; Vontas, J.; David, J.P. Crossinduction of detoxification genes by environmental xenobiotics and insecticides in the mosquito Aedes aegypti: Impact on larval tolerance to chemical insecticides. Insect Biochem. Molec. Biol. 2008, 38, 540–551. [Google Scholar] [CrossRef]
- Wei, P.; Shi, L.; Shen, G.; Xu, Z.; Liu, J.; Pan, Y.; He, L. Characteristics of carboxylesterase genes and their expression-level between acaricide-susceptible and resistant Tetranychus cinnabarinus (Boisduval). Pestic Biochem. Physiol. 2016, 131, 87–95. [Google Scholar] [CrossRef]
- Reitz, S.R. Invasion of the whiteflies. Science 2007, 318, 1733–1734. [Google Scholar] [CrossRef]
- Gelman, D.B.; Blackburn, M.B.; Hu, J.S. Identification of the molting hormone of the sweet potato (Bemisia tabaci) and greenhouse (Trialeurodes vaporariorum) whitefly. J. Insect Physiol. 2005, 51, 47–53. [Google Scholar] [CrossRef] [PubMed]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Dinsdale, A.; Cook, L.; Riginos, C.; Buckley, Y.M.; De Barro, P. Refined global analysis of Bemisia tabaci (Hemiptera: Sternorrhyncha: Aleyrodoidea: Aleyrodidae) mitochondrial cytochrome oxidase I to identify species level genetic boundaries. Ann. Entomol. Soc. Am. 2010, 103, 196–208. [Google Scholar] [CrossRef]
- Chu, D.; Wan, F.H.; Zhang, Y.J.; Brown, J.K. Change in the biotype composition of Bemisia tabaci in Shandong province of China from 2005 to 2008. Env. Entomol. 2010, 39, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Preisser, E.L.; Chu, D.; Wang, S.; Wu, Q.; Carrière, Y.; Zhou, X.; Zhang, Y. Insecticides promote viral outbreaks by altering herbivore competition. Ecol. Appl. 2015, 25, 1585–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Xie, W.; Wang, S.; Wu, Q.; Zhou, X.; Zhang, Y. Dynamic monitoring (B versus Q) and further resistance status of Q-type Bemisia tabaci in China. Crop Prot. 2017, 94, 115–122. [Google Scholar] [CrossRef]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef]
- Nauen, R.; Jeschke, P.; Copping, L. In focus: Neonicotinoid insecticides editorial. Pest Manag. Sci. 2008, 64, 1081. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Cahill, M.; Gorman, K.; Day, S.; Denholm, I.; Elbert, A.; Nauen, R. Baseline determination and detection of resistance to imidacloprid in Bemisia tabaci (Homoptera: Aleyrodidae). Bull. Entomol. Res. 1996, 86, 343–349. [Google Scholar] [CrossRef]
- Palumbo, J.C.; Horowitz, A.R.; Prabhaker, N. Insecticidal control and resistance management for Bemisia tabaci. Crop Prot. 2001, 20, 739–765. [Google Scholar] [CrossRef]
- Roditakis, E.; Morou, E.; Tsagkarakou, A.; Riga, M.; Nauen, R.; Paine, M.; Morin, S.; Vontas, J. Assessment of the Bemisia tabaci CYP6CM1vQ transcript and protein levels in laboratory and field-derived imidacloprid-resistant insects and cross-metabolism potential of the recombinant enzyme. Insect Sci. 2011, 18, 23–29. [Google Scholar] [CrossRef]
- Byrne, F.J.; Castle, S.; Prabhaker, N.; Toscano, N.C. Biochemical study of resistance to imidacloprid in biotype B Bemisia tabaci from Guatemala. Pest Manag. Sci. 2003, 59, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yan, H.; Yang, Y.; Wu, Y. Biotype and insecticide resistance status of the whitefly Bemisia tabaci from China. Pest Manag. Sci. 2010, 66, 1360–1366. [Google Scholar] [CrossRef]
- Kontsedalov, S.; Abu-moch, F.; Lebedev, G.; Czosnek, H.; Horowitz, A.R.; Ghanim, M. Bemisia tabaci biotype dynamics and resistance to insecticides in Israel during the years 2008–2010. J. Integr. Agric. 2012, 11, 312–320. [Google Scholar] [CrossRef]
- Karunker, I.; Benting, J.; Lueke, B.; Ponge, T.; Nauen, R.; Roditakis, E.; Vontas, J.; Gorman, K.; Denholm, I.; Morin, S. Over-expression of cytochrome P450 CYP6CM1 is associated with high resistance to imidacloprid in the B and Q biotypes of Bemisia tabaci (Hemiptera: Aleyrodidae). Insect Biochem. Molec. Biol. 2008, 38, 634–644. [Google Scholar] [CrossRef]
- Karunker, I.; Morou, E.; Nikou, D.; Nauen, R.; Sertchook, R.; Stevenson, B.J.; Paine, M.J.I.; Morin, S.; Vontas, J. Structural model and functional characterization of the Bemisia tabaci CYP6CM1vQ, a cytochrome P450 associated with high levels of imidacloprid resistance. Insect Biochem. Molec. Biol. 2009, 39, 697–706. [Google Scholar] [CrossRef]
- Yang, X.; He, C.; Xie, W.; Liu, Y.; Xia, J.; Yang, Z.; Guo, L.; Wen, Y.; Wang, S.; Wu, Q.; et al. Glutathione S-transferases are involved in thiamethoxam resistance in the field whitefly Bemisia tabaci Q (Hemiptera: Aleyrodidae). Pestic. Biochem. Physiol. 2016, 134, 73–78. [Google Scholar] [CrossRef]
- He, C.; Xie, W.; Yang, X.; Wang, S.L.; Wu, Q.J.; Zhang, Y.J. Identification of glutathione S-transferases in Bemisia tabaci (Hemiptera: Aleyrodidae) and evidence that GSTd7 helps explain the difference in insecticide susceptibility between B. tabaci Middle East-Minor Asia 1 and Mediterranean. Insect Mol. Biol. 2018, 27, 22–35. [Google Scholar] [CrossRef]
- Hamada, A.; Wahl, G.D.; Nesterov, A.; Nakao, T.; Kawashima, M.; Banba, S. Differential metabolism of imidacloprid and dinotefuran by Bemisia tabaci CYP6CM1 variants. Pestic. Biochem. Physiol. 2019, 159, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Chen, C.; Yang, Z.; Guo, L.; Yang, X.; Wang, D.; Chen, M.; Huang, J.; Wen, Y.; Zeng, Y.; et al. Genome sequencing of the sweetpotato whitefly Bemisia tabaci MED/Q. GigaScience 2017, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Song, T.; He, R.; Zeng, Y.; Xie, W.; Wu, Q.; Wang, S.; Zhou, X.; Zhang, Y. Genome-wide analysis of ATP-binding cassette (ABC) transporters in the sweetpotato whitefly, Bemisia tabaci. BMC Genom. 2017, 18, 330. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.Y.; Lu, C.; Li, W.L.; Xiang, Z.H.; Zhang, Z. Annotation and expression of carboxylesterases in the silkworm, Bombyx mori. BMC Genom. 2009, 10, 553. [Google Scholar] [CrossRef] [PubMed]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A deficit of detoxification enzymes: Pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef]
- Guerrero, F.D. Cloning of a horn fly cDNA, HiαE7, encoding an esterase whose transcript concentration is elevated in diazinon-resistant flies. Insect Biochem. Molec. Biol. 2000, 30, 1107–1115. [Google Scholar] [CrossRef]
- Robin, C.; Bardsley, L.M.J.; Coppin, C.; Oakeshott, J.G. Birth and death of genes and functions in the β-esterase cluster of Drosophila. J. Mol. Evol. 2009, 69, 10–21. [Google Scholar] [CrossRef]
- Scheiffele, P.; Fan, J.; Choih, J.; Fetter, R.; Serafini, T. Neuroligin expressed in nonneuronal cells triggers presynaptic development in contacting axons. Cell 2000, 101, 657–669. [Google Scholar] [CrossRef]
- Chen, X.; Liu, H.; Shim, A.H.R.; Focia, P.J.; He, X. Structural basis for synaptic adhesion mediated by neuroligin-neurexin interactions. Nat. Struct. Mol. Biol. 2008, 15, 50–56. [Google Scholar] [CrossRef]
- Tan, A.; Tanaka, H.; Tamura, T.; Shiotsuki, T. Precocious metamorphosis in transgenic silkworms overexpressing juvenile hormone esterase. Proc. Natl. Acad. Sci. USA 2005, 102, 11751–11756. [Google Scholar] [CrossRef] [Green Version]
- Riddiford, L.M.; Hiruma, K.; Zhou, X.; Nelson, C.A. Insights into the molecular basis of the hormonal control of molting and metamorphosis from Manduca sexta and Drosophila melanogaster. Insect Biochem. Molec. Biol. 2003, 33, 1327–1338. [Google Scholar] [CrossRef]
- Gu, X.; Kumar, S.; Kim, E.; Kim, Y. A whole genome screening and RNA interference identify a juvenile hormone esterase-like gene of the diamondback moth, Plutella Xylostella. J. Insect Physiol. 2015, 80, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Li, T.; Zhang, L.; Liu, N. Co-up-regulation of three P450 genes in response to permethrin exposure in permethrin resistant house flies, Musca domestica. Bmc. Physiol. 2008, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Terriere, L.C. Induction of detoxication enzymes in insects. Ann. Rev. Entomol. 1984, 29, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.Y.; Xue, X.Y.; Huang, Y.P.; Chen, X.Y.; Mao, Y.B. Gossypol-enhanced P450 gene pool contributes to cotton bollworm tolerance to a pyrethroid insecticide. Mol. Ecol. 2012, 21, 4371–4385. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, W.; Wang, S.; Wu, Q.; Pan, H.; Li, R.; Yang, N.; Liu, B.; Xu, B.; Zhou, X.; et al. Two cytochrome P450 genes are involved in imidacloprid resistance in field populations of the whitefly, Bemisia tabaci, in China. Pestic. Biochem. Physiol. 2013, 107, 343–350. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Liang, J.; Liu, S.; Wang, S.; Wu, Q.; Xie, W.; Zhang, Y. Changes in the expression of four ABC transporter genes in response to imidacloprid in Bemisia tabaci Q (Hemiptera: Aleyrodidae). Pestic. Biochem. Physiol. 2019, 153, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Molec. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Ocampo, C.B.; Salazar-Terreros, M.J.; Mina, N.J.; McAllister, J.; Brogdon, W. Insecticide resistance status of Aedes aegypti in 10 localities in Colombia. Acta. Trop. 2011, 118, 37–44. [Google Scholar] [CrossRef]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega 6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. Tophat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xie, W.; Wang, S.; Wu, Q.; Yang, N.; Yang, X.; Pan, H.; Zhou, X.; Bai, L.; Xu, B.; et al. Reference gene selection for qRT-PCR analysis in the sweetpotato whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). PLoS ONE 2013, 8, e53006. [Google Scholar] [CrossRef]
- Xia, J.; Yang, Z.; Gong, C.; Xie, W.; Pan, H.; Guo, Z.; Zheng, H.; Yang, X.; Sun, X.; Kang, S.; et al. Genome-wide identification and expression analysis of amino acid transporters in the whitefly, Bemisia tabaci (Gennadius). Int. J. Biol. Sci. 2017, 13, 735–747. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class/Clades | B. mori | D. melanogaster | Ap. mellifera | An. gambiae | T. castaneum | A. pisum | B. tabaci |
|---|---|---|---|---|---|---|---|
| Intracellular catalytic class | |||||||
| α-esterase | 55 | 13 | 8 | 16 | 26 | 5 | 6 |
| secreted catalytic class | |||||||
| JHE | 4 | 2 | 1 | 9 | 1 | 0 | 3 |
| integument esterase | 2 | 3 | 1 | 0 | 2 | 0 | 0 |
| β-esterase | 2 | 6 | 3 | 5 | 8 | 15 | 15 |
| uncharacterized | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| glutactin | 0 | 4 | 0 | 9 | 0 | 0 | 1 |
| neurodevelopmental class | |||||||
| AChE | 2 | 1 | 2 | 2 | 2 | 2 | 4 |
| uncharacterized | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| gliotactin | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| neuroligin | 6 | 4 | 5 | 5 | 4 | 3 | 10 |
| neurotactin | 2 | 1 | 1 | 2 | 2 | 0 | 0 |
| total | 76 | 37 | 24 | 51 | 48 | 28 | 42 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, J.; Xu, H.; Yang, Z.; Pan, H.; Yang, X.; Guo, Z.; Yang, F.; Guo, L.; Sun, X.; Wang, S.; et al. Genome-Wide Analysis of Carboxylesterases (COEs) in the Whitefly, Bemisia tabaci (Gennadius). Int. J. Mol. Sci. 2019, 20, 4973. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20204973
Xia J, Xu H, Yang Z, Pan H, Yang X, Guo Z, Yang F, Guo L, Sun X, Wang S, et al. Genome-Wide Analysis of Carboxylesterases (COEs) in the Whitefly, Bemisia tabaci (Gennadius). International Journal of Molecular Sciences. 2019; 20(20):4973. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20204973
Chicago/Turabian StyleXia, Jixing, Haifeng Xu, Zezhong Yang, Huipeng Pan, Xin Yang, Zhaojiang Guo, Fengshan Yang, Litao Guo, Xiaodong Sun, Shaoli Wang, and et al. 2019. "Genome-Wide Analysis of Carboxylesterases (COEs) in the Whitefly, Bemisia tabaci (Gennadius)" International Journal of Molecular Sciences 20, no. 20: 4973. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20204973