Presence of 15p Marker D15Z1 on the Short Arm of Acrocentric Chromosomes is Associated with Aneuploid Offspring in Mexican Couples

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Population Cohorts.
2.2. Frequency of Extra D15Z1 Region in the Different Groups of Study
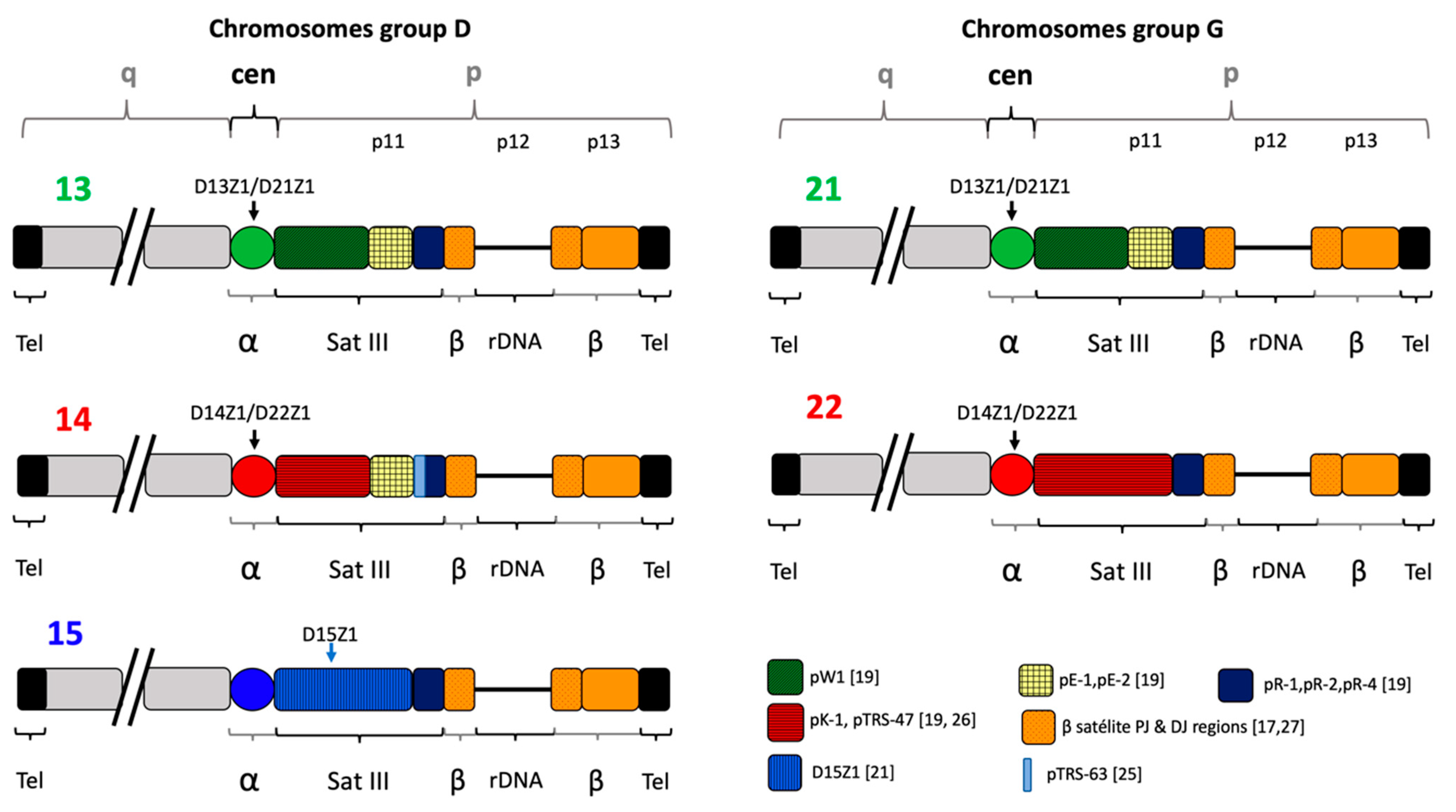
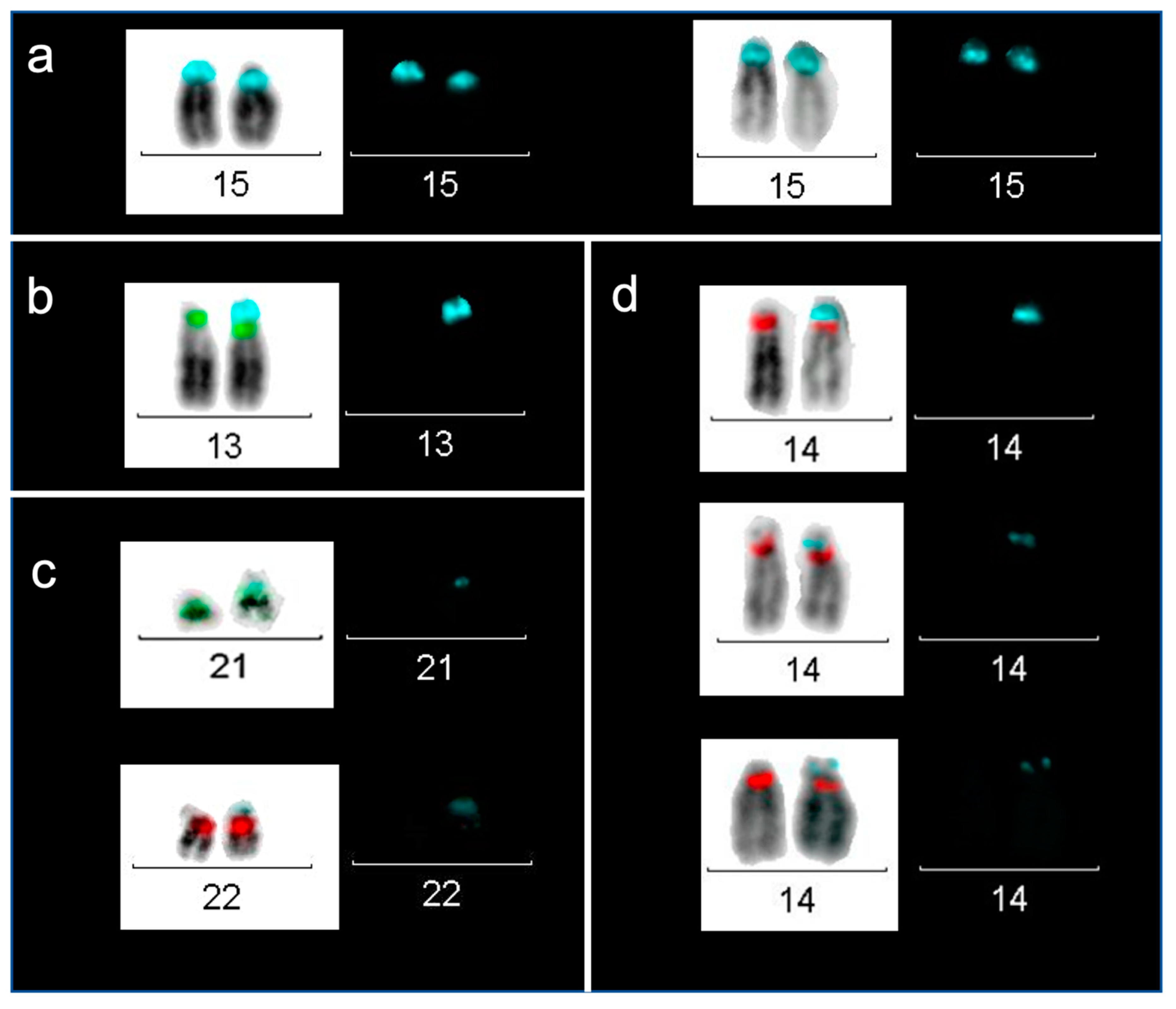
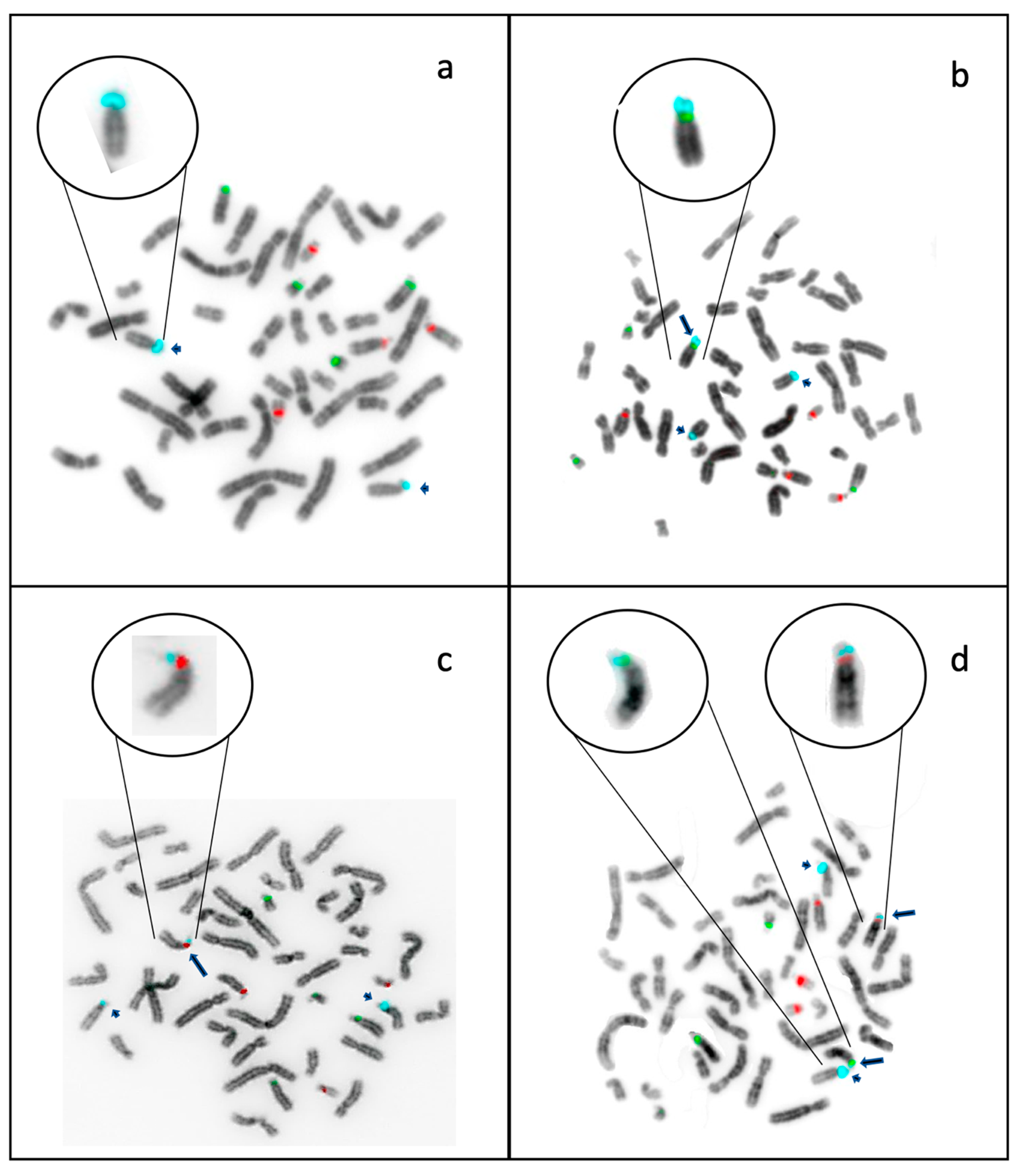
2.3. Localization of D15Z1 on Acrocentric Chromosomes other than 15.
3. Discussion
- (a)
- in this study, only a minority of the additional D15Z1 signals was found on chromosome 21p, the chromosome that is involved in aneuploidy of the majority of the offspring.
- (b)
- we detected similar frequencies for the D15Z1 polymorphism between both male and female progenitors, which rules out an exclusive association between the presence of this polymorphism and non-disjunction events during oogenesis. Non-disjunction events during oogenesis are widely known to be responsible for >90% of events leading to trisomy 21, as well as for other acrocentric chromosome trisomies [4].
- (c)
- the frequency of additional D15Z1 signals in the aneuploid offspring group was 21.9%, which is similar to a previously reported cohort of aneuploid abortions that presented the variant in 17.7% of the products, without a significant difference with the healthy population [40], although in aneuploid descendants with the polymorphism, either born alive or aborted, the acceptor chromosome was mostly 13pvar(D15Z1).
4. Conclusions
5. Materials and Methods
5.1. Patient Cohorts
5.2. Cytogenetic Study with GTG bands
5.3. Fluorescence In Situ Hybridization
5.4. FISH Analyses
5.5. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DA/DAPI | Distamycin A/4,6-diamino-2-phenyl-indole |
| DAPI | 4’,6-diamidino-2-phenylindole |
| FISH | Fluorescence In Situ Hybridization |
| GTG | Giemsa-Trypsin G banding |
| ISCN | International System for Human Cytogenomic Nomenclature |
| NOR | Nucleolar organization region |
| rDNA | Ribosomal DNA, Clusters of DNA sequences containing repeated units of rRNA genes |
| rRNA | Ribosomal ribonucleic acid |
References
- Page, S.L.; Shin, J.C.; Han, J.Y.; Choo, K.H.A.; Shaffer, L.G. Breakpoint diversity illustrates distinct mechanisms for Robertsonian translocation formation. Hum. Mol. Genet. 1996, 5, 1278–1279. [Google Scholar] [CrossRef] [PubMed]
- Bint, S.M.; Scriven, P.N.; Ogilvie, C.M. Successful PGD cycles for mosaic Robertsonian translocation carriers provide insights into the mechanism of formation of the derivative chromosomes. Am. J. Med. Genet. Part A 2013, 161, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Ohno, M.; Matsunobu, A.; Yoshihara, K.; Yabe, N. A cytogenetic survey of 14,835 consecutive liveborns. Jpn. J. Hum. Genet. 1991, 36, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Hassold, T.; Hall, H.; Hunt, P. The origin of human aneuploidy: Where we have been, where we are going. Hum. Mol. Genet. 2007, 16, 203–208. [Google Scholar] [CrossRef]
- Marchetti, F.; Massarotti, A.; Yauk, C.L.; Pacchierotti, F.; Russo, A. The adverse outcome pathway (AOP) for chemical binding to tubulin in oocytes leading to aneuploid offspring. Environ. Mol. Mutagen. 2016, 57, 87–113. [Google Scholar] [CrossRef]
- Jacobs, P.A.; Melville, M.; Ratcliffe, S.; Keay, A.J.; Syme, J. A cytogenetic survey of 11,680 newborn infants. Ann. Hum. Genet. 1974, 37, 359–376. [Google Scholar] [CrossRef]
- Franasiak, J.M.; Forman, E.J.; Hong, K.H.; Werner, M.D.; Upham, K.M.; Treff, N.R.; Scott, R.T. Aneuploidy across individual chromosomes at the embryonic level in trophectoderm biopsies: Changes with patient age and chromosome structure. J. Assist. Reprod. Genet. 2014, 31, 1501–1509. [Google Scholar] [CrossRef]
- Jia, C.W.; Wang, L.; Lan, Y.L.; Song, R.; Zhou, L.Y.; Yu, L.; Yang, Y.; Liang, Y.; Li, Y.; Ma, Y.M.; et al. Aneuploidy in early miscarriage and its related factors. Chin. Med. J. (Engl). 2015, 128, 2772–2776. [Google Scholar] [CrossRef]
- Crolla, J.A.; Youings, S.A.; Ennis, S.; Jacobs, P.A. Supernumerary marker chromosomes in man: Parental origin, mosaicism and maternal age revisited. Eur. J. Hum. Genet. 2005, 13, 154–160. [Google Scholar] [CrossRef]
- Liehr, T.; Mrasek, K.; Kosyakova, N.; Ogilvie, C.; Vermeesch, J.; Trifonov, V.; Rubtsov, N. Small supernumerary marker chromosomes (sSMC) in humans; are there B chromosomes hidden among them. Mol. Cytogenet. 2008. [Google Scholar] [CrossRef]
- Liehr, T.; Ewers, E.; Kosyakova, N.; Klaschka, V.; Rietz, F.; Wagner, R.; Weise, A. Handling small supernumerary marker chromosomes in prenatal diagnostics. Expert Rev. Mol. Diagn. 2009, 9, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Dutta, U.R.; Vempally, S.; Ranganath, P.; Dalal, A. A novel combined 15q11.2 duplication and a bisatellited supernumerary marker derived from chromosome 22: Molecular characterization of the marker. Gene 2014, 539, 162–167. [Google Scholar] [CrossRef]
- Therman, E.; Susman, B.; Denniston, C. The nonrandom participation of human acrocentric chromosomes in Robertsonian translocations. Ann. Hum. Genet. 1989, 53, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Jarmuz-Szymczak, M.; Janiszewska, J.; Szyfter, K.; Shaffer, L.G. Narrowing the localization of the region breakpoint in most frequent Robertsonian translocations. Chromosom. Res. 2014, 22, 517–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porokhovnik, L.; Gerton, J.L. Ribosomal DNA-connecting ribosome biogenesis and chromosome biology. Chromosom. Res. 2019, 27, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan-Jordan, J.; Simons, A.; Schmid, M. An international system for human cytogenomic nomenclature (2016). Cytogenet Genome Res. 2016, 149, 1–140. [Google Scholar]
- Waye, J.S.; Willard, H.F. Human β satellite DNA: Genomic organization and sequence definition of a class of highly repetitive tandem DNA. Proc. Natl. Acad. Sci. USA 1989, 86, 6250–6254. [Google Scholar] [CrossRef]
- Stults, D.M.; Killen, M.W.; Pierce, H.H.; Pierce, A.J. Genomic architecture and inheritance of human ribosomal RNA gene clusters. Genome Res. 2008, 18, 13–18. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; McQuillan, C.; Page, S.L.; Choo, K.H.A.; Shaffer, L.G. Identification and characterization of satellite III subfamilies to the acrocentric chromosomes. Chromosom. Res. 2001, 9, 223–233. [Google Scholar] [CrossRef]
- Sullivan, B.A.; Jenkins, L.S.; Karson, E.M.; Leana-Cox, J.; Schwartz, S. Evidence for structural heterogeneity from molecular cytogenetic analysis of dicentric Robertsonian translocations. Am. J. Hum. Genet. 1996, 59, 167–175. [Google Scholar]
- Higgins, M.J.; Wang, H.; Shtromas, I.; Haliotis, T.; Roder, J.C.; Holden, J.J.A.; White, B.N. Organization of a repetitive human 1.8 kb KpnI sequence localized in the heterochromatin of chromosome 15. Chromosoma 1985, 93, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Smeets, D.F.C.M.; Merkx, G.F.M.; Hopman, A.H.M. Chromosome 15p marker D15Z1 frequently maps to the short arm of other D-group chromosomes. Hum. Genet. 1992, 88, 365. [Google Scholar] [CrossRef] [PubMed]
- Stergianou, K.; Gould, C.P.; Waters, J.J.; Hultén, M. High population incidence of the 15p marker D15Z1 mapping to the short arm of one homologue 14. Hum. Genet. 1992, 88, 364. [Google Scholar] [CrossRef] [PubMed]
- Cockwell, A.E.; Jacobs, P.A.; Crolla, J.A. Distribution of the D15Z1 copy number polymorphism. Eur. J. Hum. Genet. 2007, 15, 441–445. [Google Scholar] [CrossRef]
- Floutsakou, I.; Agrawal, S.; Nguyen, T.T.; Seoighe, C.; Ganley, A.R.D.; McStay, B. The shared genomic architecture of human nucleolar organizer regions. Genome Res. 2013, 23, 2003–2012. [Google Scholar] [CrossRef] [Green Version]
- Jarmuz, M.; Glotzbach, C.D.; Bailey, K.A.; Bandyopadhyay, R.; Shaffer, L.G. The evolution of satellite III DNA subfamilies among primates. Am. J. Hum. Genet. 2007, 80, 495–501. [Google Scholar] [CrossRef]
- Choo, K.H.A.; Earle, E.; Vissel, B.; Kalitsis, P. A chromosome 14-specific human satellite III DNA subfamily that shows variable presence on different chromosomes 14. Am. J. Hum. Genet. 1992, 50, 706–716. [Google Scholar]
- Frias, S.; Ramos, S.; Molina, B.; Del Castillo, V.; Mayén, D.G. Detection of mosaicism in lymphocytes of parents of free trisomy 21 offspring. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2002, 520, 25–37. [Google Scholar] [CrossRef]
- Forero, C.M.; Lucena, Q.E.; Esteban Pérez, C. Frecuencia de mosaicismos de baja proporción del cromosoma x en parejas con antecedente de aborto recurrente. Rev. Ciencias la Salud 2006, 4, 23–45. [Google Scholar]
- Guttenbach, M.; Koschorz, B.; Bernthaler, U.; Grimm, T.; Schmid, M. Sex chromosome loss and aging: In situ hybridization studies on human interphase nuclei. Am. J. Hum. Genet. 1995, 57, 1143–1150. [Google Scholar]
- Kuo, P.L.; Guo, H.R. Mechanism of recurrent spontaneous abortions in women with mosaicism of X-chromosome aneuploidies. Fertil. Steril. 2004, 82, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, T.A.; Crolla, J.A. Molecular cytogenetic characterization of three familial cases of satellited Y chromosomes. Hum. Genet. 1993, 91, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Velissariou, V.; Sismani, C.; Christopoulou, S.; Kaminopetros, P.; Hatzaki, A.; Evangelidou, P.; Koumbaris, G.; Bartsocas, C.S.; Stylianidou, G.; Skordis, N.; et al. Loss of the Y chromosome PAR2 region and additional rearrangements in two familial cases of satellited Y chromosomes: Cytogenetic and molecular analysis. Eur. J. Med. Genet. 2007, 50, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.S.; Gogineni, S.K.; Kleyman, S.M.; Conte, R.A. Characterisation of a satellited non-fluorescent Y chromosome (Y(nfqs)) by FISH. J. Med. Genet. 1997, 34, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Kühl, H.; Röttger, S.; Heilbronner, H.; Enders, H.; Schempp, W. Loss of the Y chromosomal PAR2-region in four familial cases of satellited Y chromosomes (Yqs). Chromosom. Res. 2001, 9, 215–222. [Google Scholar] [CrossRef]
- Burk, R.D.; Szabo, P.; O’Brien, S.; Nash, W.G.; Yu, L.; Smith, K.D. Organization and chromosomal specificity of autosomal homologs of human Y chromosome repeated DNA. Chromosoma 1985, 92, 225–233. [Google Scholar] [CrossRef]
- Rahman, M.M.; Bashamboo, A.; Prasad, A.; Pathak, D.; Ali, S. Organizational variation of DYZ1 repeat sequences on the human Y chromosome and its diagnostic potentials. DNA Cell Biol. 2004, 23, 561–571. [Google Scholar] [CrossRef]
- Branco, M.R.; Pombo, A. Intermingling of chromosome territories in interphase suggests role in translocations and transcription-dependent associations. PLoS Biol. 2006, 4. [Google Scholar] [CrossRef]
- Sherman, S.L.; Lamb, N.E.; Feingold, E. Relationship of recombination patterns and maternal age among non-disjoined chromosomes 21. Biochem. Soc. Trans. 2006, 34, 578–580. [Google Scholar] [CrossRef] [Green Version]
- Ramos, S.; Molina, B.; Grether, P.; Mayén, D.G.; Castro, O.; Ángeles, M.; Rodríguez, R.; Frías, S. Translocaciones crípticas de cromosomas acrocéntricos en muestras de tejido de aborto aneuploide. Acta Pediatr. Mex. 2015, 36, 374–382. [Google Scholar] [CrossRef]
- Coppedè, F. Risk factors for Down syndrome. Arch. Toxicol. 2016. [Google Scholar] [CrossRef]
- Flores-Ramírez, F.; Palacios-Guerrero, C.; García-Delgado, C.; Morales-Jiménez, A.B.; Arias-Villegas, C.M.; Cervantes, A.; Morán-Barroso, V.F. Cytogenetic Profile in 1,921 Cases of Trisomy 21 Syndrome. Arch. Med. Res. 2015, 46, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Gruhn, J.R.; Zielinska, A.P.; Shukla, V.; Blanshard, R.; Capalbo, A.; Cimadomo, D.; Nikiforov, D.; Chan, A.C.; Newnham, L.J.; Vogel, I.; et al. Chromosome errors in human eggs shape natural fertility over reproductive life span. Science 2019, 365, 1466–1469. [Google Scholar] [PubMed] [Green Version]
- Oliver, T.R.; Middlebrooks, C.D.; Tinker, S.W.; Allen, E.G.; Bean, L.J.H.; Begum, F.; Feingold, E.; Chowdhury, R.; Cheung, V.; Sherman, S.L. An examination of the relationship between hotspots and recombination associated with chromosome 21 nondisjunction. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, N.; Bakshi, S.; Muthuswamy, S.; Agarwal, S. Young mothers and higher incidence of maternal meiosis-I non- disjunction: Interplay of environmental exposure and genetic alterations during halt phase in trisomy 21. Reprod. Toxicol. 2018, 79, 1–7. [Google Scholar] [CrossRef]
- Korzeneva, I.B.; Kostuyk, S.V.; Ershova, E.S.; Skorodumova, E.N.; Zhuravleva, V.F.; Pankratova, G.V.; Volkova, I.V.; Stepanova, E.V.; Porokhovnik, L.N.; Veiko, N.N. Human circulating ribosomal DNA content significantly increases while circulating satellite III (1q12) content decreases under chronic occupational exposure to low-dose gamma- neutron and tritium beta-radiation. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2016, 791–792, 49–60. [Google Scholar] [CrossRef]
- Van Sluis, M.; McStay, B. Nucleolar reorganization in response to rDNA damage. Curr. Opin. Cell Biol. 2017, 46, 81–86. [Google Scholar] [CrossRef]
- Ewers, E.; Yoda, K.; Hamid, A.B.; Weise, A.; Manvelyan, M.; Liehr, T. Centromere activity in dicentric small supernumerary marker chromosomes. Chromosom. Res. 2010, 18, 555–562. [Google Scholar] [CrossRef]
- Reddy, K.S.; Sulcova, V. The mobile nature of acrocentric elements illustrated by three unusual chromosome variants. Hum. Genet. 1998, 102, 653–662. [Google Scholar] [CrossRef]
- McGowan-Jordan, J.; Simons, A.; Schmid, M. ISCN 2016. Cytogenet. Genome Res. 2016, 149, 1–139. [Google Scholar]
- Cannings, C.; Edwards, A.W.F. Natural selection and the de Finetti diagram. Ann. Hum. Genet. 1968, 31, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Elston, R.C.; Forthofer, R. Testing for Hardy-Weinberg equilibrium in small samples. Biometrics 1977, 33, 536–547. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | GTG 1 Karyotype | Carrier of D15Z1 Polymorphism |
|---|---|---|
| Couples with healthy offspring | mos 45,X[4]/46,XX[89] | - |
| 46,XX,inv(8)(q13q24.1) | - | |
| Couples with aneuploid offspring | mos 47,XY,+21[3]/46,XY[62] | - |
| mos 45,X[3]/46,XY[40] | 13pvar(D15Z1+) |
| Group | n | Age ± SD | Mothers Aged ≤34 | Mothers Aged ≤34 with Extra D15Z1 | Mothers Aged ≥35 | Mothers Aged ≥35 with Extra D15Z1 |
|---|---|---|---|---|---|---|
| Mothers with healthy offspring | 75 | 25.9 ± 5.3 | 72 | 14 (19.4%) 1 | 3 | 0 (0%) 2 |
| Mothers with aneuploid offspring | 87 | 30.4 ± 7.3 | 61 | 15 (24.6%) 1 | 26 | 5 (19.2%) 2 |
| Individuals | n | With Extra D15Z1 | % | More than One Extra D15Z1 1 |
|---|---|---|---|---|
| With healthy offspring | 150 | 21/150 2 | 14 | 4/21 |
| With aneuploid offspring | 174 | 46/174 2 | 26.4 | 4/47 |
| Total | 324 | 67/324 | 20.7 | 8/68 |
| Individuals | Carriers Extra D15Z1 (%) | D15Z1 Extra-Signal Receptor Chromosomes (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Single Extra Signal | Multiple Extra Signals | ||||||||||
| 13 | 14 | 21 | 22 | 13 × 2 | 13 × 2 +14 | 13 + 14 | 13 + 21 | 14 × 2 | 14 + 21 | ||
| With healthy offspring n = 150 | 21 (14) | 6/21 (28.6) | 10/21 (47.6) | 0/21 | 1/21 (4.8) | 3/21 (14.3) | 0/21 | 0/21 | 1/21 (4.8) | 0/21 | 0/21 |
| Healthy offspring n = 10 | 5 (50%) | 1/5 (20) | 3/5 (60) | 0/5 | 0/5 | 0/5 | 0/5 | 1/5 (20) | 0/5 | 0/5 | 0/5 |
| With aneuploid offspring n = 174 | 46 (26.4) | 28/46 (60.8) | 10/46 (21.7) | 4/46 (8.7) | 0/46 | 0/46 | 1/46 (2.2) | 1/46 (2.2) | 0/46 | 1/46 (2.2) | 1/46 (2.2) |
| Aneuploid offspring n = 73 | 16 1 (21.9) | 8/16 (50) | 3/16 (18.75) | 2/16 (12.5) | 0/16 | 1/16 (6.25) | 0/16 | 2/16 (12.5) | 0/16 | 0/16 | 0/16 |
| Total n = 407 Frequency per individual | 88/407 (21.6) ~1/5 | 43 (10.6) 1/10 | 26 (6.4) 1/16 | 6 (1.5) 1/68 | 1 (0.25) 1/407 | 4 (1.0) 1/102 | 1 (0.25) 1/407 | 4 (1.0) 1/102 | 1 (0.25) 1/407 | 1 (0.25) 1/407 | 1 (0.25) 1/407 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, S.; Rodríguez, R.; Castro, O.; Grether, P.; Molina, B.; Frias, S. Presence of 15p Marker D15Z1 on the Short Arm of Acrocentric Chromosomes is Associated with Aneuploid Offspring in Mexican Couples. Int. J. Mol. Sci. 2019, 20, 5251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215251
Ramos S, Rodríguez R, Castro O, Grether P, Molina B, Frias S. Presence of 15p Marker D15Z1 on the Short Arm of Acrocentric Chromosomes is Associated with Aneuploid Offspring in Mexican Couples. International Journal of Molecular Sciences. 2019; 20(21):5251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215251
Chicago/Turabian StyleRamos, Sandra, Rebeca Rodríguez, Oscar Castro, Patricia Grether, Bertha Molina, and Sara Frias. 2019. "Presence of 15p Marker D15Z1 on the Short Arm of Acrocentric Chromosomes is Associated with Aneuploid Offspring in Mexican Couples" International Journal of Molecular Sciences 20, no. 21: 5251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215251