Blood Testis Barrier and Somatic Cells Impairment in a Series of 35 Adult Klinefelter Syndrome Patients


Abstract
:1. Introduction
2. Results
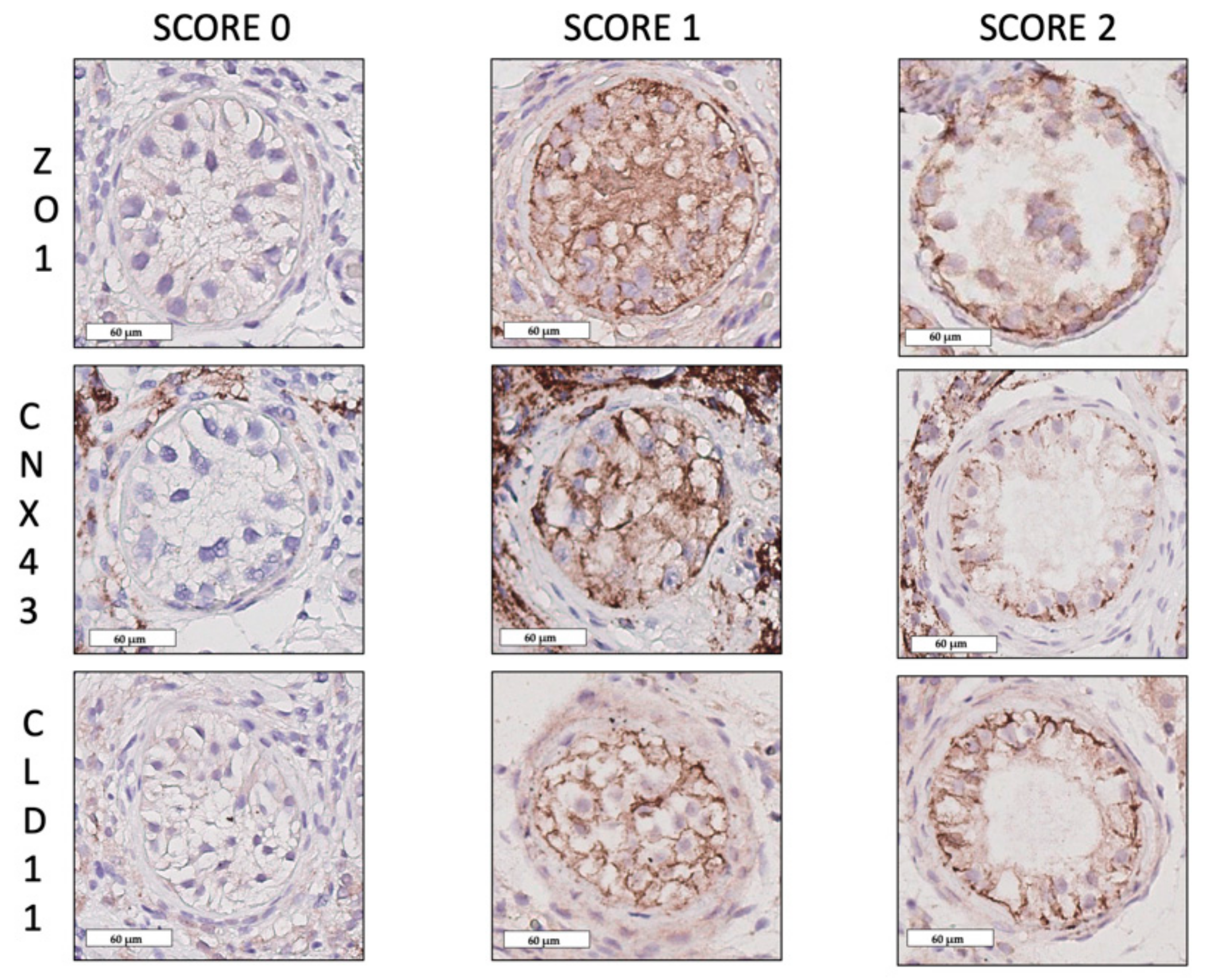
2.1. Blood–Testis Barrier Protein Expression Patterns
2.2. Sertoli Cell Maturation and Function
2.3. Leydig Cells Maturation and Function
3. Discussion
4. Materials and Methods
4.1. Source of Human Tissue
4.2. Immunohistochemical Procedures
4.3. Immunohistochemical Evaluation
4.4. Blood–Testis Barrier
4.5. Sertoli Cells
4.6. Leydig Cells
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| KS | Klinefelter Syndrome |
| GCs | Germ Cells |
| SSCs | Spermatogonial Stem Cells |
| LCs | Leydig Cells |
| TESE | Testicular Sperm Extraction |
| STs | Seminiferous Tubules |
| SCs | Sertoli Cells |
| AMH | Anti-Müllerian Hormone |
| AR | Androgen Receptors |
| BTB | Blood–Testis Barrier |
| CLD11 | Claudin-11 |
| CNX43 | Connexin-43 |
| ZO1 | Zonula Occludens-1 |
| INSL3 | Insulin-Like Peptide 3 |
| LH | Luteinizing Hormone |
| StAR | Steroidogenic Acute Regulatory Protein |
| SCO | Sertoli Cell Only |
| NSP | Normal Spermatogenesis |
| GDNF | Glial Cell Line Derived Neurotrophic Factor |
References
- Bojesen, A.; Juul, S.; Gravholt, C.H. Prenatal and postnatal prevalence of Klinefelter syndrome: A national registry study. J. Clin. Endocrinol. Metab. 2003, 88, 622–626. [Google Scholar] [CrossRef]
- Aksglaede, L.; Link, K.; Giwercman, A.; Jorgensen, N.; Skakkebaek, N.E.; Juul, A. 47,XXY Klinefelter syndrome: Clinical characteristics and age-specific recommendations for medical management. Am. J. Med. Genet. C. Semin. Med. Genet. 2013, 163, 55–63. [Google Scholar] [CrossRef]
- Aksglaede, L.; Wikstrom, A.M.; Rajpert-De Meyts, E.; Dunkel, L.; Skakkebaek, N.E.; Juul, A. Natural history of seminiferous tubule degeneration in Klinefelter syndrome. Hum. Reprod. 2006, 12, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Gies, I.; De Schepper, J.; Goossens, E.; Van Saen, D.; Pennings, G.; Tournaye, H. Spermatogonial stem cell preservation in boys with Klinefelter syndrome: To bank or not to bank, that’s the question. Fertil. Steril. 2012, 98, 28–49. [Google Scholar] [CrossRef]
- Van Saen, D.; Gies, I.; De Schepper, J.; Tournaye, H.; Goossens, E. Can pubertal boys with Klinefelter syndrome benefit from spermatogonial stem cell banking? Hum. Reprod. 2012, 27, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Van Saen, D.; Vloeberghs, V.; Gies, I.; Mateizel, I.; Sermon, K.; De Schepper, J.; Tournaye, H.; Goossens, E. When does germ cell loss and fibrosis occur in patients with Klinefelter syndrome? Hum. Reprod. 2018, 33, 1009–1022. [Google Scholar] [CrossRef]
- Rives, N.; Milazzo, J.P.; Perdrix, A.; Castanet, M.; Joly, H.G.; Sibert, L.; Bironneau, A.; Way, A.; Mace, B. The feasibility of fertility preservation in adolescents with Klinefelter syndrome. Hum. Reprod. 2013, 28, 1468–1479. [Google Scholar] [CrossRef]
- Rives, N.; Rives, A.; Rondanino, C.; Castanet, M.; Cuny, A.; Sibert, L. Fertility preservation in klinefelter syndrome patients during the transition period. Endocr. Dev. 2018, 33, 149–157. [Google Scholar]
- Giudice, M.G.; Del Vento, F.; Wyns, C. Male fertility preservation in DSD, XXY, pre-gonadotoxic treatments – update, methods, ethical issues, current outcomes, future directions. Best Pract. Res. Clin. Endocrinol. Metab 2019. [Google Scholar] [CrossRef] [PubMed]
- Mruk, D.D.; Cheng, C.Y. Sertoli-Sertoli and Sertoli-germ cell interactions and their significance in germ cell movement in the seminiferous epithelium during spermatogenesis. Endocr. Rev. 2004, 25, 747–806. [Google Scholar] [CrossRef]
- Aksglaede, L.; Christiansen, P.; Sorensen, K.; Boas, M.; Linneberg, A.; Main, K.M.; Andersson, A.M.; Skakkebaek, N.E.; Juul, A. Serum concentrations of Anti-Mullerian Hormone (AMH) in 95 patients with Klinefelter syndrome with or without cryptorchidism. Acta Paediatr. 2011, 100, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Wikstrom, A.M.; Hoei-Hansen, C.E.; Dunkel, L.; Rajpert-De Meyts, E. Immunoexpression of androgen receptor and nine markers of maturation in the testes of adolescent boys with Klinefelter syndrome: Evidence for degeneration of germ cells at the onset of meiosis. J. Clin. Endocrinol. Metab. 2007, 92, 714–719. [Google Scholar] [CrossRef]
- O’Hara, L.; Smith, L.B. Androgen receptor roles in spermatogenesis and infertility. Best. Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 595–605. [Google Scholar] [CrossRef]
- De Michele, F.; Poels, J.; Smedt, D.F.; Ambroise, J.; Vermeulen, M.; Gruson, D.; Wyns, C. In-vitro formation of the blood-testis barrier during long-term organotypic culture of human prepubertal tissue: Comparison with a large cohort of pre/peripubertal boys. Mol. Hum. Reprod. 2018, 24, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Steed, E.; Balda, M.S.; Matter, K. Dynamics and functions of tight junctions. Trends Cell. Biol. 2010, 20, 142–149. [Google Scholar] [CrossRef]
- Harris, A.L. Connexin channel permeability to cytoplasmic molecules. Prog. Biophys. Mol. Biol. 2007, 94, 120–143. [Google Scholar] [CrossRef]
- Fink, C.; Weigel, R.; Hembes, T.; Lauke-Wettwer, H.; Kliesch, S.; Bergmann, M.; Brehm, R.H. Altered expression of ZO-1 and ZO-2 in Sertoli cells and loss of blood-testis barrier integrity in testicular carcinoma in situ. Neoplasia 2006, 8, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Gerber, J.; Heinrich, J.; Brehm, R. Blood-testis barrier and Sertoli cell function: Lessons from SCCx43KO mice. Reproduction 2016, 151, 15–27. [Google Scholar] [CrossRef]
- Lie, P.P.; Cheng, C.Y.; Mruk, D.D. Signalling pathways regulating the blood-testis barrier. Int. J. Biochem. Cell Biol. 2013, 45, 621–625. [Google Scholar] [CrossRef]
- Kotula-Balak, M.; Hejmej, A.; Sadowska, J.; Bilinska, B. Connexin 43 expression in human and mouse testes with impaired spermatogenesis. Eur. J. Histochem. 2007, 51, 261–268. [Google Scholar]
- Wikstrom, A.M.; Dunkel, L. Klinefelter syndrome. Best. Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 239–250. [Google Scholar]
- Wikstrom, A.M.; Bay, K.; Hero, M.; Andersson, A.M.; Dunkel, L. Serum insulin-like factor 3 levels during puberty in healthy boys and boys with Klinefelter syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 4705–4708. [Google Scholar] [CrossRef]
- Van Saen, D.; Sanchez, P.J.; van der Werff ten Bosch, J.; Tournaye, H.; Gossens, E. Is the protein expression window during testicular development affected in patients at risk for stem cell loss? Hum. Reprod. 2015, 30, 2859–2870. [Google Scholar] [CrossRef]
- D’Aurora, M.; Ferlin, A.; Di Nicola, M.; Garolla, A.; De Toni, L.; Franchi, S.; Palka, G.; Foresta, C.; Stuppia, L.; Gatta, V. Deregulation of sertoli and leydig cells function in patients with Klinefelter syndrome as evidenced by testis transcriptome analysis. BMC Genom. 2015, 16, 156. [Google Scholar] [CrossRef] [PubMed]
- Rohayem, J.; Fricken, R.; Caeloth, K.; Mallidis, C.; Wistuba, J.; Krallmann, C.; Zitzmannn, M.; Kliesch, S. Age and markers of Leydig cell function, but not of Sertoli cell function predict the success of sperm retrieval in adolescents and adults with Klinefelter’s syndrome. Andrology 2015, 3, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Maiburg, M.; Repping, S.; Giltay, J. The genetic origin of Klinefelter syndrome and its effect on spermatogenesis. Fertil. Steril. 2012, 98, 253–260. [Google Scholar] [CrossRef]
- Foresta, C.; Galeazzi, C.; Bettella, A.; Marin, P.; Rossato, M.; Garolla, A.; Ferlin, A. Analysis of meiosis in intratesticular germ cells from subjects affected by classic Klinefelter’s syndrome. J. Clin. Endocrinol. Metab. 1999, 84, 3807–3810. [Google Scholar]
- Sciurano, R.B.; Luna, Hisano, C.V.; Rahn, M.I.; Brugo, O.S.; Rey, V.G.; Coco, R.; Solari, A.J. Focal spermatogenesis originates in euploid germ cells in classical Klinefelter patients. Hum. Reprod. 2009, 24, 2353–2560. [Google Scholar] [CrossRef] [PubMed]
- Mazaud, G.S.; Meugenier, E.; Pesenti, S.; Wu, X.; Vidal, H.; Gow, A.; Battistoni-Magueresse, L.B. Claudin 11 deficiency in mice results in loss of the Sertoli cell epithelial phenotype in the testis. Biol. Reprod. 2010, 82, 202–213. [Google Scholar] [CrossRef]
- Chiba, K.; Yamaguchi, K.; Ando, M.; Miyake, H.; Fujisawa, M. Expression pattern of testicular claudin-11 in infertile men. Urology 2012, 80, 13–17. [Google Scholar] [CrossRef]
- Haverfield, J.T.; Meachem, S.J.; O’Bryan, M.K.; McLachlan, R.I.; Stanton, P.G. Claudin-11 and connexin-43 display altered spatial patterns of organization in men with primary seminiferous tubule failure compared with controls. Fertil. Steril. 2013, 100, 658–666. [Google Scholar] [CrossRef]
- Nah, W.H.; Lee, J.E.; Park, H.J.; Park, N.C.; Gye, M.C. Claudin-11 expression increased in spermatogenic defect in human testes. Fertil. Steril. 2011, 95, 385–388. [Google Scholar] [CrossRef]
- Raleigh, D.; O’Donnell, L.; Southwick, G.J.; de Kretser, D.M.; McLachlan, R.I. Stereological analysis of the human testis after vasectomy indicates impairment of spermatogenic efficiency with increasing obstructive interval. Fertil. Steril. 2004, 81, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- McDonald, S.W. Cellular responses to vasectomy. Int. Rev. Cytol. 2000, 199, 295–339. [Google Scholar] [PubMed]
- Turner, T.T.; D’Addario, D.A.; Howards, S.S. Effects of vasectomy on the blood-testis barrier of the hamster. J. Reprod. Fertil. 1979, 55, 323–328. [Google Scholar]
- McCabe, M.J.; Tarulli, G.A.; Laven, L.G.; Matthiesson, K.L.; Meachem, S.J.; Mclachlan, R.I.; Dinger, M.E.; Stanton, P.G. Gonadotropin suppression in men leads to a reduction in claudin-11 at the Sertoli cell tight junction. Hum. Reprod. 2016, 31, 875–886. [Google Scholar] [CrossRef] [Green Version]
- Lui, W.Y.; Mruk, D.; Lee, W.M.; Cheng, C.Y. Sertoli cell tight junction dynamics: Their regulation during spermatogenesis. Biol. Reprod. 2003, 68, 1087–1097. [Google Scholar] [CrossRef]
- Russell, L.D.; Peterson, R.N. Determination of the elongate spermatid-Sertoli cell ratio in various mammals. J. Reprod. Fertil. 1984, 70, 635–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naughton, C.K.; Jain, S.; Strickland, A.M.; Gupta, A.; Milbrandt, J. Glial cell-line derived neurotrophic factor-mediated RET signaling regulates spermatogonial stem cell fate. Biol. Reprod. 2006, 74, 314–321. [Google Scholar] [CrossRef]
- Lee, M.M.; Donahoe, P.K.; Hasegawa, T.; Silverman, B.; Crist, G.B.; Best, S.; Hasegawa, Y.; Noto, R.A.; Schoenfeld, D.; MacLaughlin, D.T. Mullerian inhibiting substance in humans: Normal levels from infancy to adulthood. J. Clin. Endocrinol. Metab. 1996, 81, 571–576. [Google Scholar] [PubMed] [Green Version]
- Wikstrom, A.M.; Raivio, T.; Hadziselimovic, F.; Wikstrom, S.; Tuuri, T.; Dunkel, L. Klinefelter syndrome in adolescence: Onset of puberty is associated with accelerated germ cell depletion. J. Clin. Endocrinol. Metab. 2004, 89, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Trabado, S.; Maione, L.; Bry-Gauillard, H.; Affres, H.; Salenave, S.; Sarfati, J.; Bouvattier, C.; Delemer, B.; Chanson, P.; Le Bouc, Y.; et al. Insulin-like peptide 3 (INSL3) in men with congenital hypogonadotropic hypogonadism/Kallmann syndrome and effects of different modalities of hormonal treatment: A single-center study of 281 patients. J. Clin. Endocrinol. Metab. 2014, 99, 268–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lottrup, G.; Nielsen, J.E.; Maroun, L.L.; Moller, L.M.A.; Yassin, M.; Leffers, H.; Skakkebaek, N.E.; Rajpert-de, M.E. Expression patterns of DLK1 and INSL3 identify stages of Leydig cell differentiation during normal development and in testicular pathologies, including testicular cancer and Klinefelter syndrome. Hum. Reprod. 2014, 29, 1637–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksglaede, L.; Skakkebaek, N.E.; Juul, A. Abnormal sex chromosome constitution and longitudinal growth: Serum levels of insulin-like growth factor (IGF)-I, IGF binding protein-3, luteinizing hormone, and testosterone in 109 males with 47,XXY, 47,XYY, or sex-determining region of the Y chromosome (SRY)-positive 46,XX karyotypes. J. Clin. Endocrinol. Metab. 2008, 93, 169–176. [Google Scholar]
- D’Aurora, M.; Ferlin, A.; Garolla, A.; Franchi, S.; D’Onofrio, L.; Trubiani, O.; Palka, G.; Foresta, C.; Stuppia, L.; Gatta, V. Testis Transcriptome Modulation in Klinefelter Patients with Hypospermatogenesis. Sci. Rep. 2017, 7, 45729. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N Patients | N TESE or μTESE | N Biopsies |
|---|---|---|
| 35 KS | 5 unilateral μTESE 30 bilateral μTESE | 65 |
| 9 NSP | 9 unilateral TESE | 9 |
| 28 SCO | 2 unilateral μTESE 26 bilateral μTESE | 54 |
| Antibody | Target Cell | Dilution | Mono/Polyclonal | Producer | Reference Number |
|---|---|---|---|---|---|
| AMH | Sertoli cells | 1/200 | Monoclonal | Serotec | MICA2246 |
| GDNF | Sertoli cells | 1/150 | Polyclonal | Sigma-Aldrich | SAB1405863 |
| AR | Sertoli cells | 1/100 | Monoclonal | DAKO | AR441 |
| CLD11 | BTB | 1/400 | Polyclonal | Santa Cruz | SC25711 |
| CNX43 | BTB | 1/4000 | Polyclonal | Abcam | AB 11370 |
| ZO1 | BTB | 1/1500 | Polyclonal | Sigma-Aldrich | HPA001637 |
| INSL3 | Leydig cells | 1/2000 | Polyclonal | Sigma-Aldrich | HPA028615 |
| StAR | Leydig cells | 1/400 | Polyclonal | Sigma-Aldrich | HPA027318 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giudice, M.G.; Vermeulen, M.; Wyns, C. Blood Testis Barrier and Somatic Cells Impairment in a Series of 35 Adult Klinefelter Syndrome Patients. Int. J. Mol. Sci. 2019, 20, 5717. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225717
Giudice MG, Vermeulen M, Wyns C. Blood Testis Barrier and Somatic Cells Impairment in a Series of 35 Adult Klinefelter Syndrome Patients. International Journal of Molecular Sciences. 2019; 20(22):5717. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225717
Chicago/Turabian StyleGiudice, Maria Grazia, Maxime Vermeulen, and Christine Wyns. 2019. "Blood Testis Barrier and Somatic Cells Impairment in a Series of 35 Adult Klinefelter Syndrome Patients" International Journal of Molecular Sciences 20, no. 22: 5717. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225717