Identification of a Major QTL (qRRs-10.1) That Confers Resistance to Ralstonia solanacearum in Pepper (Capsicum annuum) Using SLAF-BSA and QTL Mapping

 ,
,
Abstract
:1. Introduction
2. Results
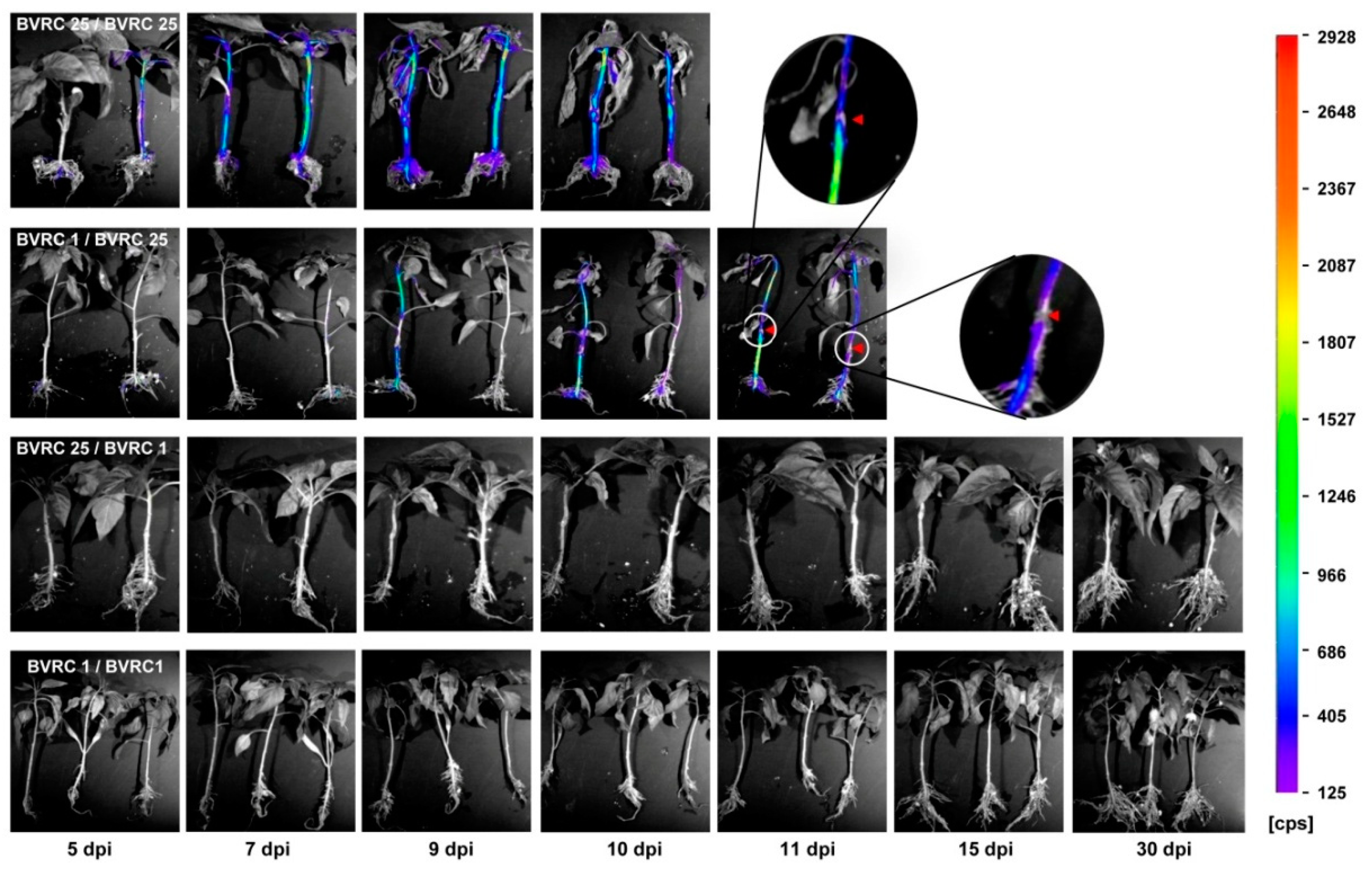
2.1. Visualization of Bioluminescent R. solanacearum Colonization of Grafted Plants
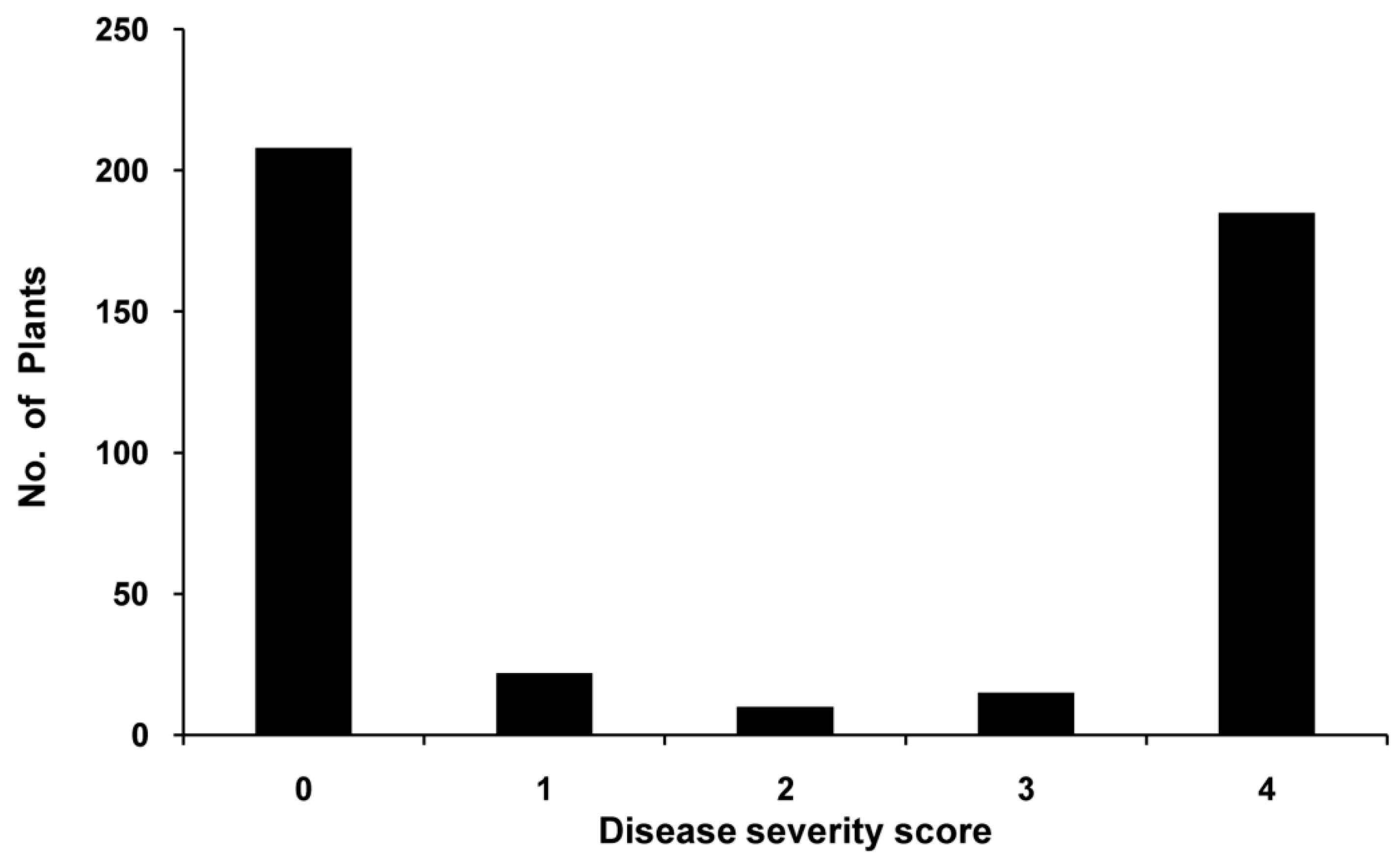
2.2. BW Resistance Inheritance in the Pepper Line BVRC 1
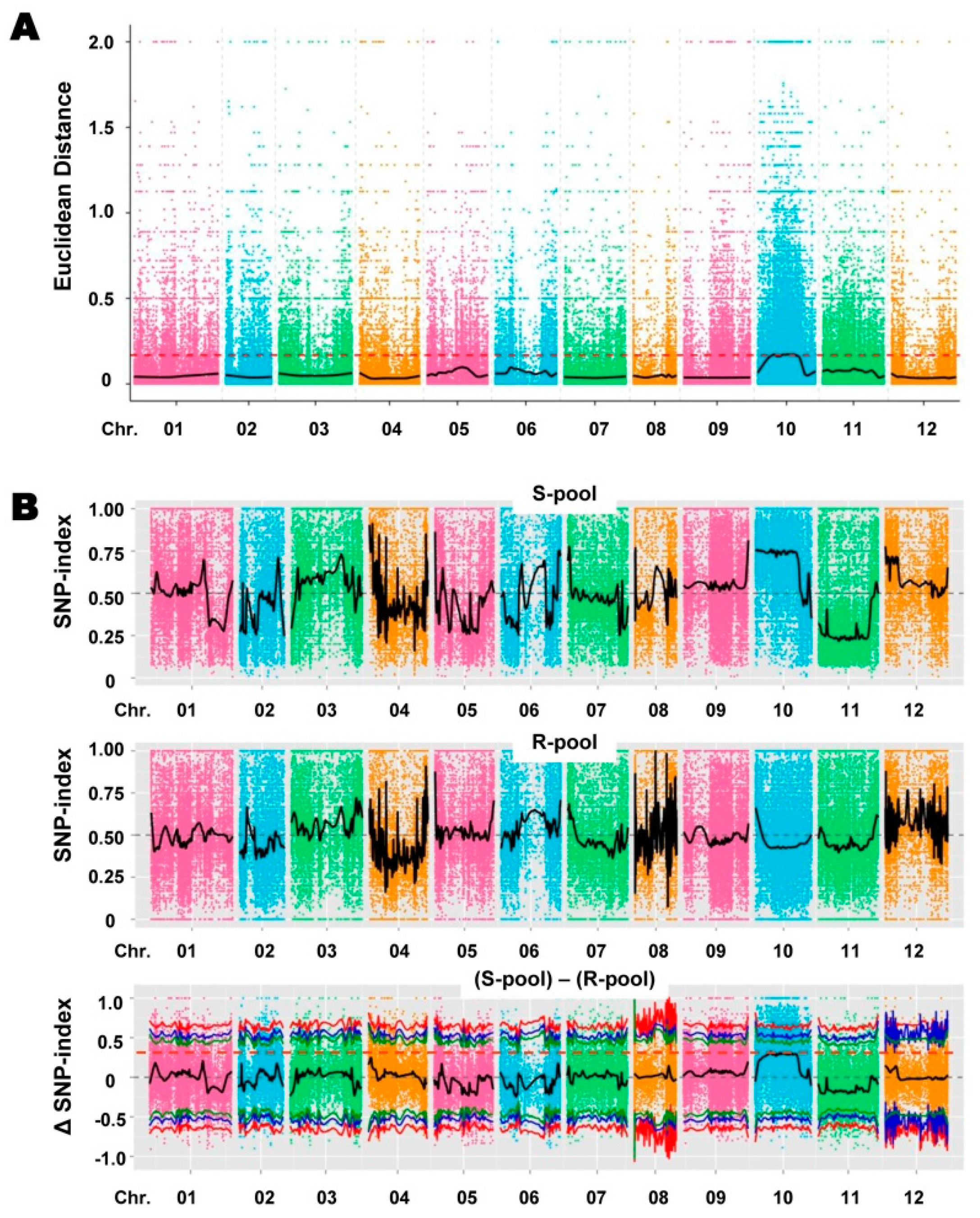
2.3. Association Analysis Based on Euclidean Distance (ED) and SNP Index
2.4. Construction of a Genetic Linkage Map of Chromosome 10
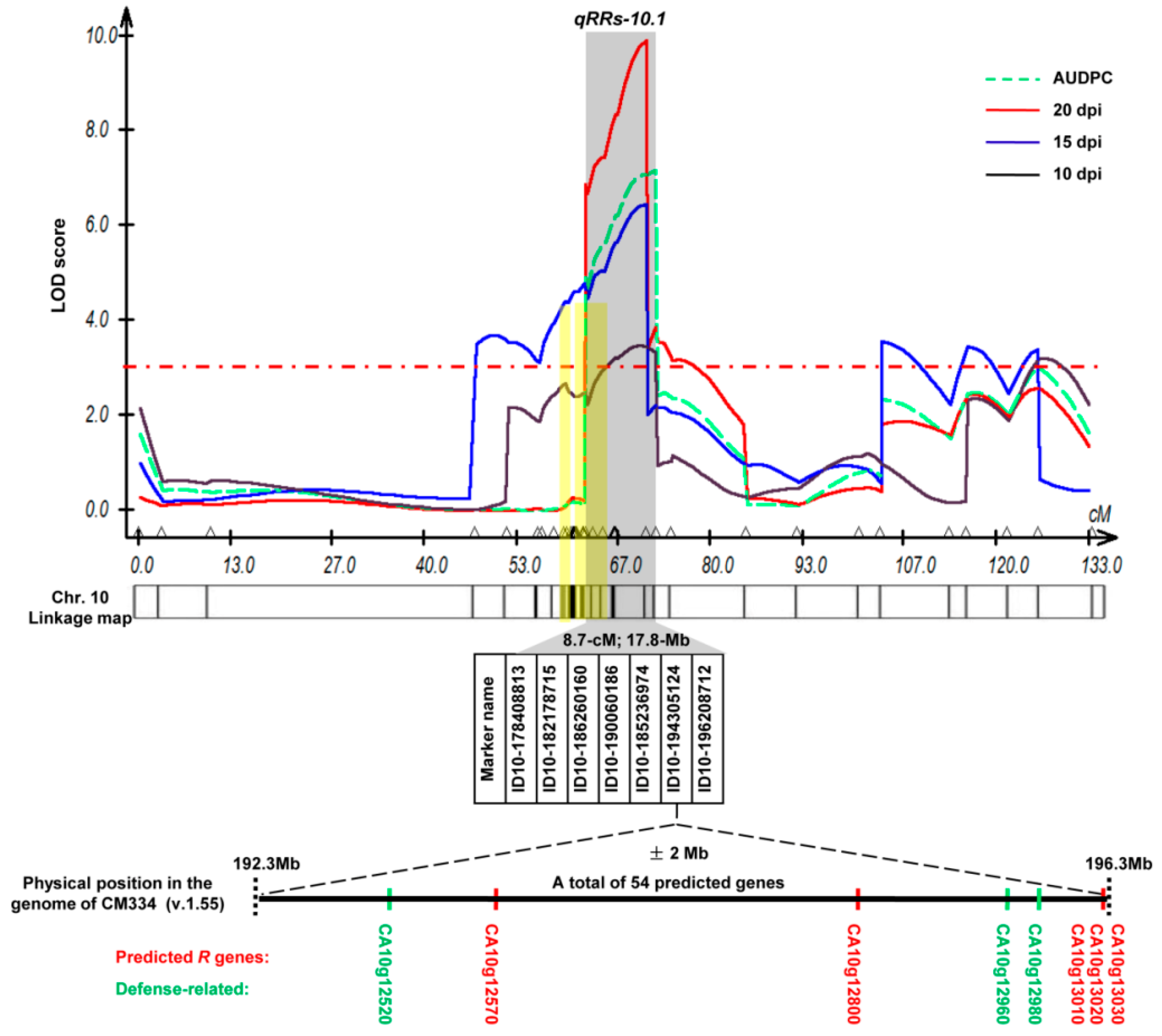
2.5. Confirmation of the BW Resistance-Associated Regions by QTL Mapping
2.6. Annotated Resistance-Related Genes and SNPs within the qRRs-10.1 Locus
3. Discussion
3.1. BVRC 1 Is a Good Rootstock for BW Resistance
3.2. Like Most BW-Resistant Pepper Lines, BVRC 1 Has Partially Dominant Resistance
3.3. qRRs-10.1 Is a Partially Dominant Major Resistance QTL on Chromosome 10 of the Pepper Line BVRC 1
3.4. qRRs-10.1 Candidate Genes
4. Materials and Methods
4.1. Grafting Experiments
4.2. Inoculum Preparation and Inoculation
4.3. Bioluminescence Imaging
4.4. Genetic Population Construction
4.5. Inoculation and Disease Evaluation
4.6. DNA Extraction
4.7. SLAF Library Construction and Sequencing
4.8. SLAF-seq Data Analysis
4.9. Association Mapping Analysis
4.10. Insertion/Deletion (InDel) and SNP Marker Development
4.11. Genotyping of the Population
4.12. Linkage Map Construction and QTL Mapping
4.13. Annotation of the Predicted Genes and Sequence Variants in the QTL Region
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| QTL | quantitative trait locus |
| SNP | single-nucleotide polymorphism |
| InDel | insertion/deletion |
| SLAF | specific-locus amplified fragment |
| BSA | bulked segregant analysis |
| ED | Euclidean distance |
| BW | bacterial wilt |
| CIM | composite interval mapping |
| DSI | disease severity index |
| dpi | days post-inoculation |
| AUDPC | area under the disease progress curve |
| LOD | log-likelihood |
| NGS | next-generation sequencing |
| DR | dominance ratio |
| GO | gene ontology |
| KEGG | kyoto encyclopedia of genes and genomes |
| COG | cluster of orthologous groups of proteins |
| KASPar | kompetitive allele-specific PCR |
| LRR | leucine-rich repeat |
| NB | nucleotide binding |
| PAMP | pathogen-associated molecular pattern |
| MAS | marker-assisted selection |
References
- Du, H.S.; Chen, B.; Zhang, X.F.; Zhang, F.L.; Miller, S.A.; Rajashekara, G.; Xu, X.L.; Geng, S.S. Evaluation of Ralstonia solanacearum infection dynamics in resistant and susceptible pepper lines using bioluminescence imaging. Plant Dis. 2017, 101, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Hayward, A.C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Peeters, N.; Guidot, A.; Vailleau, F.; Valls, M. Ralstonia solanacearum, a widespread bacterial plant pathogen in the post-genomic era. Mol. Plant Pathol. 2013, 14, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Pan, Z.C.; Prior, P.; Xu, J.S.; Zhang, Z.; Zhang, H.; Zhang, L.Q.; He, L.Y.; Feng, J. Genetic diversity of Ralstonia solanacearum strains from China. Eur. J. Plant Pathol. 2009, 125, 641–653. [Google Scholar] [CrossRef]
- Genin, S.; Denny, T.P. Pathogenomics of the Ralstonia solanacearum species complex. Annu. Rev. Phytopathol. 2012, 50, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Huet, G. Breeding for resistances to Ralstonia solanacearum. Front. Plant Sci. 2014, 5, 715. [Google Scholar] [CrossRef]
- Murakoshi, S.; Takahashi, M. Trials of some control of tomato bacterial wilt caused by Pseudomonas solanacearum. Bull. Kanagawa Hortic. Exp. Stn. 1984, 31, 50–56. [Google Scholar]
- Kim, B.S.; Cheung, J.D.; Cha, Y.S.; Hwang, H.S. Resistance to bacterial wilt of introduced peppers. Korean J. Plant Pathol. 1998, 14, 217–219. [Google Scholar]
- Shekhawat, G.S.; Gadewar, A.V.; Chakrabarti, S.K. Spontaneous phenotypic reversion from afluidal to fluidal state in strains of Pseudomonas solanacearum. Bact. Wilt Newsl. 1992, 8, 5–9. [Google Scholar]
- Tran, N.H.; Kim, B.-S. Inheritance of resistance to bacterial wilt (Ralstonia solanacearum) in Pepper (Capsicum annuum L.). Hortic. Environ. Biotechnol. 2010, 51, 431–439. [Google Scholar]
- Naresh, P.; Meenu, K.; Acharya, G.C.; Reddy, A.C.; Reddy, D.C.L. Genetics and molecular markers for resistance to major soil borne pathogens in chilli (Capsicum annuum L.). Res. J. Biotechnol. 2019, 14, 101–105. [Google Scholar]
- Matsunaga, H.; Monma, S. Varietal differences in resistance to bacterial wilt in related species of Capsicum annuum. Capsicum Eggplant Newsl. 1995, 14, 60–61. [Google Scholar]
- Lafortune, D.; Beramis, M.; Daubeze, A.M.; Boissot, N.; Palloix, A. Partial resistance of pepper to bacterial wilt is oligogenic and stable under tropical conditions. Plant Dis. 2005, 89, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Mimura, Y.; Yoshikawa, M.; Hirai, M. Pepper accession LS2341 is highly resistant to Ralstonia solanacearum strains from Japan. HortScience 2009, 44, 2038–2040. [Google Scholar] [CrossRef]
- Tsuro, M.; Minamiyama, Y.; Hirai, M. QTL analysis for bacterial wilt resistance in Japanese pepper (Capsicum annuum L.). Breed. Res. 2007, 9, 111–115. [Google Scholar]
- Matsunaga, H.; Sato, T.; Monma, S. Inheritance of bacterial wilt resistance in the sweet pepper cv. Mie-Midori. In Proceedings of the 10th Eucarpia Meeting on Genetics and Breeding of Capsicum and Eggplant, Avignon, France, 7–11 September 1998; p. 172. [Google Scholar]
- Mimura, Y.; Kageyama, T.; Minamiyama, Y.; Hirai, M. QTL analysis for resistance to Ralstonia solanacearum in Capsicum accession LS2341. J. Jpn. Soc. Hortic. Sci. 2009, 78, 307–313. [Google Scholar] [CrossRef]
- Thakur, P.P.; Mathew, D.; Nazeem, P.A.; Abida, P.S.; Indira, P.; Girija, D.; Shylaja, M.R.; Valsala, A. Identification of allele specific AFLP markers linked with bacterial wilt Ralstonia solanacearum (Smith) Yabuuchi et al. resistance in hot peppers (Capsicum annuum L.). Physiol. Mol. Plant Pathol. 2014, 87, 19–24. [Google Scholar] [CrossRef]
- Carmeille, A.; Caranta, C.; Dintinger, J.; Prior, P.; Luisetti, J.; Besse, P. Identification of QTLs for Ralstonia solanacearum race 3-phylotype II resistance in tomato. Theor. Appl. Genet. 2006, 113, 110–121. [Google Scholar] [CrossRef]
- Thoquet, P.; Olivier, J.; Sperisen, C.; Rogowsky, P.; Prior, P.; Anais, G.; Mangin, B.; Nazer, R.; Grimsley, N. Polygenic resistance of tomato plants to bacterial wilt in the French West Indies. Mol. Plant Microbe Interact. 1996, 9, 837–842. [Google Scholar] [CrossRef]
- Wang, J.F.; Olivier, J.; Thoquet, P.; Mangin, B.; Sauviac, L.; Grimsley, N.H. Resistance of tomato line Hawaii7996 to Ralstonia solanacearum Pss4 in Taiwan is controlled mainly by a major strain-specific locus. Mol. Plant Microbe Interact. 2000, 13, 6–13. [Google Scholar] [CrossRef]
- Wang, J.F.; Ho, F.I.; Truong, H.T.H.; Huang, S.M.; Balatero, C.H.; Dittapongpitch, V.; Hidayati, N. Identification of major QTLs associated with stable resistance of tomato cultivar ‘Hawaii 7996’ to Ralstonia solanacearum. Euphytica 2013, 190, 241–252. [Google Scholar] [CrossRef]
- Kim, B.; Hwang, I.S.; Lee, H.J.; Lee, J.M.; Seo, E.; Choi, D.; Oh, C.S. Identification of a molecular marker tightly linked to bacterial wilt resistance in tomato by genome-wide SNP analysis. Theor. Appl. Genet. 2018, 131, 1017–1030. [Google Scholar] [CrossRef]
- Michelmore, R.W.; Paran, I.; Kesseli, R.V. Identification of markers linked to disease-resistance genes by bulked segregant analysis—A rapid method to detec markers in specific genomic regions by using segregating populations. Proc. Natl. Acad. Sci. USA 1991, 88, 9828–9832. [Google Scholar] [CrossRef]
- Schneeberger, K.; Ossowski, S.; Lanz, C.; Juul, T.; Petersen, A.H.; Nielsen, K.L.; Jorgensen, J.E.; Weigel, D.; Andersen, S.U. SHOREmap: Simultaneous mapping and mutation identification by deep sequencing. Nat. Methods 2009, 6, 550–551. [Google Scholar] [CrossRef]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M.; et al. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef]
- Sun, X.; Liu, D.; Zhang, X.; Li, W.; Liu, H.; Hong, W.; Jiang, C.; Guan, N.; Ma, C.; Zeng, H.; et al. SLAF-seq: An efficient method of large-scale de novo SNP discovery and genotyping using high-throughput sequencing. PLoS ONE 2013, 8, e0058700. [Google Scholar] [CrossRef]
- Geng, X.; Jiang, C.; Yang, J.; Wang, L.; Wu, X.; Wei, W. Rapid identification of candidate genes for seed weight using the SLAF-Seq method in Brassica napus. PLoS ONE 2016, 11, e0147580. [Google Scholar] [CrossRef]
- Xu, F.; Sun, X.; Chen, Y.; Huang, Y.; Tong, C.; Bao, J. Rapid identification of major QTLs associated with rice grain weight and their utilization. PLoS ONE 2015, 10, e0122206. [Google Scholar] [CrossRef]
- Xu, X.; Ji, J.; Xu, Q.; Qi, X.; Weng, Y.; Chen, X. The major-effect quantitative trait locus CsARN6.1 encodes an AAA ATPase domain-containing protein that is associated with waterlogging stress tolerance by promoting adventitious root formation. Plant J. 2018, 93, 917–930. [Google Scholar] [CrossRef]
- Zhang, X.F.; Wang, G.Y.; Chen, B.; Du, H.S.; Zhang, F.L.; Zhang, H.Y.; Wang, Q.; Geng, S.S. Candidate genes for first flower node identified in pepper using combined SLAF-seq and BSA. PLoS ONE 2018, 13, e0194071. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chao, J.; Cheng, X.; Wang, R.; Sun, B.; Wang, H.; Luo, S.; Xu, X.; Wu, T.; Li, Y. Mapping of a novel race specific resistance gene to Phytophthora root rot of pepper (Capsicum annuum) using bulked segregant analysis combined with specific length amplified fragment sequencing strategy. PLoS ONE 2016, 11, e0151401. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Wang, S.; Liu, J.; Pan, B.; Diao, W.; Ge, W.; Gao, C.; Snyder, J.C. Rapid identification of QTLs underlying resistance to cucumber mosaic virus in pepper (Capsicum frutescens). Theor. Appl. Genet. 2017, 130, 41–52. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.T.; Demarest, B.L.; Bisgrove, B.W.; Gorsi, B.; Su, Y.C.; Yost, H.J. MMAPPR: Mutation mapping analysis pipeline for pooled RNA-seq. Genome Res. 2013, 23, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gomez, L.; Boller, T. FLS2: An LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Knoth, C.; Eulgem, T. The oomycete response gene LURP1 is required for defense against Hyaloperonospora parasitica in Arabidopsis thaliana. Plant J. 2008, 55, 53–64. [Google Scholar] [CrossRef]
- Louws, F.J.; Rivard, C.L.; Kubota, C. Grafting fruiting vegetables to manage soilborne pathogens, foliar pathogens, arthropods and weeds. Sci. Hortic. 2010, 127, 127–146. [Google Scholar] [CrossRef]
- Rivard, C.L.; O’Connell, S.; Peet, M.M.; Welker, R.M.; Louws, F.J. Grafting tomato to manage bacterial wilt caused by Ralstonia solanacearum in the Southeastern United States. Plant Dis. 2012, 96, 973–978. [Google Scholar] [CrossRef] [Green Version]
- Ben, C.; Debelle, F.; Berges, H.; Bellec, A.; Jardinaud, M.F.; Anson, P.; Huguet, T.; Gentzbittel, L.; Vailleau, F. MtQRRS1, an R-locus required for Medicago truncatula quantitative resistance to Ralstonia solanacearum. New Phytol. 2013, 199, 758–772. [Google Scholar] [CrossRef] [PubMed]
- Eynck, C.; Koopmann, B.; Karlovsky, P.; von Tiedemann, A. Internal resistance in winter oilseed rape inhibits systemic spread of the vascular pathogen Verticillium longisporum. Phytopathology 2009, 99, 802–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vailleau, F.; Sartorel, E.; Jardinaud, M.F.; Chardon, F.; Genin, S.; Huguet, T.; Gentzbittel, L.; Petitprez, M. Characterization of the interaction between the bacterial wilt pathogen Ralstonia solanacearum and the model legume plant Medicago truncatula. Mol. Plant Microbe Interact. 2007, 20, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Korol, A.; Shirak, A.; Cnaani, A.; Hallerman, E.M. Detection and analysis of quantitative trait loci (QTL) for economic traits in aquatic species. Aquac. Genome Technol. 2007, 31, 169. [Google Scholar]
- Bent, A.F.; Mackey, D. Elicitors, effectors, and R genes: The new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol. 2007, 45, 399–436. [Google Scholar] [CrossRef] [Green Version]
- Jones, N.; Ougham, H.; Thomas, H.; Pasakinskiene, I. Markers and mapping revisited: Finding your gene. New Phytol. 2009, 183, 935–966. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Godiard, L.; Sauviac, L.; Torii, K.U.; Grenon, O.; Mangin, B.; Grimsley, N.H.; Marco, Y. ERECTA, an LRR receptor-like kinase protein controlling development pleiotropically affects resistance to bacterial wilt. Plant J. 2003, 36, 353–365. [Google Scholar] [CrossRef]
- Deslandes, L.; Olivier, J.; Theulieres, F.; Hirsch, J.; Feng, D.X.; Bittner-Eddy, P.; Beynon, J.; Marco, Y. Resistance to Ralstonia solanacearum in Arabidopsis thaliana is conferred by the recessive RRS1-R gene, a member of a novel family of resistance genes. Proc. Natl. Acad. Sci. USA 2002, 99, 2404–2409. [Google Scholar] [CrossRef] [Green Version]
- Van der Linden, L.; Bredenkamp, J.; Naidoo, S.; Fouche-Weich, J.; Denby, K.J.; Genin, S.; Marco, Y.; Berger, D.K. Gene-for-gene tolerance to bacterial wilt in Arabidopsis. Mol. Plant Microbe Interact. 2013, 26, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Narusaka, M.; Shirasu, K.; Noutoshi, Y.; Kubo, Y.; Shiraishi, T.; Iwabuchi, M.; Narusaka, Y. RRS1 and RPS4 provide a dual resistance-gene system against fungal and bacterial pathogens. Plant J. 2009, 60, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Huh, S.U.; Cevik, V.; Ding, P.T.; Duxbury, Z.; Ma, Y.; Tomlinson, L.; Sarris, P.F.; Jones, J.D.G. Protein-protein interactions in the RPS4/RRS1 immune receptor complex. PLoS Pathog. 2017, 13, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashikawa, I.; Hayashi, N.; Yamane, H.; Kanamori, H.; Wu, J.; Matsumoto, T.; Ono, K.; Yano, M. Two adjacent nucleotide-binding site-leucine-rich repeat class genes are required to confer Pikm-specific rice blast resistance. Genetics 2008, 180, 2267–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Roux, C.; Huet, G.; Jauneau, A.; Camborde, L.; Trémousaygue, D.; Kraut, A.; Zhou, B.; Levaillant, M.; Adachi, H.; Yoshioka, H.; et al. A receptor pair with an integrated decoy converts pathogen disabling of transcriptor factors to immunity. Cell 2015, 161, 1074–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarris, P.F.; Duxbury, Z.; Huh, S.U.; Ma, Y.; Segonzac, C.; Sklenar, J.; Derbyshire, P.; Cevik, V.; Rallapalli, G.; Saucet, S.B.; et al. A plant immune receptor detects pathogen effectors that target WRKY transcriptors. Cell 2015, 161, 1089–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.J.; Kim, K.H.; Shim, S.; Yoon, M.Y.; Sun, S.; Kim, M.Y.; Van, K.; Lee, S.H. Genome-wide mapping of NBS-LRR genes and their association with disease resistance in soybean. BMC Plant Biol. 2012, 12, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilet-Nayel, M.L.; Prosperi, J.M.; Hamon, C.; Lesne, A.; Lecointe, R.; Le Goff, I.; Herve, M.; Deniot, G.; Delalande, M.; Huguet, T.; et al. AER1, a major gene conferring resistance to Aphanomyces euteiches in Medicago truncatula. Phytopathology 2009, 99, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinapidou, E.; Williams, K.; Nott, L.; Bahkt, S.; Tor, M.; Crute, I.; Bittner-Eddy, P.; Beynon, J. Two TIR:NB:LRR genes are required to specify resistance to Peronospora parasitica isolate Cala2 in Arabidopsis. Plant J. 2004, 38, 898–909. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.B.; Kang, W.H.; Hoang Ngoc, H.; Yeom, S.I.; An, J.T.; Kim, S.; Kang, M.Y.; Kim, H.J.; Jo, Y.D.; Ha, Y.; et al. Divergent evolution of multiple virus-resistance genes from a progenitor in Capsicum spp. New Phytol. 2017, 213, 886–899. [Google Scholar] [CrossRef]
- Tai, T.H.; Dahlbeck, D.; Clark, E.T.; Gajiwala, P.; Pasion, R.; Whalen, M.C.; Stall, R.E.; Staskawicz, B.J. Expression of the Bs2 pepper gene confers resistance to bacterial spot disease in tomato. Proc. Natl. Acad. Sci. USA 1999, 96, 14153–14158. [Google Scholar] [CrossRef] [Green Version]
- Kearney, B.; Staskawicz, B.J. Widespread distribution and fitness contribution of Xanthomonas campestris avirulence gene avrBs2. Nature 1990, 346, 385–386. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, S.; Iriarte, F.; Fan, Q.; da Silva, E.E.; Ritchie, L.; Nghi Song, N.; Freeman, J.H.; Stall, R.E.; Jones, J.B.; Minsavage, G.V.; et al. Transgenic expression of EFR and Bs2 genes for field management of bacterial wilt and bacterial spot of tomato. Phytopathology 2018, 108, 1402–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutrot, F.; Zipfel, C. Function, discovery, and exploitation of plant pattern recognition receptors for broad-spectrum disease resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef] [PubMed]
- Kelman, A. The relationship of pathogenicity of Pseudomonas solanacearum to colony appearance in a tetrazolium medium. Phytopathology 1954, 44, 693–695. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; University of California, College of Agriculture, Agricultural Experiment Station: Berkeley, CA, USA, 1950; Volume 347. [Google Scholar]
- Xu, X.; Rajashekara, G.; Paul, P.A.; Miller, S.A. Colonization of tomato seedlings by bioluminescent Clavibacter michiganensis subsp. michiganensis under different humidity regimes. Phytopathology 2012, 102, 177–184. [Google Scholar]
- Agricolae: Statistical Procedures for Agricultural Research. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 4 April 2019).
- Fulton, T.M.; Chunwongse, J.; Tanksley, S.D. Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep. 1995, 13, 207–209. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Su, T.B.; Li, P.R.; Yang, J.J.; Sui, G.L.; Yu, Y.J.; Zhang, D.S.; Zhao, X.Y.; Wang, W.H.; Wen, C.L.; Yu, S.C.; et al. Development of cost-effective single nucleotide polymorphism marker assays for genetic diversity analysis in Brassica rapa. Mol. Breed. 2018, 38. [Google Scholar] [CrossRef]
- Lander, E.S.; Green, P. Construction of multilocus genetic linkage maps in humans. Proc. Natl. Acad. Sci. USA 1987, 84, 2363–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Z.B. Precision mapping of quantitative trait loci. Genetics 1994, 136, 1457–1468. [Google Scholar] [PubMed]
- Wang, S.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer 2.5. Department of Statistics; North Carolina State University: Raleigh, NC, USA, 2006. [Google Scholar]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernet, A.; Hoisington, D.; Franco, J.; Isnard, M.; Jewell, D.; Jiang, C.; Marchand, J.L.; Reynaud, B.; Glaszmann, J.C.; de Leon, D.G. Genetic mapping of maize streak virus resistance from the Mascarene source. I. Resistance in line D211 and stability against different virus clones. Theor. Appl. Genet. 1999, 99, 524–539. [Google Scholar] [CrossRef] [PubMed]
- Stuber, C.W.; Edwards, M.D.; Wendel, J.F. Molecular marker-facilitated investigations of quantitative trait loci in maize. II. Factors influencing yield and its component traits. Crop Sci. 1987, 27, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.H.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.Y.; Li, J.Q.; Wu, S.F.; Zhu, Y.P.; Chen, Y.W.; He, F.C. Integrated nr database in protein annotation system and its localization. Comput. Eng. 2006, 32, 71–72. [Google Scholar]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.Y.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w(1118); iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generations a | Experimental Design for Disease Evaluation Assays | Resistance Components | ||||
|---|---|---|---|---|---|---|
| Test × Block × Plants | No. of Plants | Inoculation Time | DSI b | Wilt Rate (%) c | AUDPC d | |
| Ps | 3 × 1 × 20 − 25 | 68 | Summer of 2016 | 3.98 ± 0.04 | 100 | 42.92 ± 1.84 |
| Autumn of 2016 | ||||||
| Summer of 2017 | ||||||
| Pr | 3 × 1 × 15 − 25 | 66 | Summer of 2016 | 0.31 ± 0.24 | 4.58 ± 3.95 | 1.68 ± 1.28 |
| Autumn of 2016 | ||||||
| Summer of 2017 | ||||||
| F1 (♀Ps × ♂Pr) | 3 × 1 × 15 − 25 | 68 | Summer of 2016 | 1.76 ± 0.17 | 48.59 ± 5.38 | 14.56 ± 3.08 |
| Autumn of 2016 | ||||||
| Summer of 2017 | ||||||
| BC1Ps | 1 × 1 × 31 | 31 | Summer of 2017 | 2.71 | 71.97 | 23.08 |
| BC1Pr | 1 × 1 × 33 | 33 | Summer of 2017 | 0.86 | 24.94 | 7.2 |
| F2 | 1 × 1 × 440 | 440 | Autumn of 2016 | 1.88 | 47.78 | 18.36 |
| Chr ID | Start Position (bp) | End Position (bp) | Interval Size (Mb) |
|---|---|---|---|
| Pepper.v.1.55 Chr10 | 56,910,000 | 69,110,000 | ~12.20 |
| Pepper.v.1.55 Chr10 | 111,090,000 | 183,670,000 | ~72.58 |
| InDel Markers a | Genetic Distance (cM) | Composite Interval Mapping Results | Mean AUDPC of Three Genotypic Classes in F2 Plants g | ||||||
|---|---|---|---|---|---|---|---|---|---|
| LOD b | R2 (%) c | A d | D e | DR f | B:B | A:A | A:B | ||
| ID10-178408813 | 63.6 | 7.47 | 13.65 | 0.81 | −0.21 | 0.52 | 7.30 | 33.80 | 16.87 |
| ID10-182178715 | 64.9 | 8.12 | 14.00 | 0.86 | −0.20 | 0.46 | 7.69 | 33.97 | 17.71 |
| ID10-185236974 | 66.8 | 8.37 | 17.82 | 0.94 | −0.17 | 0.37 | 7.12 | 35.00 | 17.60 |
| ID10-186260160 | 66.5 | 8.37 | 17.82 | 0.87 | −0.21 | 0.47 | 7.04 | 34.91 | 17.74 |
| ID10-190060186 | 66.6 | 8.38 | 17.82 | 0.87 | −0.21 | 0.47 | 7.12 | 34.91 | 17.74 |
| ID10-194305124 | 71.0 | 9.93 | 19.01 | 0.97 | −0.10 | 0.21 | 6.36 | 35.52 | 18.05 |
| ID10-196208712 | 72.3 | 3.59 | 18.41 | 0.73 | −0.13 | 0.35 | 7.45 | 35.55 | 17.85 |
| Gene ID | Functional Annotation | Database a | No. of Non-Synonymous in Coding Region |
|---|---|---|---|
| CA10g12520 | Pathogenesis-related protein (PR-1) | NR; Swiss-Prot; KEGG: plant–pathogen interaction; | 0 |
| CA10g12570 | Tospovirus resistance protein E | CM334 (v.1.55); COG; Swiss-Prot | 0 |
| CA10g12800 | Putative disease resistance protein | CM334 (v.1.55); Pfam; Swiss-Prot | 0 |
| CA10g12960 | protein LURP-one-related 15-like | CM334 (v.1.55); Nr, Swiss-Prot; Pfam; GO: defense response; | 2 |
| CA10g12980 | protein LURP-one-related 15-like | CM334 (v.1.55); Nr, Swiss-Prot; Pfam, GO: defense response; | 1 |
| CA10g13010 | Disease resistance protein BS2 | CM334 (v.1.55); NR | 8 |
| CA10g13020 | Disease resistance protein BS2 | CM334 (v.1.55); Pfam; Swiss-Prot | 7 |
| CA10g13030 | NBS-coding resistance gene analog | CM334 (v.1.55); NR | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, H.; Wen, C.; Zhang, X.; Xu, X.; Yang, J.; Chen, B.; Geng, S. Identification of a Major QTL (qRRs-10.1) That Confers Resistance to Ralstonia solanacearum in Pepper (Capsicum annuum) Using SLAF-BSA and QTL Mapping. Int. J. Mol. Sci. 2019, 20, 5887. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235887
Du H, Wen C, Zhang X, Xu X, Yang J, Chen B, Geng S. Identification of a Major QTL (qRRs-10.1) That Confers Resistance to Ralstonia solanacearum in Pepper (Capsicum annuum) Using SLAF-BSA and QTL Mapping. International Journal of Molecular Sciences. 2019; 20(23):5887. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235887
Chicago/Turabian StyleDu, Heshan, Changlong Wen, Xiaofen Zhang, Xiulan Xu, Jingjing Yang, Bin Chen, and Sansheng Geng. 2019. "Identification of a Major QTL (qRRs-10.1) That Confers Resistance to Ralstonia solanacearum in Pepper (Capsicum annuum) Using SLAF-BSA and QTL Mapping" International Journal of Molecular Sciences 20, no. 23: 5887. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235887