Platelet Proteasome Activity and Metabolism Is Upregulated during Bacterial Sepsis

Abstract
:1. Introduction
2. Results
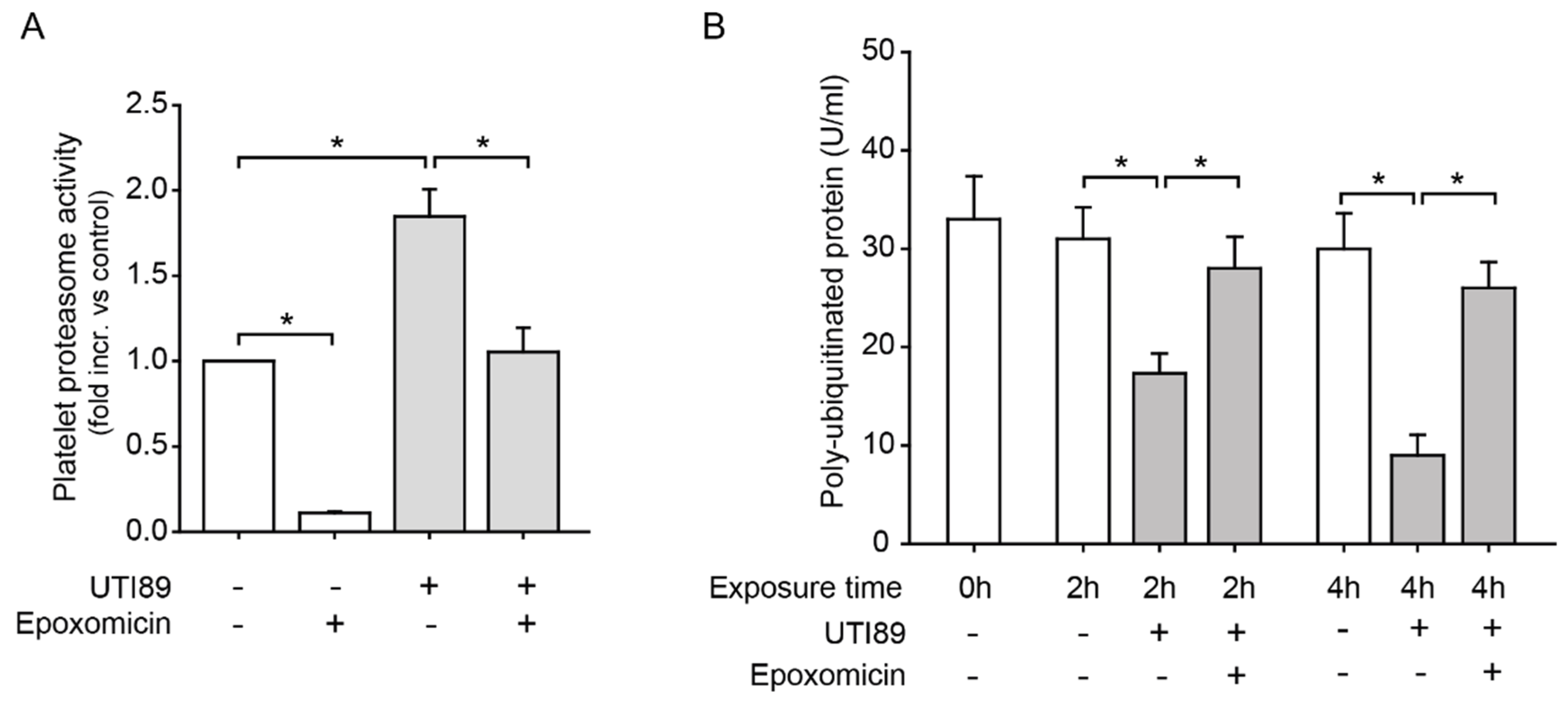
2.1. Platelet Proteasome Activity and Protein Metabolism is Increased in the Septic Milieu
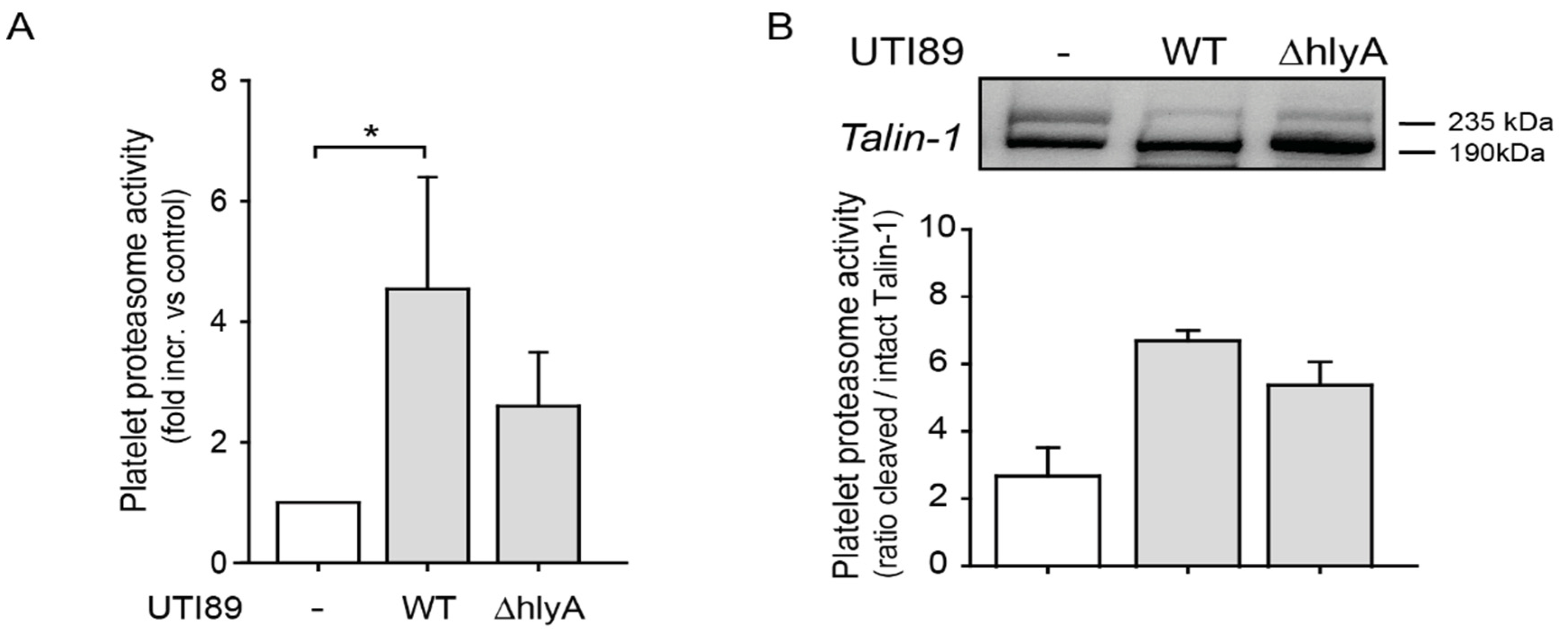
2.2. E. coli Exotoxin α-Hemolysin (hlyA) may be a Contributing Factor to Increased Proteasome Activity in Platelets
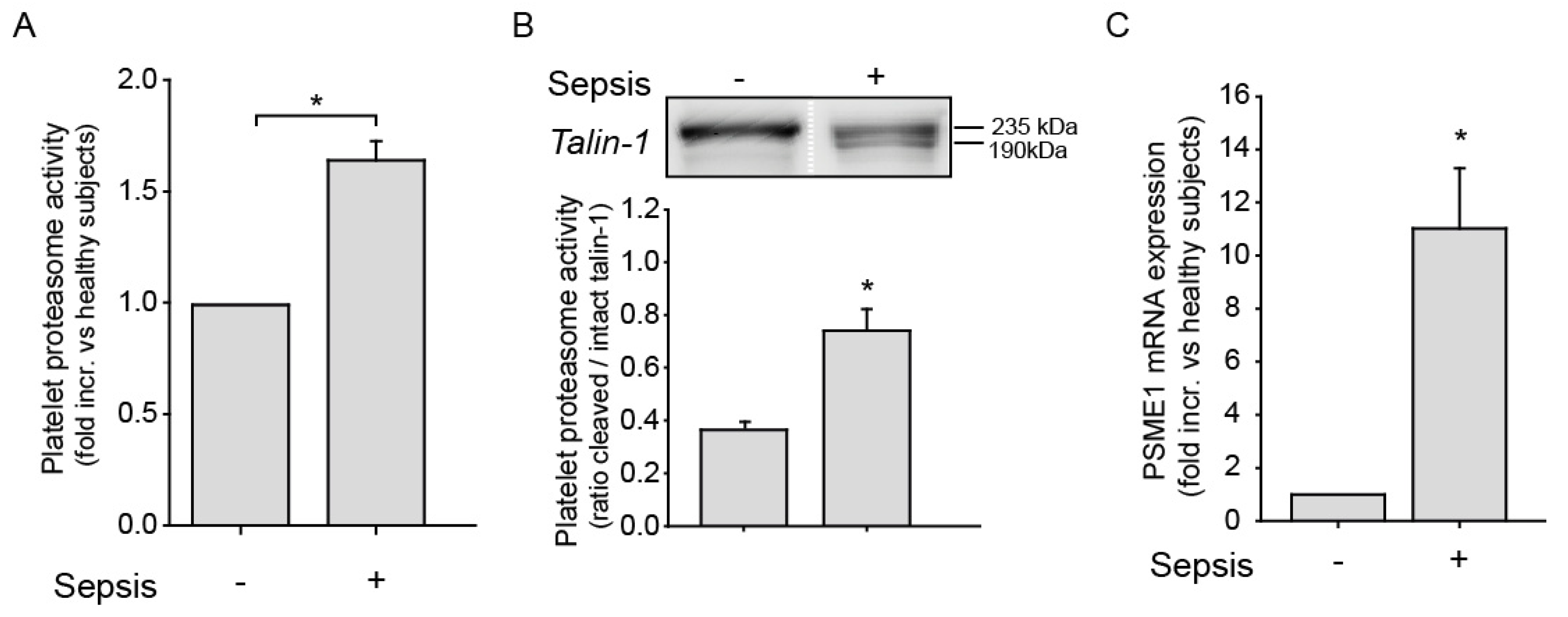
2.3. Platelet Proteasome Activity is Increased in Platelets from Sepsis Patients
2.4. Clinical Characteristics of Sepsis Patients
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Bacteria
4.3. Platelet Isolation and Preparation
4.4. RNA Isolation
4.5. Detection of Platelet Proteasome Activity
4.6. Quantification of Polyubiquitinated Protein in Human Platelets
4.7. Immunoblotting
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Assinger, A.; Schrottmaier, W.C.; Salzmann, M.; Rayes, J. Platelets in Sepsis: An Update on Experimental Models and Clinical Data. Front. Immunol. 2019, 10, 1687. [Google Scholar] [CrossRef] [PubMed]
- Nayak, M.K.; Kumar, K.; Dash, D. Regulation of proteasome activity in activated human platelets. Cell Calcium 2011, 49, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Grundler, K.; Rotter, R.; Tilley, S.; Pircher, J.; Czermak, T.; Yakac, M.; Gaitzsch, E.; Massberg, S.; Krotz, F.; Sohn, H.Y.; et al. The proteasome regulates collagen-induced platelet aggregation via nuclear-factor-kappa-B (NFkB) activation. Thromb. Res. 2016, 148, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Li, W.; Willard, B.; Silverstein, R.L.; McIntyre, T.M. Proteasome proteolysis supports stimulated platelet function and thrombosis. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Ferrington, D.A.; Gregerson, D.S. Immunoproteasomes: Structure, function, and antigen presentation. Prog. Mol. Biol. Transl. Sci. 2012, 109, 75–112. [Google Scholar] [PubMed]
- Hobler, S.C.; Williams, A.; Fischer, D.; Wang, J.J.; Sun, X.; Fischer, J.E.; Monaco, J.J.; Hasselgren, P.O. Activity and expression of the 20S proteasome are increased in skeletal muscle during sepsis. Am. J. Physiol. 1999, 277, R434–R440. [Google Scholar] [CrossRef] [PubMed]
- Klaude, M.; Fredriksson, K.; Tjader, I.; Hammarqvist, F.; Ahlman, B.; Rooyackers, O.; Wernerman, J. Proteasome proteolytic activity in skeletal muscle is increased in patients with sepsis. Clin. Sci. (Lond.) 2007, 112, 499–506. [Google Scholar] [CrossRef]
- Tanaka, K. The proteasome: Overview of structure and functions. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 12–36. [Google Scholar] [CrossRef]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef]
- Kraemer, B.F.; Weyrich, A.S.; Lindemann, S. Protein degradation systems in platelets. Thromb. Haemost. 2013, 110, 920–924. [Google Scholar] [CrossRef]
- Avcu, F.; Ural, A.U.; Cetin, T.; Nevruz, O. Effects of bortezomib on platelet aggregation and ATP release in human platelets, in vitro. Thromb. Res. 2008, 121, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Caturegli, P.; Takahashi, M.; Suzuki, K. New Insights into the Function of the Immunoproteasome in Immune and Nonimmune Cells. J. Immunol. Res. 2015, 2015, 541984. [Google Scholar] [CrossRef] [PubMed]
- Seifert, U.; Bialy, L.P.; Ebstein, F.; Bech-Otschir, D.; Voigt, A.; Schroter, F.; Prozorovski, T.; Lange, N.; Steffen, J.; Rieger, M.; et al. Immunoproteasomes preserve protein homeostasis upon interferon-induced oxidative stress. Cell 2010, 142, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Chapman, L.M.; Aggrey, A.A.; Field, D.J.; Srivastava, K.; Ture, S.; Yui, K.; Topham, D.J.; Baldwin, W.M., 3rd; Morrell, C.N. Platelets present antigen in the context of MHC class I. J. Immunol. 2012, 189, 916–923. [Google Scholar] [CrossRef]
- Klockenbusch, C.; Walsh, G.M.; Brown, L.M.; Hoffman, M.D.; Ignatchenko, V.; Kislinger, T.; Kast, J. Global proteome analysis identifies active immunoproteasome subunits in human platelets. Mol. Cell. Proteomics 2014, 13, 3308–3319. [Google Scholar] [CrossRef]
- Dahlmann, B. Role of proteasomes in disease. BMC Biochem. 2007, 8 (Suppl. 1), S3. [Google Scholar] [CrossRef]
- Kraemer, B.F.; Campbell, R.A.; Schwertz, H.; Franks, Z.G.; Vieira de Abreu, A.; Grundler, K.; Kile, B.T.; Dhakal, B.K.; Rondina, M.T.; Kahr, W.H.; et al. Bacteria differentially induce degradation of Bcl-xL, a survival protein, by human platelets. Blood 2012, 120, 5014–5020. [Google Scholar] [CrossRef]
- Ristow, L.C.; Welch, R.A. Hemolysin of uropathogenic Escherichia coli: A cloak or a dagger? Biochim. Biophys. Acta 2016, 1858, 538–545. [Google Scholar] [CrossRef]
- Mulvey, M.A.; Schilling, J.D.; Hultgren, S.J. Establishment of a persistent Escherichia coli reservoir during the acute phase of a bladder infection. Infect. Immun. 2001, 69, 4572–4579. [Google Scholar] [CrossRef]
- Wiles, T.J.; Dhakal, B.K.; Eto, D.S.; Mulvey, M.A. Inactivation of host Akt/protein kinase B signaling by bacterial pore-forming toxins. Mol. Biol. Cell 2008, 19, 1427–1438. [Google Scholar] [CrossRef]
- Ostrowska, H.; Ostrowska, J.K.; Worowski, K.; Radziwon, P. Human platelet 20S proteasome: Inhibition of its chymotrypsin-like activity and identification of the proteasome activator PA28. A preliminary report. Platelets 2003, 14, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Trugilho, M.R.O.; Hottz, E.D.; Brunoro, G.V.F.; Teixeira-Ferreira, A.; Carvalho, P.C.; Salazar, G.A.; Zimmerman, G.A.; Bozza, F.A.; Bozza, P.T.; Perales, J. Platelet proteome reveals novel pathways of platelet activation and platelet-mediated immunoregulation in dengue. PLoS Pathog. 2017, 13, e1006385. [Google Scholar] [CrossRef] [PubMed]
- Weyrich, A.S.; Elstad, M.R.; McEver, R.P.; McIntyre, T.M.; Moore, K.L.; Morrissey, J.H.; Prescott, S.M.; Zimmerman, G.A. Activated platelets signal chemokine synthesis by human monocytes. J. Clin. Invest. 1996, 97, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, B.F.; Campbell, R.A.; Schwertz, H.; Cody, M.J.; Franks, Z.; Tolley, N.D.; Kahr, W.H.; Lindemann, S.; Seizer, P.; Yost, C.C.; et al. Novel anti-bacterial activities of beta-defensin 1 in human platelets: Suppression of pathogen growth and signaling of neutrophil extracellular trap formation. PLoS Pathog. 2011, 7, e1002355. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics of Seven Patients with Bacterial Sepsis | |
|---|---|
| Age | 63 [IQR 49; 69] |
| Leukocyte count (1000/µL) | 16 [IQR 11.0; 21.0] |
| CRP level (mg/dl) | 25 [IQR 19.5; 28.5] |
| Procalcitonin (ng/mL) | 5.5 [IQR 4.0; 9.8] |
| Positive bacterial culture | 7 of 7 |
| Antibiotic treatment | 7 of 7 |
| SOFA score | 9 [IQR 6; 11] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grundler Groterhorst, K.; Mannell, H.; Pircher, J.; Kraemer, B.F. Platelet Proteasome Activity and Metabolism Is Upregulated during Bacterial Sepsis. Int. J. Mol. Sci. 2019, 20, 5961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235961
Grundler Groterhorst K, Mannell H, Pircher J, Kraemer BF. Platelet Proteasome Activity and Metabolism Is Upregulated during Bacterial Sepsis. International Journal of Molecular Sciences. 2019; 20(23):5961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235961
Chicago/Turabian StyleGrundler Groterhorst, Katharina, Hanna Mannell, Joachim Pircher, and Bjoern F Kraemer. 2019. "Platelet Proteasome Activity and Metabolism Is Upregulated during Bacterial Sepsis" International Journal of Molecular Sciences 20, no. 23: 5961. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235961