Genetic Restoration of Heme Oxygenase-1 Expression Protects from Type 1 Diabetes in NOD Mice

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
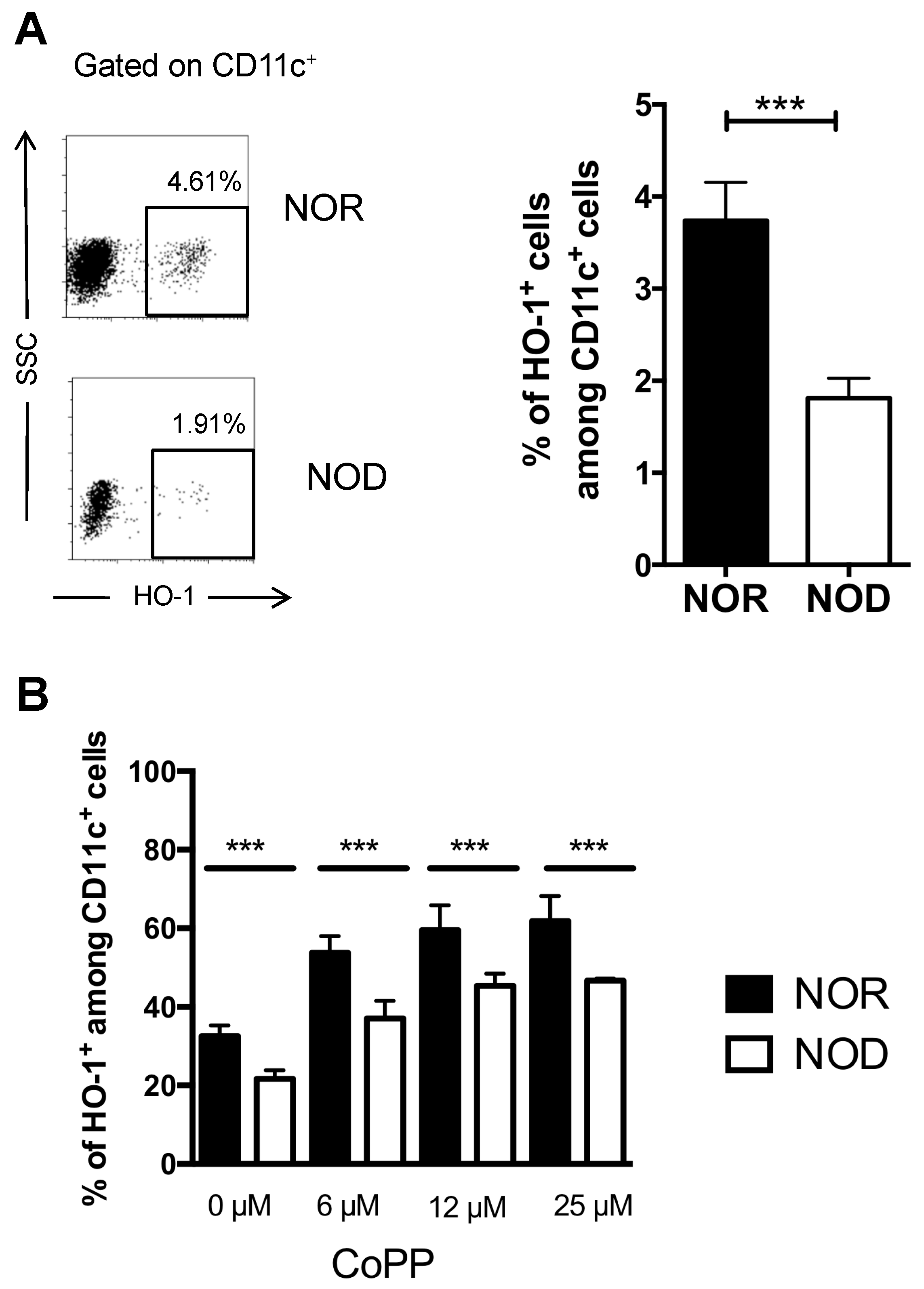
2.1. DCs from NOD Mice are Deficient in HO-1 Expression
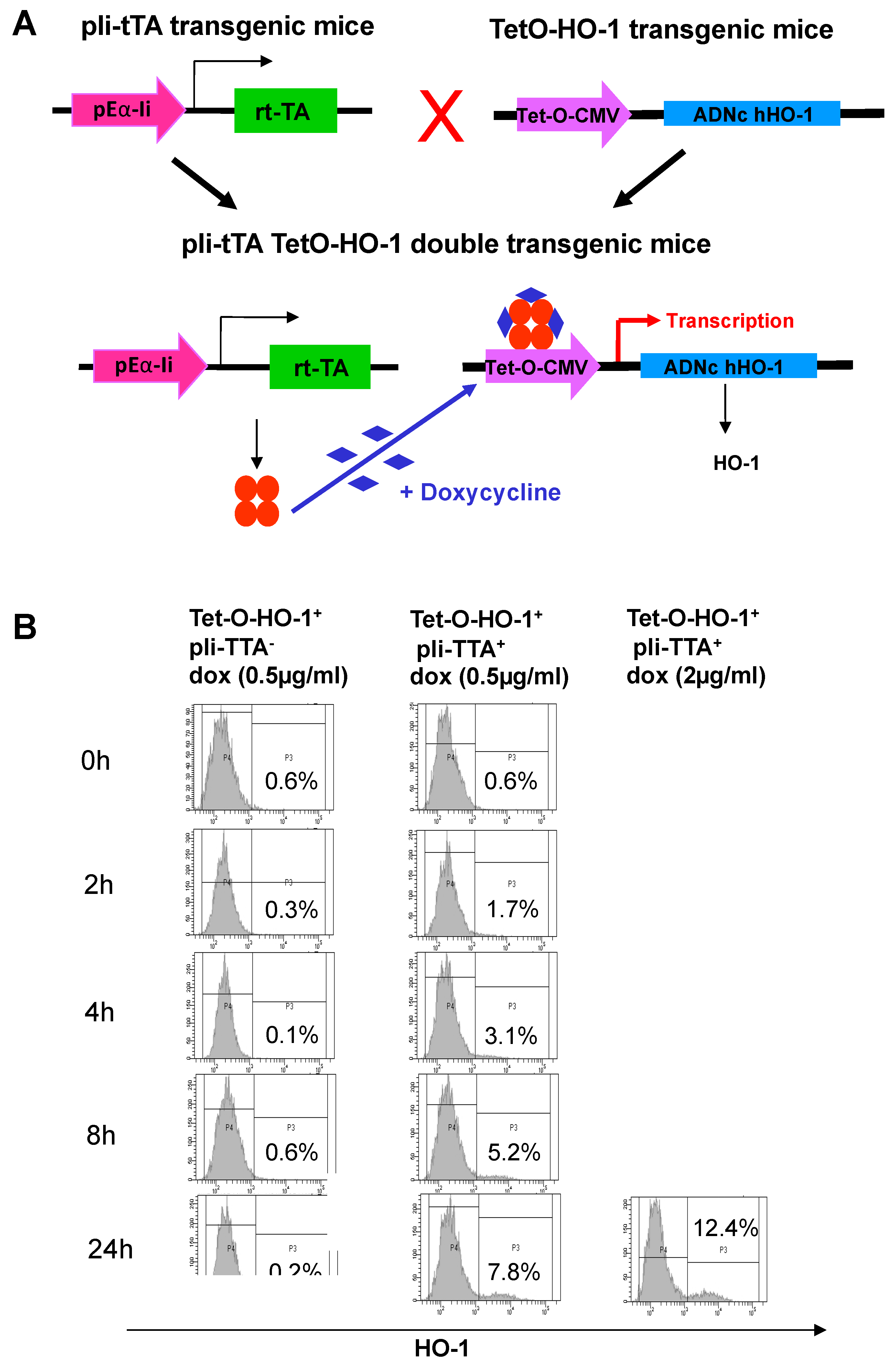
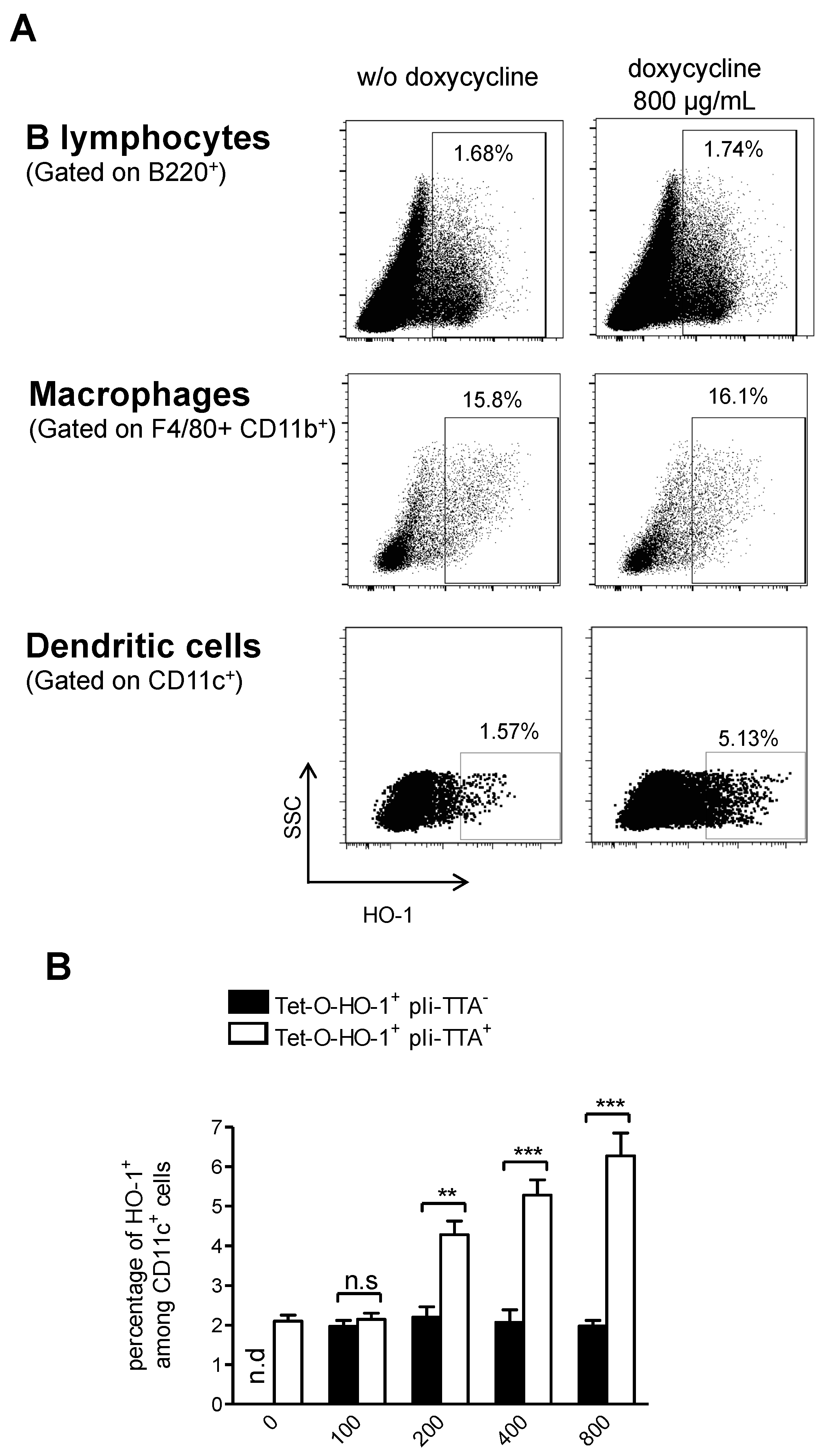
2.2. Generation of a HO-1 Inducible Transgenic Mouse Strain on the NOD Background
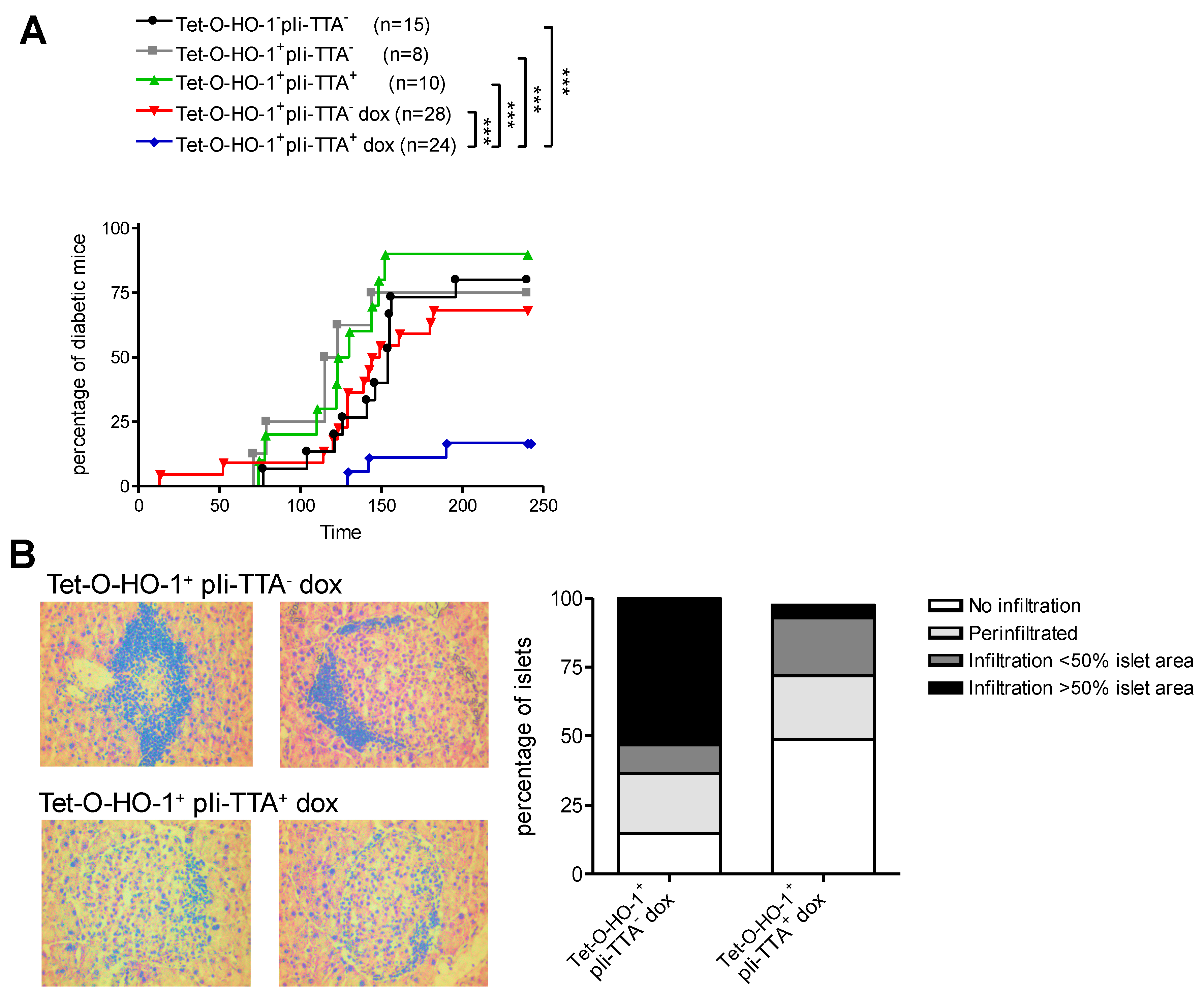
2.3. Upregulation of HO-1 in DCs Prevents T1D in NOD Mice
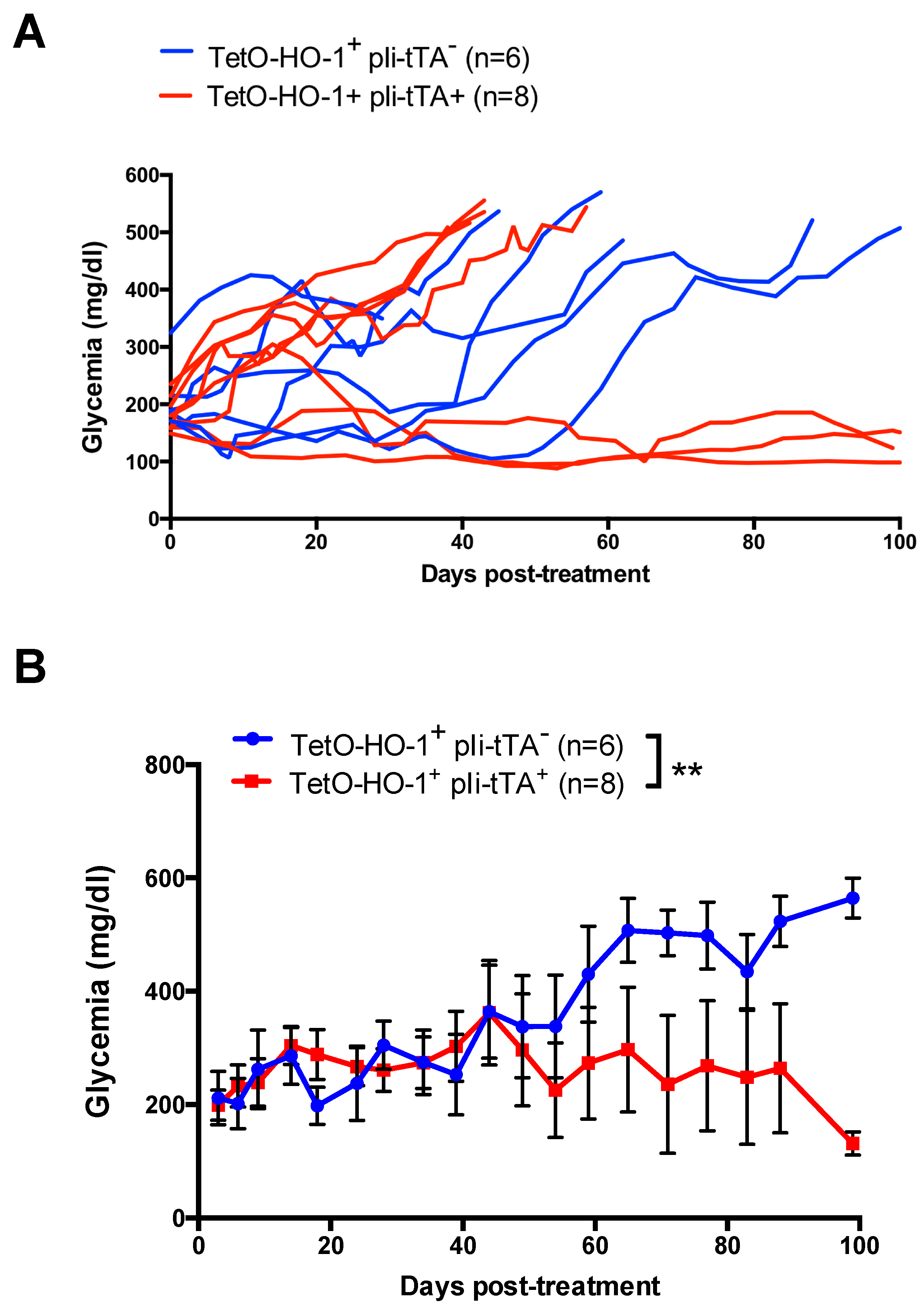
2.4. Upregulation of HO-1 in DCs Prevent A Further Increase in Glycemia in Recently Diabetic NOD Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Genotyping of rTA-HO-1
4.3. Doxycycline Treatment
4.4. Diabetes Follow-Up
4.5. Flow Cytometry
4.6. Histological Analysis.
4.7. Bone-Marrow Derived DCs
4.8. Isolation of Splenic DCs
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| APC | Antigen Presenting Cells |
| BMDC | Bone Marrow-Derived Dendritic Cells |
| CO | Carbon Monoxide |
| CoPP | Cobalt-Protoporphyrin |
| DCs | Dendritic Cells |
| DOX | Doxycycline |
| HO-1 | Heme Oxygenase 1 |
| idd | Insulin-Dependent Diabetes |
| MHC-II | Major Histocompatibility Complex II |
| NOD | Non-Obese Diabetic |
| NOR | Non-Obese Resistant |
| T1D | Type 1 Diabetes |
| Tregs | Regulatory T-cells |
| tTA | Tetracycline TransActivator |
References
- Kobayashi, M.; Yamamoto, M. Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv. Enzyme Regul. 2006, 46, 113–140. [Google Scholar] [CrossRef] [PubMed]
- Poss, K.D.; Tonegawa, S. Reduced stress defense in heme oxygenase 1-deficient cells. Proc. Natl. Acad. Sci. USA 1997, 94, 10925–10930. [Google Scholar] [CrossRef] [PubMed]
- Yachie, A.; Niida, Y.; Wada, T.; Igarashi, N.; Kaneda, H.; Toma, T.; Ohta, K.; Kasahara, Y.; Koizumi, S. Oxidative stress causes enhanced endothelial cell injury in human heme oxygenase-1 deficiency. J. Clin. Investig. 1999, 103, 129–135. [Google Scholar] [CrossRef]
- Chora, A.A.; Fontoura, P.; Cunha, A.; Pais, T.F.; Cardoso, S.; Ho, P.P.; Lee, L.Y.; Sobel, R.A.; Steinman, L.; Soares, M.P. Heme oxygenase-1 and carbon monoxide suppress autoimmune neuroinflammation. J. Clin. Investig. 2007, 117, 438–447. [Google Scholar] [CrossRef]
- Li, M.; Peterson, S.; Husney, D.; Inaba, M.; Guo, K.; Kappas, A.; Ikehara, S.; Abraham, N.G. Long-lasting expression of HO-1 delays progression of type I diabetes in NOD mice. Cell Cycle 2007, 6, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-M.; Lin, H.-H.; Chiang, M.-T.; Chang, P.-F.; Chau, L.-Y. Systemic expression of heme oxygenase-1 ameliorates type 1 diabetes in NOD mice. Diabetes 2007, 56, 1240–1247. [Google Scholar] [CrossRef]
- Xia, Z.-W.; Zhong, W.-W.; Xu, L.-Q.; Sun, J.-L.; Shen, Q.-X.; Wang, J.-G.; Shao, J.; Li, Y.-Z.; Yu, S.-C. Heme oxygenase-1-mediated CD4+CD25high regulatory T cells suppress allergic airway inflammation. J. Immunol. 2006, 177, 5936–5945. [Google Scholar] [CrossRef]
- Chauveau, C.; Bouchet, D.; Roussel, J.-C.; Mathieu, P.; Braudeau, C.; Renaudin, K.; Tesson, L.; Soulillou, J.-P.; Iyer, S.; Buelow, R.; et al. Gene transfer of heme oxygenase-1 and carbon monoxide delivery inhibit chronic rejection. Am. J. Transplant. 2002, 2, 581–592. [Google Scholar] [CrossRef]
- Braudeau, C.; Bouchet, D.; Tesson, L.; Iyer, S.; Rémy, S.; Buelow, R.; Anegon, I.; Chauveau, C. Induction of long-term cardiac allograft survival by heme oxygenase-1 gene transfer. Gene Ther. 2004, 11, 701–710. [Google Scholar] [CrossRef]
- Soares, M.P.; Lin, Y.; Anrather, J.; Csizmadia, E.; Takigami, K.; Sato, K.; Grey, S.T.; Colvin, R.B.; Choi, A.M.; Poss, K.D.; et al. Expression of heme oxygenase-1 can determine cardiac xenograft survival. Nat. Med. 1998, 4, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Hancock, W.W.; Buelow, R.; Sayegh, M.H.; Turka, L.A. Antibody-induced transplant arteriosclerosis is prevented by graft expression of anti-oxidant and anti-apoptotic genes. Nat. Med. 1998, 4, 1392–1396. [Google Scholar] [CrossRef]
- Zhao, Y.; Jia, Y.; Wang, L.; Chen, S.; Huang, X.; Xu, B.; Zhao, G.; Xiang, Y.; Yang, J.; Chen, G. Upregulation of Heme Oxygenase-1 Endues Immature Dendritic Cells With More Potent and Durable Immunoregulatory Properties and Promotes Engraftment in a Stringent Mouse Cardiac Allotransplant Model. Front. Immunol. 2018, 9, 1515. [Google Scholar] [CrossRef] [PubMed]
- Biburger, M.; Theiner, G.; Schädle, M.; Schuler, G.; Tiegs, G. Pivotal Advance: Heme oxygenase 1 expression by human CD4+ T cells is not sufficient for their development of immunoregulatory capacity. J. Leukoc. Biol. 2010, 87, 193–202. [Google Scholar] [CrossRef]
- Zelenay, S.; Chora, A.; Soares, M.P.; Demengeot, J. Heme oxygenase-1 is not required for mouse regulatory T cell development and function. Int. Immunol. 2007, 19, 11–18. [Google Scholar] [CrossRef]
- Blancou, P.; Anegon, I. Editorial: Heme oxygenase-1 and dendritic cells: What else? J. Leukoc. Biol. 2010, 87, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Rémy, S.; Blancou, P.; Tesson, L.; Tardif, V.; Brion, R.; Royer, P.J.; Motterlini, R.; Foresti, R.; Painchaut, M.; Pogu, S.; et al. Carbon monoxide inhibits TLR-induced dendritic cell immunogenicity. J. Immunol. 2009, 182, 1877–1884. [Google Scholar] [CrossRef] [PubMed]
- Simon, T.; Pogu, S.; Tardif, V.; Rigaud, K.; Rémy, S.; Piaggio, E.; Bach, J.-M.; Anegon, I.; Blancou, P. Carbon monoxide-treated dendritic cells decrease β1-integrin induction on CD8+ T cells and protect from type 1 diabetes. Eur. J. Immunol. 2013, 43, 209–218. [Google Scholar] [CrossRef]
- Moreau, A.; Hill, M.; Thébault, P.; Deschamps, J.Y.; Chiffoleau, E.; Chauveau, C.; Moullier, P.; Anegon, I.; Alliot-Licht, B.; Cuturi, M.C. Tolerogenic dendritic cells actively inhibit T cells through heme oxygenase-1 in rodents and in nonhuman primates. FASEB J. 2009, 23, 3070–3077. [Google Scholar] [CrossRef]
- Simon, T.; Pogu, J.; Rémy, S.; Brau, F.; Pogu, S.; Maquigneau, M.; Fonteneau, J.-F.; Poirier, N.; Vanhove, B.; Blancho, G.; et al. Inhibition of effector antigen-specific T cells by intradermal administration of heme oxygenase-1 inducers. J. Autoimmun. 2017, 81, 44–55. [Google Scholar] [CrossRef]
- Wong, T.-H.; Chen, H.-A.; Gau, R.-J.; Yen, J.-H.; Suen, J.-L. Heme Oxygenase-1-Expressing Dendritic Cells Promote Foxp3+ Regulatory T Cell Differentiation and Induce Less Severe Airway Inflammation in Murine Models. PLoS ONE 2016, 11, e0168919. [Google Scholar] [CrossRef]
- Mullen, Y. Development of the Nonobese Diabetic Mouse and Contribution of Animal Models for Understanding Type 1 Diabetes. Pancreas 2017, 46, 455–466. [Google Scholar] [CrossRef]
- Herold, K.C.; Vignali, D.A.A.; Cooke, A.; Bluestone, J.A. Type 1 diabetes: Translating mechanistic observations into effective clinical outcomes. Nat. Rev. Immunol. 2013, 13, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Prochazka, M.; Serreze, D.V.; Frankel, W.N.; Leiter, E.H. NOR/Lt Mice: MHC-Matched Diabetes-Resistant Control Strain for NOD Mice. Diabetes 1992, 41, 98–106. [Google Scholar] [CrossRef]
- Van Santen, H.; Benoist, C.; Mathis, D. A cassette vector for high-level reporter expression driven by a hybrid invariant chain promoter in transgenic mice. J. Immunol. Methods 2000, 245, 133–137. [Google Scholar] [CrossRef]
- Lee, T.-S.; Chau, L.-Y. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat. Med. 2002, 8, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Clayton, D.G.; Concannon, P.; Akolkar, B.; Cooper, J.D.; Erlich, H.A.; Julier, C.; Morahan, G.; Nerup, J.; Nierras, C.; et al. Genome-wide association study and meta-analysis find that over 40 loci affect risk of type 1 diabetes. Nat. Genet. 2009, 41, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Ricchetti, G.A.; Williams, L.M.; Foxwell, B.M.J. Heme oxygenase 1 expression induced by IL-10 requires STAT-3 and phosphoinositol-3 kinase and is inhibited by lipopolysaccharide. J. Leukoc. Biol. 2004, 76, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Alleva, D.G.; Pavlovich, R.P.; Grant, C.; Kaser, S.B.; Beller, D.I. Aberrant macrophage cytokine production is a conserved feature among autoimmune-prone mouse strains: Elevated interleukin (IL)-12 and an imbalance in tumor necrosis factor-alpha and IL-10 define a unique cytokine profile in macrophages from young nonobese diabetic mice. Diabetes 2000, 49, 1106–1115. [Google Scholar] [PubMed]
- Szelachowska, M.; Kretowski, A.; Kinalska, I. Decreased in vitro IL-4 [corrected] and IL-10 production by peripheral blood in first degree relatives at high risk of diabetes type-I. Horm. Metab. Res. 1998, 30, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D.J.; Poligone, B.; Bui, T.; Abdel-Motal, U.M.; Baldwin, A.S.; Tisch, R. Dendritic cells from nonobese diabetic mice exhibit a defect in NF-kappa B regulation due to a hyperactive I kappa B kinase. J. Immunol. 2001, 167, 1461–1468. [Google Scholar] [CrossRef]
- Welzen-Coppens, J.M.C.; van Helden-Meeuwsen, C.G.; Leenen, P.J.M.; Drexhage, H.A.; Versnel, M.A. Reduced numbers of dendritic cells with a tolerogenic phenotype in the prediabetic pancreas of NOD mice. J. Leukoc. Biol. 2012, 92, 1207–1213. [Google Scholar] [CrossRef]
- Marleau, A.M.; Singh, B. Myeloid dendritic cells in non-obese diabetic mice have elevated costimulatory and T helper-1-inducing abilities. J. Autoimmun. 2002, 19, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Blancou, P.; Tardif, V.; Simon, T.; Rémy, S.; Carreño, L.; Kalergis, A.; Anegon, I. Immunoregulatory properties of heme oxygenase-1. Methods Mol. Biol. 2011, 677, 247–268. [Google Scholar]
- Tardif, V.; Riquelme, S.A.; Remy, S.; Carreño, L.J.; Cortés, C.M.; Simon, T.; Hill, M.; Louvet, C.; Riedel, C.A.; Blancou, P.; et al. Carbon monoxide decreases endosome-lysosome fusion and inhibits soluble antigen presentation by dendritic cells to T cells. Eur. J. Immunol. 2013, 43, 2832–2844. [Google Scholar] [CrossRef]
- George, J.F.; Braun, A.; Brusko, T.M.; Joseph, R.; Bolisetty, S.; Wasserfall, C.H.; Atkinson, M.A.; Agarwal, A.; Kapturczak, M.H. Suppression by CD4+CD25+ regulatory T cells is dependent on expression of heme oxygenase-1 in antigen-presenting cells. Am. J. Pathol. 2008, 173, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Ndisang, J.F.; Jadhav, A. Heme oxygenase system enhances insulin sensitivity and glucose metabolism in streptozotocin-induced diabetes. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E829–E841. [Google Scholar] [CrossRef] [PubMed]
- Mabley, J.G.; Southan, G.J.; Salzman, A.L.; Szabó, C. The combined inducible nitric oxide synthase inhibitor and free radical scavenger guanidinoethyldisulfide prevents multiple low-dose streptozotocin-induced diabetes in vivo and interleukin-1beta-induced suppression of islet insulin secretion in vitro. Pancreas 2004, 28, E39–E44. [Google Scholar] [CrossRef]
- Witherden, D.; van Oers, N.; Waltzinger, C.; Weiss, A.; Benoist, C.; Mathis, D. Tetracycline-controllable selection of CD4(+) T cells: Half-life and survival signals in the absence of major histocompatibility complex class II molecules. J. Exp. Med. 2000, 191, 355–364. [Google Scholar] [CrossRef]
- Chauveau, C.; Rémy, S.; Royer, P.J.; Hill, M.; Tanguy-Royer, S.; Hubert, F.-X.; Tesson, L.; Brion, R.; Beriou, G.; Gregoire, M.; et al. Heme oxygenase-1 expression inhibits dendritic cell maturation and proinflammatory function but conserves IL-10 expression. Blood 2005, 106, 1694–1702. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pogu, J.; Tzima, S.; Kollias, G.; Anegon, I.; Blancou, P.; Simon, T. Genetic Restoration of Heme Oxygenase-1 Expression Protects from Type 1 Diabetes in NOD Mice. Int. J. Mol. Sci. 2019, 20, 1676. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071676
Pogu J, Tzima S, Kollias G, Anegon I, Blancou P, Simon T. Genetic Restoration of Heme Oxygenase-1 Expression Protects from Type 1 Diabetes in NOD Mice. International Journal of Molecular Sciences. 2019; 20(7):1676. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071676
Chicago/Turabian StylePogu, Julien, Sotiria Tzima, Georges Kollias, Ignacio Anegon, Philippe Blancou, and Thomas Simon. 2019. "Genetic Restoration of Heme Oxygenase-1 Expression Protects from Type 1 Diabetes in NOD Mice" International Journal of Molecular Sciences 20, no. 7: 1676. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071676