The Effects of Cobalt Protoporphyrin IX and Tricarbonyldichlororuthenium (II) Dimer Treatments and Its Interaction with Nitric Oxide in the Locus Coeruleus of Mice with Peripheral Inflammation

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Antinociceptive Effects of CoPP in WT, NOS1-KO, and NOS2-KO Mice
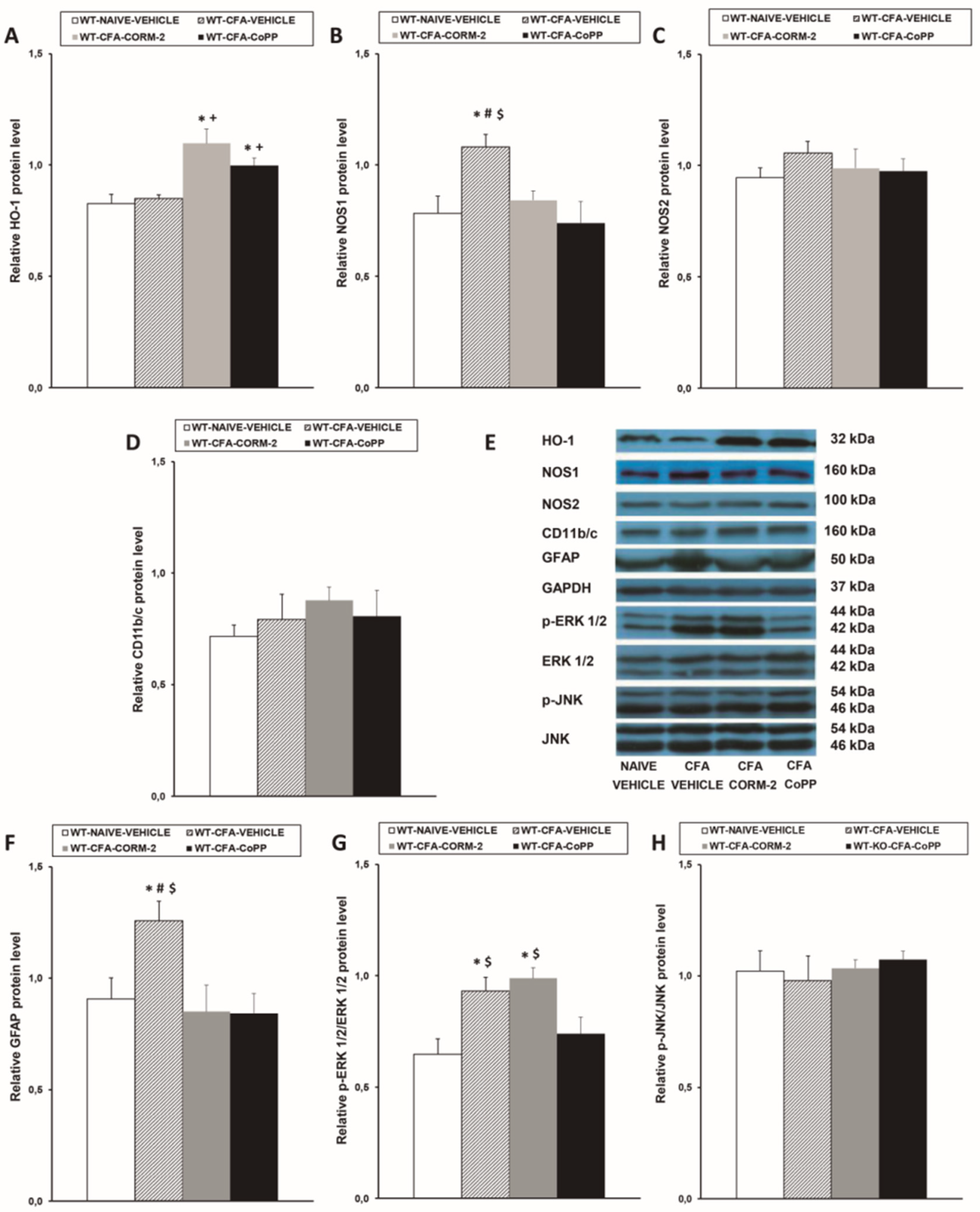
2.2. Effects of CoPP and CORM-2 on HO-1, NOS1, NOS2, CD11b/c, GFAP, p-ERK 1/2, and p-JNK Expression in the LC of WT Mice with Peripheral Inflammation
2.3. Effects of CoPP and CORM-2 on HO-1, NOS2, CD11b/c, GFAP, p-ERK 1/2, and p-JNK Expression in the LC of NOS1-KO Mice with Peripheral Inflammation
2.4. Effects of CoPP and CORM-2 on HO-1, NOS1, CD11b/c, GFAP, p-ERK 1/2, and p-JNK Expression in the LC from NOS2-KO Mice with Peripheral Inflammation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chronic Inflammatory Pain Induction
4.3. Nociceptive Behavioral Tests
4.4. Western Blot Analysis
4.5. Experimental Procedure
4.6. Drugs
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HO-1 | Heme oxygenase 1 |
| LC | Locus coeruleus |
| WT | Wild-type |
| NOS1 | Neuronal nitric oxide synthase |
| NOS2 | Inducible nitric oxide synthase |
| KO | Knockout |
| CFA | Complete Freund’s adjuvant |
| CoPP | Cobalt protoporphyrin IX |
| CORM-2 | Tricarbonyldichlororuthenium(II) dimer |
| ERK 1/2 | Extracellular signal-regulated kinase 1/2 |
| JNK | c-Jun N-terminal kinase |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GFAP | Glial fibrillary acidic protein |
| SEM | Standard error of the mean |
| ANOVA | Analysis of variance |
References
- Carvalho, P.G.; Branco, L.G.; Panissi, C.R. Involvement of the heme oxygenase-carbon monoxide-cGMP pathway in the nociception induced by acute painful stimulus in rats. Brain Res. 2011, 1385, 107–113. [Google Scholar] [CrossRef]
- Hervera, A.; Leánez, S.; Negrete, R.; Motterlini, R.; Pol, O. Carbon monoxide reduces neuropathic pain and spinal microglial activation by inhibiting nitric oxide synthesis in mice. PLoS ONE 2012, 7, e43693. [Google Scholar] [CrossRef] [PubMed]
- Bijjem, K.R.; Padi, S.S.; lal Sharma, P. Pharmacological activation of heme oxygenase (HO)-1/carbon monoxide pathway prevents the development of peripheral neuropathic pain in Wistar rats. Naunyn Schmiedebergs Arch. Pharmacol. 2013, 386, 79–90. [Google Scholar] [CrossRef]
- Castany, S.; Carcolé, M.; Leánez, S.; Pol, O. The role of carbon monoxide on the anti-nociceptive effects and expression of cannabinoid 2 receptors during painful diabetic neuropathy in mice. Psychopharmacology 2016, 233, 2209–2219. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Naito, Y.; Inoue, M.; Akagiri, S.; Mizushima, K.; Handa, O.; Kokura, S.; Ichikawa, H.; Yoshikawa, T. Inhalation of carbon monoxide ameliorates collagen-induced arthritis in mice and regulates the articular expression of IL-1beta and MCP-1. Inflammation 2009, 32, 83–88. [Google Scholar] [CrossRef]
- Bonelli, M.; Savitskaya, A.; Steiner, C.W.; Rath, E.; Bilban, M.; Wagner, O.; Bach, F.H.; Smolen, J.S.; Scheinecker, C. Heme oxygenase-1 end-products carbon monoxide and biliverdin ameliorate murine collagen induced arthritis. Clin. Exp. Rheumatol. 2012, 30, 73–78. [Google Scholar]
- Hervera, A.; Gou, G.; Leánez, S.; Pol, O. Effects of treatment with a carbon monoxide-releasing molecule and a heme oxygenase 1 inducer in the antinociceptive effects of morphine in different models of acute and chronic pain in mice. Psychopharmacology 2013, 228, 463–477. [Google Scholar] [CrossRef]
- Wang, H.; Sun, X. Carbon Monoxide-Releasing Molecule-2 Inhibits Connexin 43-Hemichannel Activity in Spinal Cord Astrocytes to Attenuate Neuropathic Pain. J. Mol. Neurosci. 2017, 63, 58–69. [Google Scholar] [CrossRef]
- Chu, Y.C.; Guan, Y.; Skinner, J.; Raja, S.N.; Johns, R.A.; Tao, Y.X. Effect of genetic knockout or pharmacologic inhibition of neuronal nitric oxide synthase on complete Freund’s adjuvant-induced persistent pain. Pain 2005, 119, 113–123. [Google Scholar] [CrossRef] [PubMed]
- De Alba, J.; Clayton, N.M.; Collins, S.D.; Colthup, P.; Chessell, I.; Knowles, R.G. GW274150, a novel and highly selective inhibitor of the inducible isoform of nitric oxide synthase (iNOS), shows analgesic effects in rat models of inflammatory and neuropathic pain. Pain 2006, 120, 170–181. [Google Scholar] [CrossRef]
- Boettger, M.K.; Uceyler, N.; Zelenka, M.; Schmitt, A.; Reif, A.; Chen, Y.; Sommer, C. Differences in inflammatory pain in nNOS-, iNOS- and eNOS-deficient mice. Eur. J. Pain 2007, 11, 810–818. [Google Scholar] [CrossRef]
- Leánez, S.; Hervera, A.; Pol, O. Peripheral antinociceptive effects of mu- and delta-opioid receptor agonists in NOS2 and NOS1 knockout mice during chronic inflammatory pain. Eur. J. Pharmacol. 2009, 602, 41–49. [Google Scholar] [CrossRef]
- Hervera, A.; Negrete, R.; Leánez, S.; Martín-Campos, J.; Pol, O. The role of nitric oxide in the local antiallodynic and antihyperalgesic effects and expression of delta-opioid and cannabinoid-2 receptors during neuropathic pain in mice. J. Pharmacol. Exp. Ther. 2010, 334, 887–896. [Google Scholar] [CrossRef]
- Durante, W.; Schafer, A.I. Carbon monoxide and vascular cell function (review). Int. J. Mol. Med. 1998, 2, 255–262. [Google Scholar] [CrossRef]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Ferrone, A.; Patruno, A.; Riccioni, G.; De Lutiis, M.A.; Felaco, M.; Grilli, A. Negative feedback interaction of HO-1/INOS in PBMC of acute congestive heart failure patients. J. Biol. Regul. Homeost. Agents 2013, 27, 739–748. [Google Scholar]
- Luo, W.; Wang, Y.; Yang, H.; Dai, C.; Hong, H.; Li, J.; Liu, Z.; Guo, Z.; Chen, X.; He, P.; et al. Heme oxygenase-1 ameliorates oxidative stress-induced endothelial senescence via regulating endothelial nitric oxide synthase activation and coupling. Aging 2018, 10, 1722–1744. [Google Scholar] [CrossRef]
- Soriano, R.N.; Kwiatkoski, M.; Batalhao, M.E.; Branco, L.G.; Carnio, E.C. Interaction between the carbon monoxide and nitric oxide pathways in the locus coeruleus during fever. Neuroscience 2012, 206, 69–80. [Google Scholar] [CrossRef]
- Duvigneau, J.C.; Kozlov, A.V. Pathological Impact of the Interaction of NO and CO with Mitochondria in Critical Care Diseases. Front. Med. 2017, 4, 223. [Google Scholar] [CrossRef]
- Steiner, A.A.; Branco, L.G.; Cunha, F.Q.; Ferreira, S.H. Role of the haeme oxygenase/carbon monoxide pathway in mechanical nociceptor hypersensitivity. Br. J. Pharmacol. 2001, 132, 1673–1682. [Google Scholar] [CrossRef]
- Negrete, R.; Hervera, A.; Leánez, S.; Pol, O. Treatment with a carbon monoxide-releasing molecule inhibits chronic inflammatory pain in mice: Nitric oxide contribution. Psychopharmacology 2014, 231, 853–861. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Cheng, Z.; Zhang, J.; Xu, S.; Liu, H.; Jia, H.; Jin, Y. Spinal Heme Oxygenase-1 (HO-1) Exerts Antinociceptive Effects Against Neuropathic Pain in a Mouse Model of L5 Spinal Nerve Ligation. Pain Med. 2016, 17, 220–229. [Google Scholar] [CrossRef]
- Riego, G.; Redondo, A.; Leánez, S.; Pol, O. Mechanism implicated in the antiallodynic and antihyperalgesic effects induced by the activation of heme oxygenase 1/carbon monoxide signaling pathway in the central nervous system of mice with neuropathic pain. Biochem. Pharmacol. 2018, 148, 52–63. [Google Scholar] [CrossRef]
- Benarroch, E.E. Locus coeruleus. Cell. Tissue Res. 2018, 373, 221–232. [Google Scholar] [CrossRef]
- Hundahl, C.A.; Kelsen, J.; Dewilde, S.; Hay-Schmidt, A. Neuroglobin in the rat brain (II): Co-localisation with neurotransmitters. Neuroendocrinology 2008, 88, 183–198. [Google Scholar] [CrossRef]
- Cazuza, R.A.; Pol, O.; Leite-Panissi, C.R.A. Enhanced expression of heme oxygenase-1 in the locus coeruleus can be associated with anxiolytic-like effects. Behav. Brain Res. 2018, 336, 204–210. [Google Scholar] [CrossRef]
- Le Maître, E.; Barde, S.S.; Palkovits, M.; Diaz-Heijtz, R.; Hökfelt, T.G. Distinct features of neurotransmitter systems in the human brain with focus on the galanin system in locus coeruleus and dorsal raphe. Proc. Natl. Acad. Sci. USA 2013, 110, E536–E545. [Google Scholar]
- Pablos, P.; Mendiguren, A.; Pineda, J. Contribution of nitric oxide-dependent guanylate cyclase and reactive oxygen species signaling pathways to desensitization of μ-opioid receptors in the rat locus coeruleus. Neuropharmacology 2015, 99, 422–431. [Google Scholar] [CrossRef]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef]
- Ji, R.R.; Gereau, R.W., 4th; Malcangio, M.; Strichartz, G.R. MAP kinase and pain. Brain Res. Rev. 2009, 60, 135–148. [Google Scholar] [CrossRef]
- Redondo, A.; Chamorro, P.A.F.; Riego, G.; Leánez, S.; Pol, O. Treatment with Sulforaphane Produces Antinociception and Improves Morphine Effects during Inflammatory Pain in Mice. J. Pharmacol. Exp. Ther. 2017, 363, 293–302. [Google Scholar] [CrossRef]
- Imbe, H.; Okamoto, K.; Donishi, T.; Kawai, S.; Enoki, K.; Senba, E.; Kimura, A. Activation of ERK in the locus coeruleus following acute noxious stimulation. Brain Res. 2009, 1263, 50–57. [Google Scholar] [CrossRef]
- Borges, G.; Neto, F.; Mico, J.A.; Berrocoso, E. Reversal of monoarthritis-induced affective disorders by diclofenac in rats. Anesthesiology 2014, 120, 1476–1490. [Google Scholar] [CrossRef]
- Gou, G.; Leánez, S.; Pol, O. The role of gaseous neurotransmitters in the antinociceptive effects of morphine during acute thermal pain. Eur. J. Pharmacol. 2014, 737, 41–46. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, Z.J.; Zhu, M.D.; Jiang, B.C.; Yang, T.; Gao, Y.J. Exogenous induction of HO-1 alleviates vincristine-induced neuropathic pain by reducing spinal glial activation in mice. Neurobiol. Dis. 2015, 79, 100–110. [Google Scholar] [CrossRef]
- McDonnell, C.; Leánez, S.; Pol, O. The inhibitory effects of cobalt protoporphyrin IX and cannabinoid 2 receptor agonists in type 2 diabetic mice. Int. J. Mol. Sci. 2017, 18, 2268. [Google Scholar] [CrossRef]
- Carcolé, M.; Castany, S.; Leánez, S.; Pol, O. Treatment with a heme oxygenase 1 inducer enhances the antinociceptive effects of µ-opioid, δ-opioid, and cannabinoid 2 receptors during inflammatory pain. J. Pharmacol. Exp. Ther. 2014, 351, 224–232. [Google Scholar] [CrossRef]
- Infante, C.; Díaz, M.; Hernández, A.; Constandil, L.; Pelissier, T. Expression of nitric oxide synthase isoforms in the dorsal horn of monoarthritic rats: Effect of competitive and uncompetitive N-methyl-d-aspartate antagonists. Arthritis Res. Ther. 2007, 9, R53. [Google Scholar] [CrossRef]
- Negrete, R.; Hervera, A.; Leánez, S.; Martín-Campos, J.M.; Pol, O. The antinociceptive effects of JWH-015 in chronic inflammatory pain are produced by nitric oxide-cGMP-PKG-KATP pathway activation mediated by opioids. PLoS ONE 2011, 6, e26688. [Google Scholar] [CrossRef]
- Raghavendra, V.; Tanga, F.Y.; DeLeo, J.A. Complete Freunds adjuvant-induced peripheral inflammation evokes glial activation and proinflammatory cytokine expression in the CNS. Eur. J. Neurosci. 2004, 20, 467–473. [Google Scholar] [CrossRef]
- Mika, J.; Osikowicz, M.; Rojewska, E.; Korostynski, M.; Wawrzczak-Bargiela, A.; Przewlocki, R.; Przewlocka, B. Differential activation of spinal microglial and astroglial cells in a mouse model of peripheral neuropathic pain. Eur. J. Pharmacol. 2009, 623, 65–72. [Google Scholar] [CrossRef]
- Ikeda, H.; Mochizuki, K.; Murase, K. Astrocytes are involved in long-term facilitation of neuronal excitation in the anterior cingulate cortex of mice with inflammatory pain. Pain 2013, 154, 2836–2843. [Google Scholar] [CrossRef]
- Xu, Y.; Cheng, G.; Zhu, Y.; Zhang, X.; Pu, S.; Wu, J.; Lv, Y.; Du, D. Anti-nociceptive roles of the glia-specific metabolic inhibitor fluorocitrate in paclitaxel-evoked neuropathic pain. Acta Biochim. Biophys. Sin. 2016, 48, 902–908. [Google Scholar] [CrossRef]
- Edelmayer, R.M.; Brederson, J.D.; Jarvis, M.F.; Bitner, R.S. Biochemical and pharmacological assessment of MAP-kinase signaling along pain pathways in experimental rodent models: A potential tool for the discovery of novel antinociceptive therapeutics. Biochem. Pharmacol. 2014, 87, 390–398. [Google Scholar] [CrossRef]
- Borges, G.; Miguelez, C.; Neto, F.; Mico, J.A.; Ugedo, L.; Berrocoso, E. Activation of Extracellular Signal-Regulated Kinases (ERK 1/2) in the Locus Coeruleus Contributes to Pain-Related Anxiety in Arthritic Male Rats. Int. J. Neuropsychopharmacol. 2017, 20, 463. [Google Scholar] [CrossRef]
- Jurga, A.M.; Piotrowska, A.; Makuch, W.; Przewlocka, B.; Mika, J. Blockade of P2X4 receptors inhibits neuropathic pain-related behavior by preventing MMP-9 activation and consequently, pronociceptive interleukin release in a rat model. Front. Pharmacol. 2017, 8, 48. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Hargreaves, K.; Dubner, R.; Brown, F.; Flores, C.; Joris, J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain 1988, 32, 77–88. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, P.; Cazuza, R.A.; Mendes-Gomes, J.; Díaz, A.F.; Polo, S.; Leánez, S.; Leite-Panissi, C.R.A.; Pol, O. The Effects of Cobalt Protoporphyrin IX and Tricarbonyldichlororuthenium (II) Dimer Treatments and Its Interaction with Nitric Oxide in the Locus Coeruleus of Mice with Peripheral Inflammation. Int. J. Mol. Sci. 2019, 20, 2211. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092211
Moreno P, Cazuza RA, Mendes-Gomes J, Díaz AF, Polo S, Leánez S, Leite-Panissi CRA, Pol O. The Effects of Cobalt Protoporphyrin IX and Tricarbonyldichlororuthenium (II) Dimer Treatments and Its Interaction with Nitric Oxide in the Locus Coeruleus of Mice with Peripheral Inflammation. International Journal of Molecular Sciences. 2019; 20(9):2211. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092211
Chicago/Turabian StyleMoreno, Patricia, Rafael Alves Cazuza, Joyce Mendes-Gomes, Andrés Felipe Díaz, Sara Polo, Sergi Leánez, Christie Ramos Andrade Leite-Panissi, and Olga Pol. 2019. "The Effects of Cobalt Protoporphyrin IX and Tricarbonyldichlororuthenium (II) Dimer Treatments and Its Interaction with Nitric Oxide in the Locus Coeruleus of Mice with Peripheral Inflammation" International Journal of Molecular Sciences 20, no. 9: 2211. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092211