Phenotypic and Genotypic Characterization of West Nile Virus Isolate 2004Hou3

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Sequencing
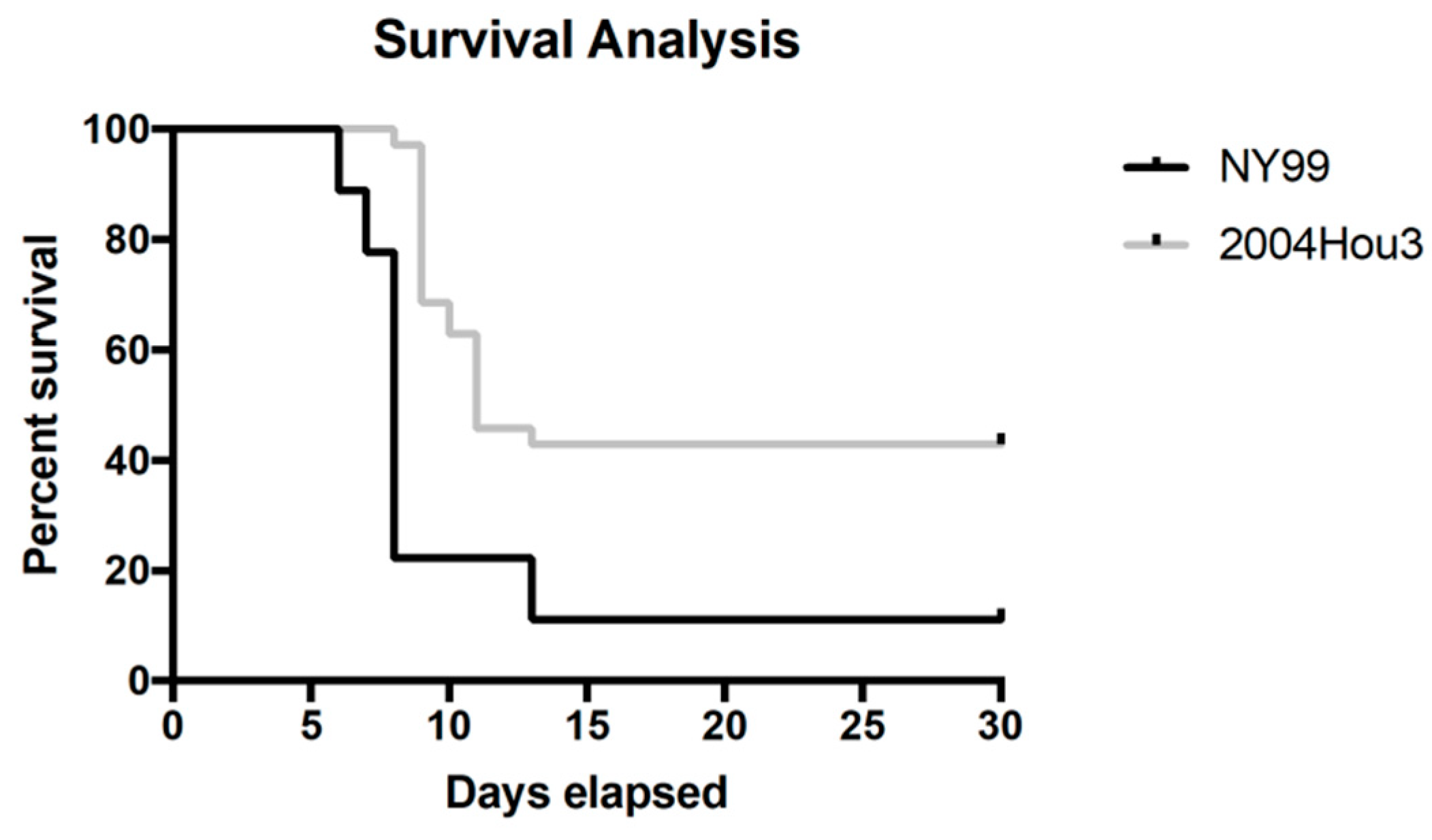
2.2. Pathogenesis in Mice
2.3. Viral Persistance
3. Discussion
4. Materials and Methods
4.1. Cells and Viruses
4.2. Sequencing
4.3. Animals
4.4. Viral RNA Isolation and PCR of Animal Tissue
4.5. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DPI | Days post infection |
| MPI | Months post infection |
| PFU | Plaque forming units |
| SNP | Single nucleotide polymorhpisms |
| UTR | Untranslated region |
| WNV | West Nile virus |
| WNF | West Nile fever |
| WNND | West Nile neuroinvasive disease |
References
- Ronca, S.E.; Murray, K.O.; Nolan, M.S. Cumulative Incidence of West Nile Virus Infection, Continental United States, 1999–2016. Emerg. Infect. Dis. 2019, 25, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Weatherhead, J.E.; Miller, V.E.; Garcia, M.N.; Hasbun, R.; Salazar, L.; Dimachkie, M.M.; Murray, K.O. Long-term neurological outcomes in West Nile virus-infected patients: An observational study. Am. J. Trop. Med. Hyg. 2015, 92, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Nolan, M.S.; Podoll, A.S.; Hause, A.M.; Akers, K.M.; Finkel, K.W.; Murray, K.O. Prevalence of chronic kidney disease and progression of disease over time among patients enrolled in the Houston West Nile virus cohort. PLoS ONE 2012, 7, e40374. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.O.; Kolodziej, S.; Ronca, S.E.; Gorchakov, R.; Navarro, P.; Nolan, M.S.; Podoll, A.; Finkel, K.; Mandayam, S. Visualization of West Nile Virus in Urine Sediment using Electron Microscopy and Immunogold up to Nine Years Postinfection. Am. J. Trop. Med. Hyg. 2017, 97, 1913–1919. [Google Scholar] [CrossRef] [PubMed]
- Borisevich, V.; Seregin, A.; Nistler, R.; Mutabazi, D.; Yamshchikov, V. Biological properties of chimeric West Nile viruses. Virology 2006, 349, 371–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beasley, D.W.; Li, L.; Suderman, M.T.; Barrett, A.D. Mouse neuroinvasive phenotype of West Nile virus strains varies depending upon virus genotype. Virology 2002, 296, 17–23. [Google Scholar] [CrossRef] [PubMed]
- McMullen, A.R.; May, F.J.; Li, L.; Guzman, H.; Bueno, R., Jr.; Dennett, J.A.; Tesh, R.B.; Barrett, A.D. Evolution of new genotype of West Nile virus in North America. Emerg. Infect. Dis. 2011, 17, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Lu, L.; Guzman, H.; Tesh, R.B.; Xiao, S.Y. Persistent infection and associated nucleotide changes of West Nile virus serially passaged in hamsters. J. Gen. Virol. 2008, 89, 3073–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, V.; Welte, T.; Bao, X.; Xie, G.; Wang, J.; Higgs, S.; Tesh, R.B.; Wang, T. A hamster-derived West Nile virus strain is highly attenuated and induces a differential proinflammatory cytokine response in two murine cell lines. Virus Res. 2012, 167, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Saxena, V.; Xie, G.; Li, B.; Farris, T.; Welte, T.; Gong, B.; Boor, P.; Wu, P.; Tang, S.J.; Tesh, R.; et al. A hamster-derived West Nile virus isolate induces persistent renal infection in mice. PLoS Negl. Trop. Dis. 2013, 7, e2275. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.T.; Ebel, G.D.; Lanciotti, R.S.; Brault, A.C.; Guzman, H.; Siirin, M.; Lambert, A.; Parsons, R.E.; Beasley, D.W.; Novak, R.J.; et al. Phylogenetic analysis of North American West Nile virus isolates, 2001-2004: Evidence for the emergence of a dominant genotype. Virology 2005, 342, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Mann, B.R.; McMullen, A.R.; Swetnam, D.M.; Salvato, V.; Reyna, M.; Guzman, H.; Bueno, R., Jr.; Dennett, J.A.; Tesh, R.B.; Barrett, A.D. Continued evolution of West Nile virus, Houston, Texas, USA, 2002–2012. Emerg. Infect. Dis. 2013, 19, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Morrey, J.D.; Siddharthan, V.; Wang, H.; Hall, J.O.; Skirpstunas, R.T.; Olsen, A.L.; Nordstrom, J.L.; Koenig, S.; Johnson, S.; Diamond, M.S. West Nile virus-induced acute flaccid paralysis is prevented by monoclonal antibody treatment when administered after infection of spinal cord neurons. J. Neurovirol. 2008, 14, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.A.; Wang, H.; Siddharthan, V.; Morrey, J.D.; Diamond, M.S. Axonal transport mediates West Nile virus entry into the central nervous system and induces acute flaccid paralysis. Proc. Natl. Acad. Sci. USA 2007, 104, 17140–17145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, J.B.; Swarts, J.L.; Wilkins, C.; Thomas, S.; Green, R.; Sekine, A.; Voss, K.M.; Ireton, R.C.; Mooney, M.; Choonoo, G.; et al. A Mouse Model of Chronic West Nile Virus Disease. PLoS Pathog. 2016, 12, e1005996. [Google Scholar] [CrossRef] [PubMed]
- Vasek, M.J.; Garber, C.; Dorsey, D.; Durrant, D.M.; Bollman, B.; Soung, A.; Yu, J.; Perez-Torres, C.; Frouin, A.; Wilton, D.K.; et al. A complement-microglial axis drives synapse loss during virus-induced memory impairment. Nature 2016, 534, 538–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piorkowski, G.; Baronti, C.; de Lamballerie, X.; de Fabritus, L.; Bichaud, L.; Pastorino, B.A.; Bessaud, M. Development of generic Taqman PCR and RT-PCR assays for the detection of DNA and mRNA of beta-actin-encoding sequences in a wide range of animal species. J. Virol. Methods 2014, 202, 101–105. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Nucleotide position | NY-99 nucleotide (amino acid) | 2004Hou3 nucleotide (amino acid) | Genome region |
|---|---|---|---|
| 660 | C (I) | U (I) | prM |
| 774 | U (T) | C (T) | prM |
| 1244 | G (R) | A (K) | E |
| 1442 | U (V) | C (A) | E |
| 1465 | C (L) | U (F) | E |
| 1797 | C (N) | U (N) | E |
| 1815 | G (S) | A (S) | E |
| 1997 | C (A) | U (V) | E |
| 2049 | U (F) | C (F) | E |
| 2253 | A (G) | G (G) | E |
| 2428 | G (V) | U (F) | E |
| 2466 | C (H) | U (H) | E |
| 4107 | C (A) | U (A) | NS2A |
| 4146 | A (L) | G (L) | NS2A |
| 4803 | C (R) | U (R) | NS3 |
| 5275 | C (L) | U (L) | NS3 |
| 5280 | G (A) | A (A) | NS3 |
| 5736 | C (C) | U (C) | NS3 |
| 6045 | C (H) | U (H) | NS3 |
| 6138 | C (F) | U (F) | NS3 |
| 6238 | C (L) | U (L) | NS3 |
| 6405 | U (D) | C (D) | NS3 |
| 6426 | C (H) | U (H) | NS3 |
| 6589 | A (R) | C (R) | NS4A |
| 6726 | C (V) | U (V) | NS4A |
| 6996 | C (F) | U (F) | NS4B |
| 7416 | C (P) | U (P) | NS4B |
| 7938 | U (G) | C (G) | NS5 |
| 7977 | A (E) | G (E) | NS5 |
| 8199 | C (H) | U (H) | NS5 |
| 8235 | U (C) | C (C) | NS5 |
| 8325 | C (S) | U (S) | NS5 |
| 8520 | G (K) | A (K) | NS5 |
| 8727 | U (T) | C (T) | NS5 |
| 8835 | G (A) | A (A) | NS5 |
| 8970 | U (D) | C (D) | NS5 |
| 9013 | C (L) | U (L) | NS5 |
| 9144 | C (F) | U (F) | NS5 |
| 9352 | C (L) | U (L) | NS5 |
| 10,338 | C (D) | U (D) | NS5 |
| 10,440 | A | G | 3’UTR |
| 10,496 | T | C | 3’UTR |
| 10,703 | G | A | 3’UTR |
| Days Post Infection | Brain | Lung | Heart | Spleen | Liver | Kidney | Bladder |
|---|---|---|---|---|---|---|---|
| 47 | 4/6 | 0/6 | 0/6 | 2/6 | 1/6 | 2/6 | NA |
| 180 | 1/8 | 0/3 | 0/3 | 1/8 | 0/8 | 1/8 | 0/8 |
| Days Post Infection | Whole Blood | Urine |
|---|---|---|
| 47 | 5/6 | 1/6 |
| 60 | 3/6 | 0/6 |
| 90 | 3/6 | 0/6 |
| 180 | 2/6 | 0/6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronca, S.E.; Gorchakov, R.; Berry, R.; Alvarado, R.E.; Gunter, S.M.; Murray, K.O. Phenotypic and Genotypic Characterization of West Nile Virus Isolate 2004Hou3. Int. J. Mol. Sci. 2019, 20, 1936. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20081936
Ronca SE, Gorchakov R, Berry R, Alvarado RE, Gunter SM, Murray KO. Phenotypic and Genotypic Characterization of West Nile Virus Isolate 2004Hou3. International Journal of Molecular Sciences. 2019; 20(8):1936. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20081936
Chicago/Turabian StyleRonca, Shannon E., Rodion Gorchakov, Rebecca Berry, R. Elias Alvarado, Sarah M. Gunter, and Kristy O. Murray. 2019. "Phenotypic and Genotypic Characterization of West Nile Virus Isolate 2004Hou3" International Journal of Molecular Sciences 20, no. 8: 1936. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20081936