Differential Subcellular Distribution and Translocation of Seven 14-3-3 Isoforms in Response to EGF and During the Cell Cycle


Abstract
:1. Introduction
2. Results
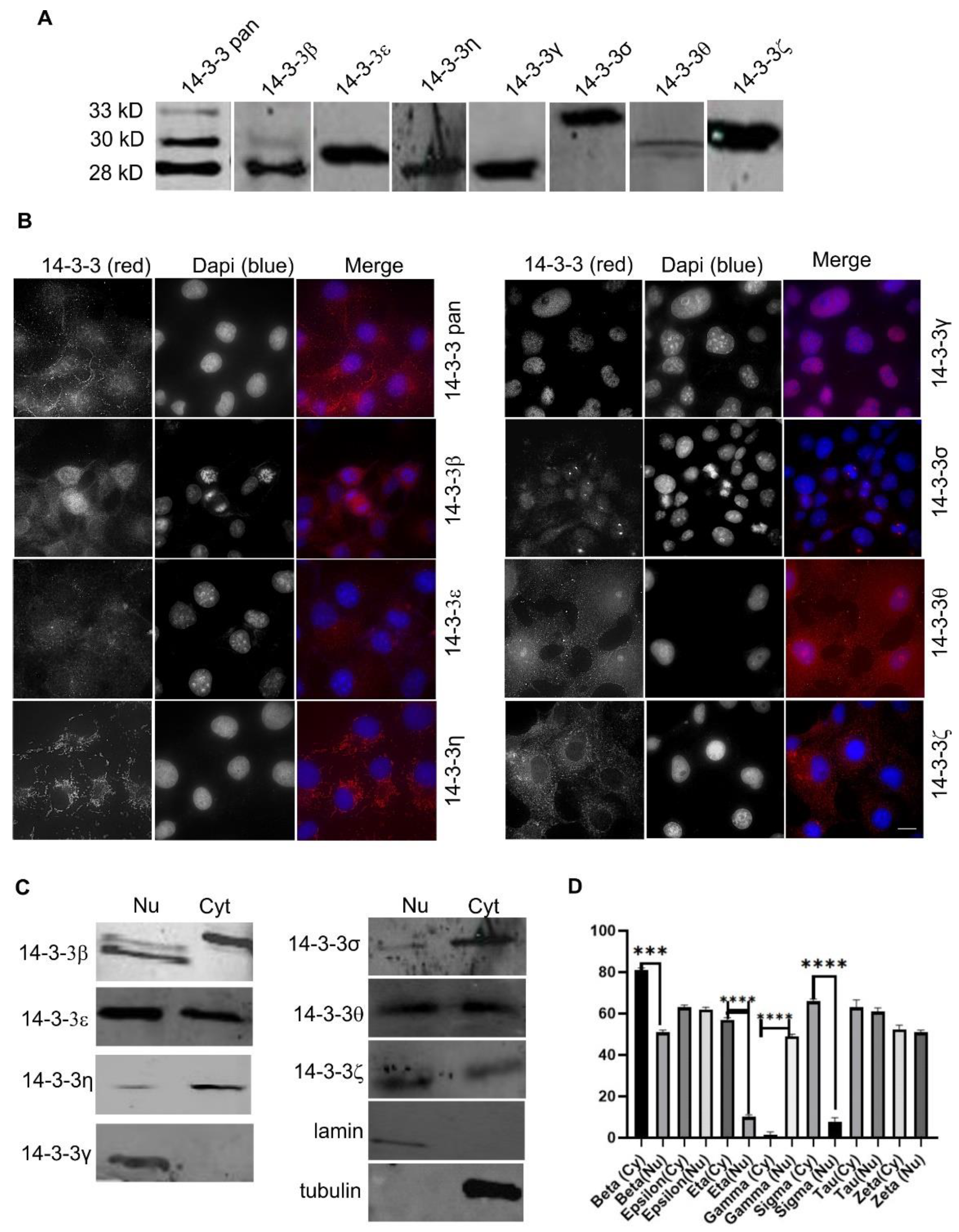
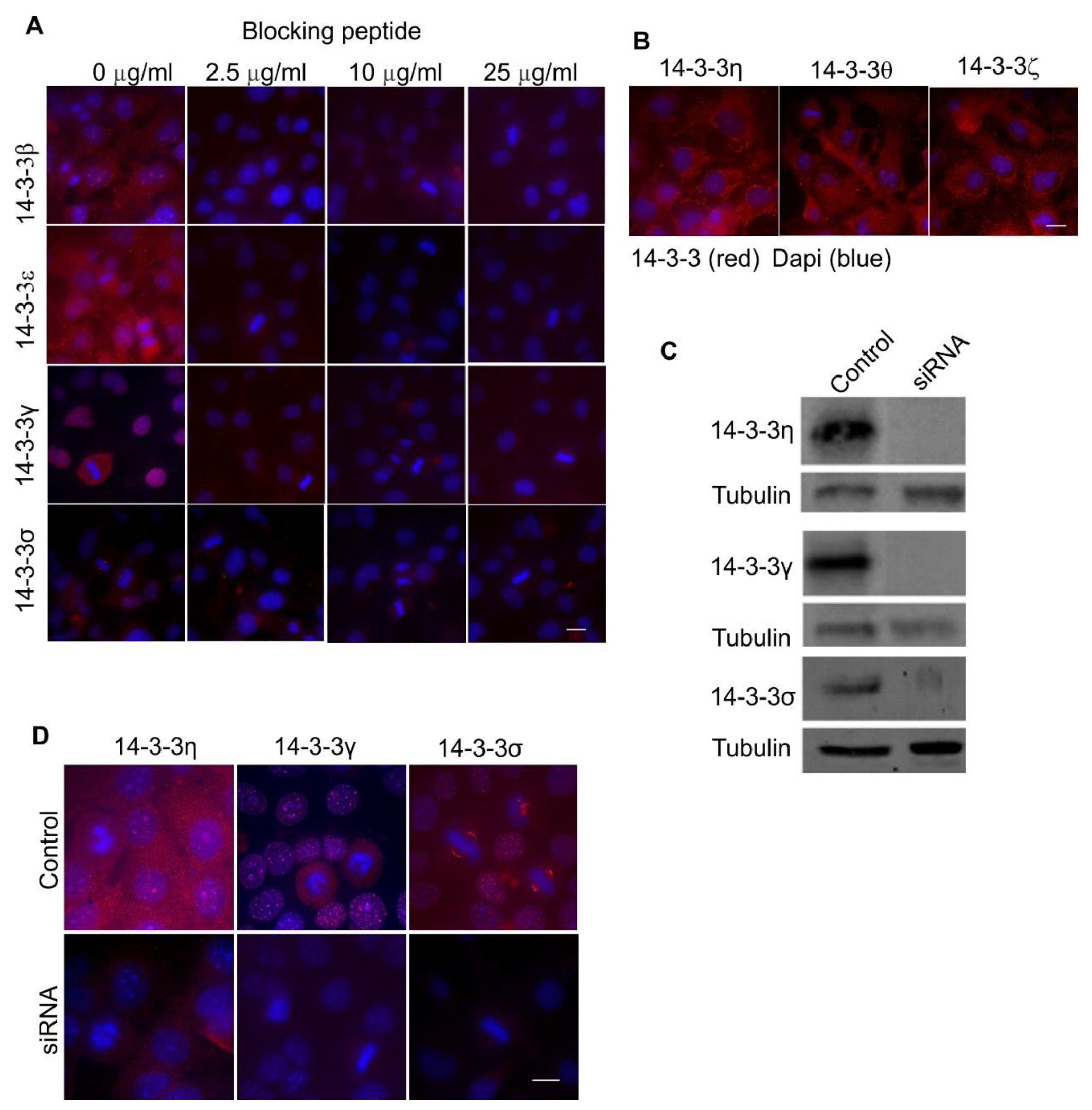
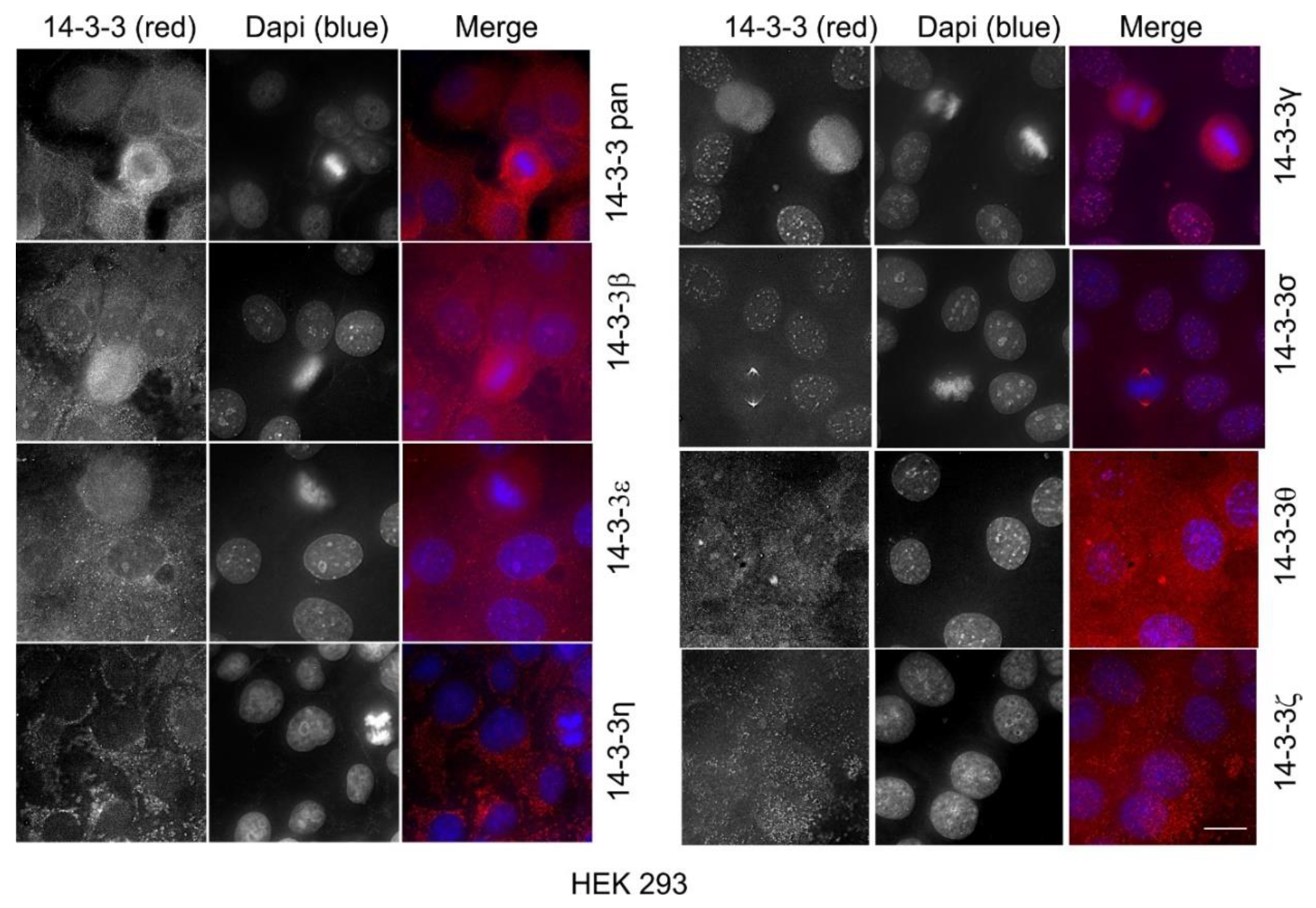
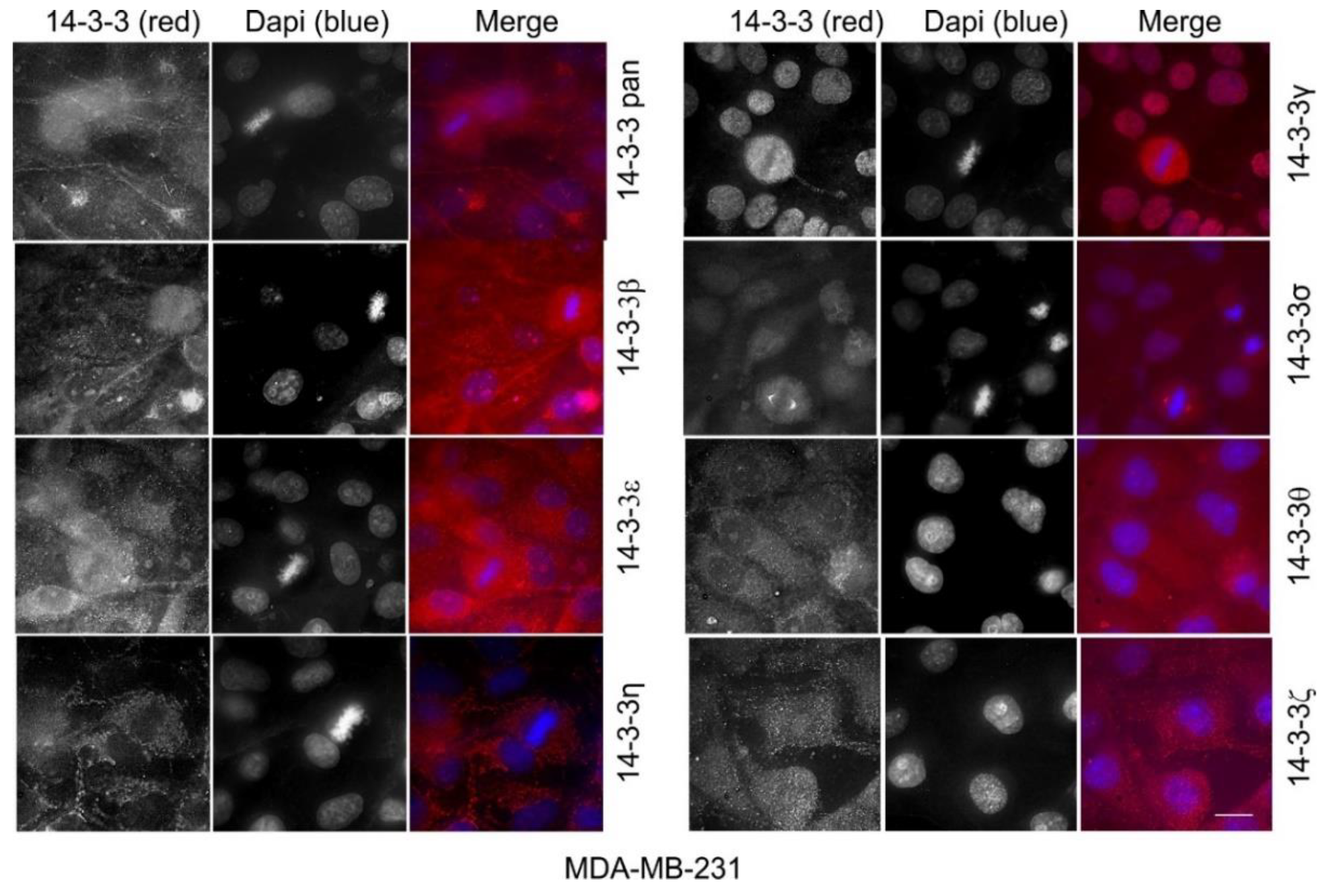
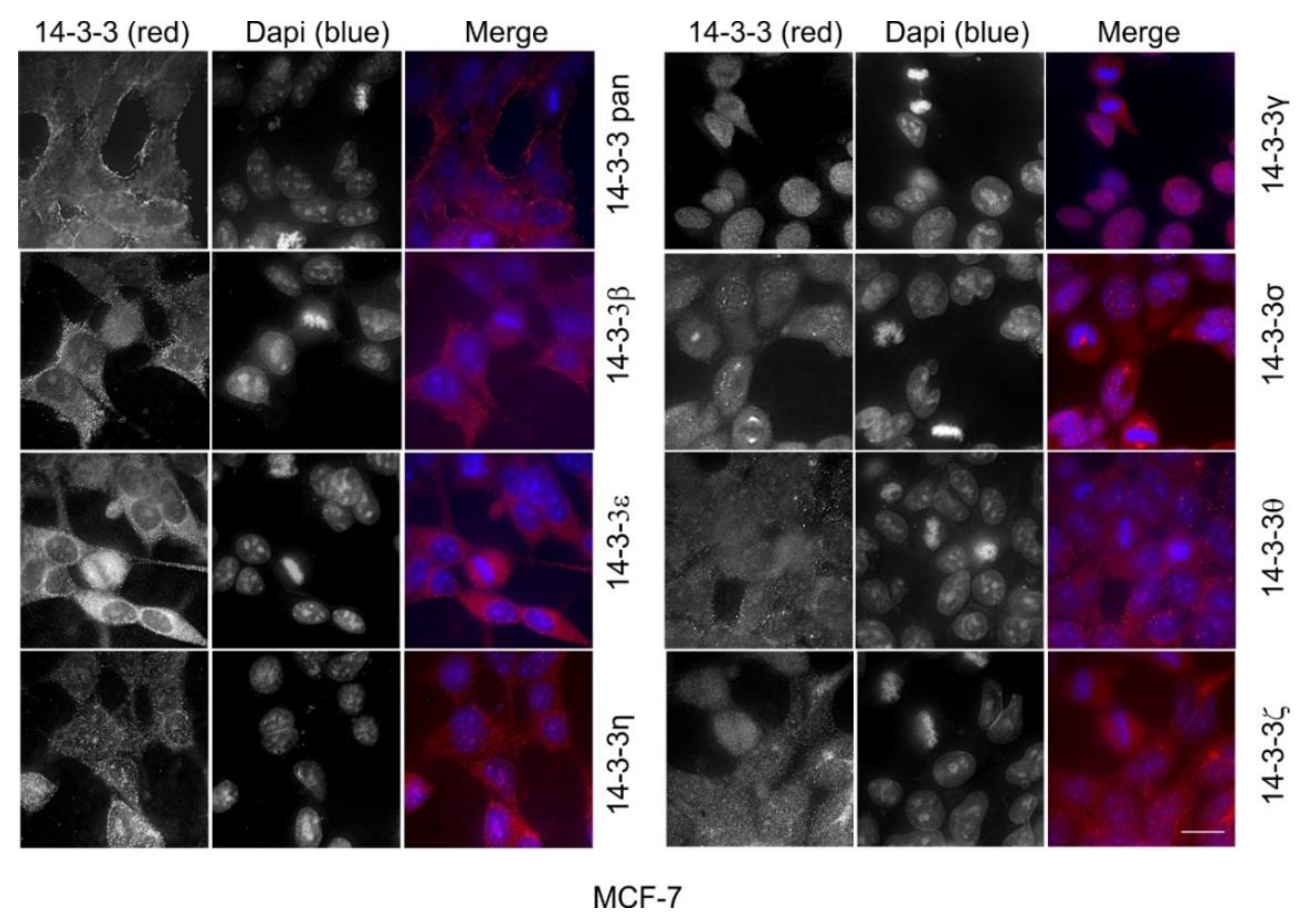
2.1. The Expression and Subcellular Localization of Total and Each Isoform of 14-3-3 Proteins
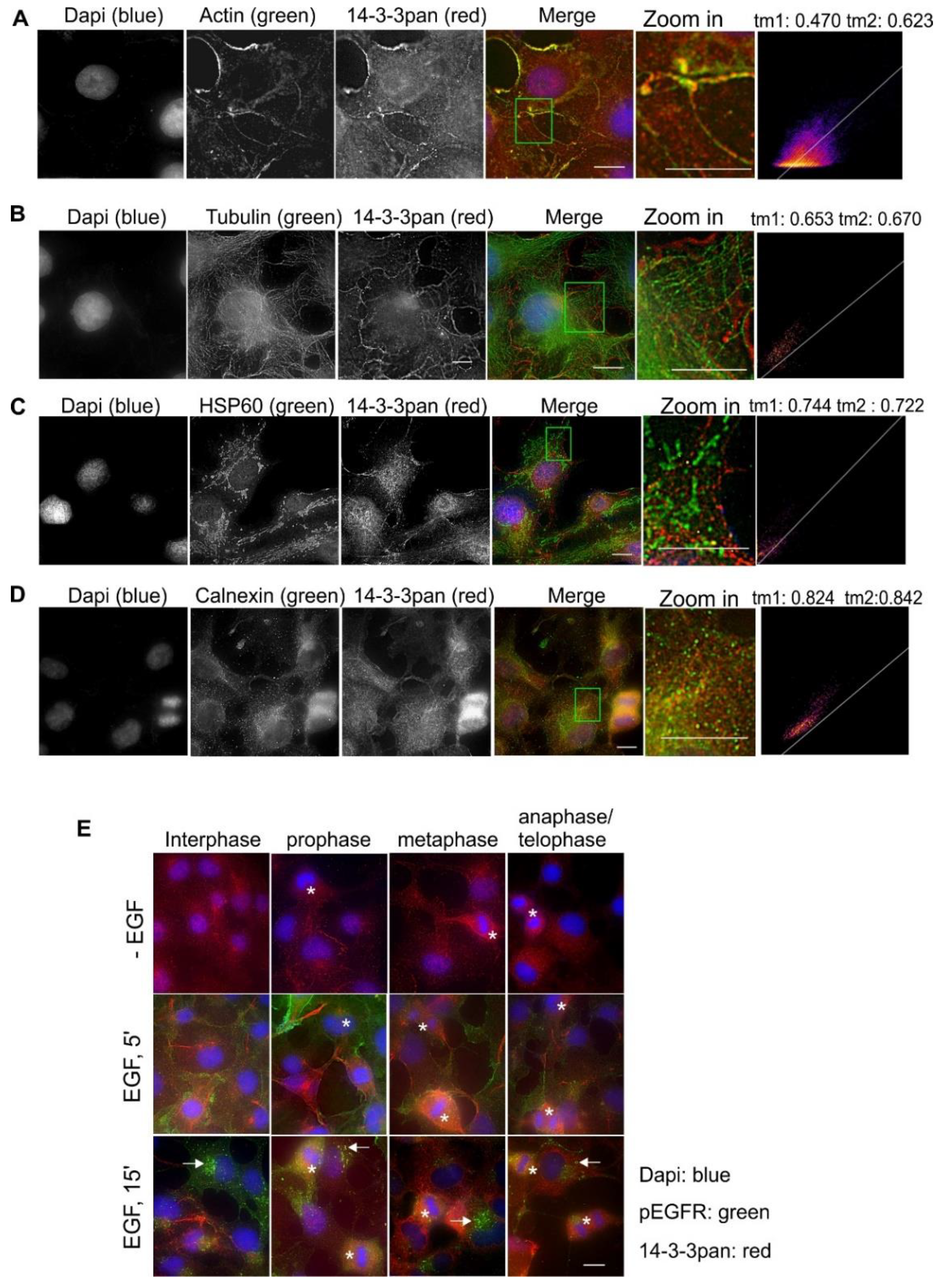
2.2. Subcellular Localization and Translocation of Pan 14-3-3 in Response to EGF and During the Cell Cycle
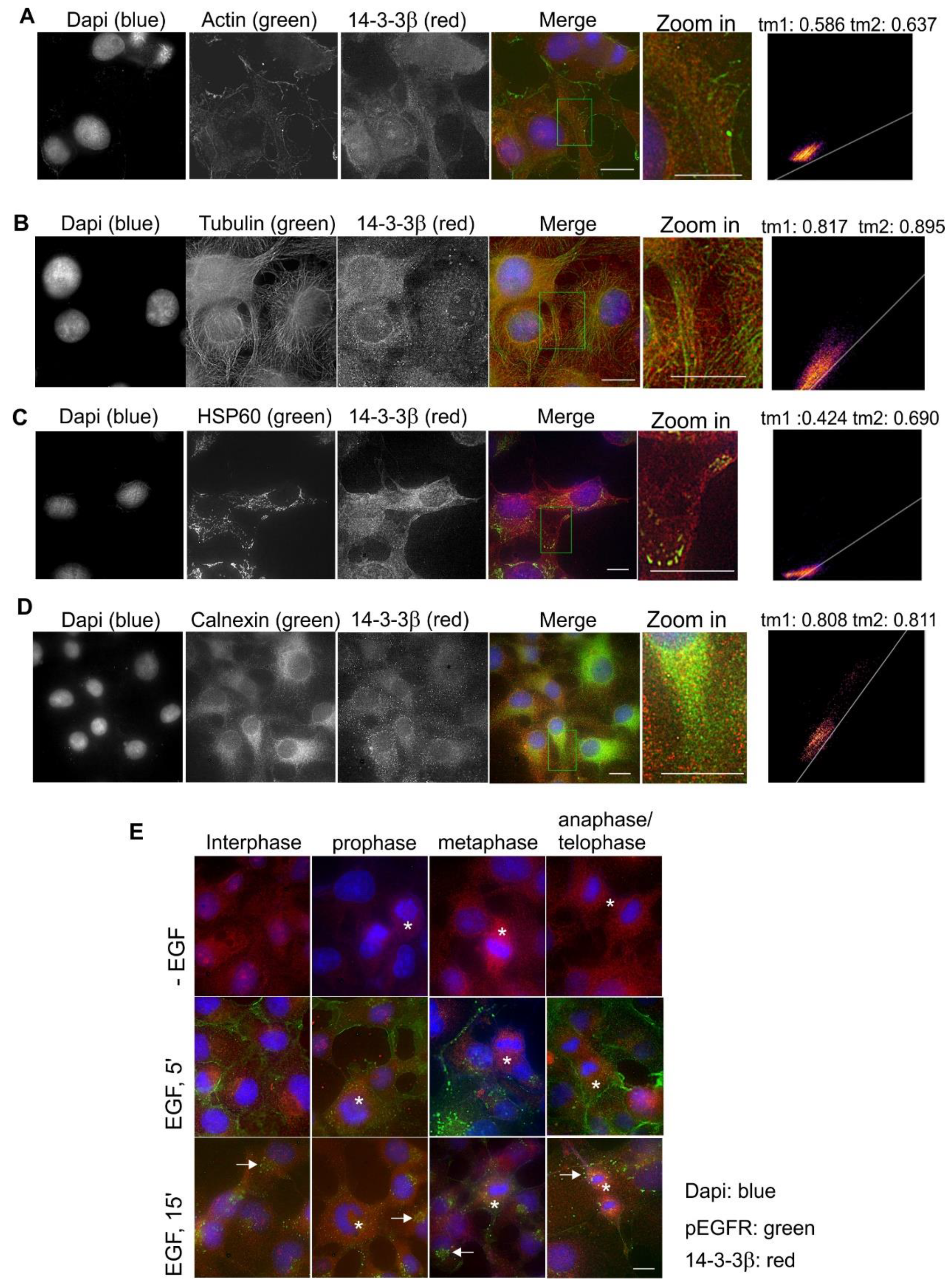
2.3. Subcellular Localization and Translocation of 14-3-3β in Response to EGF and During the Cell Cycle
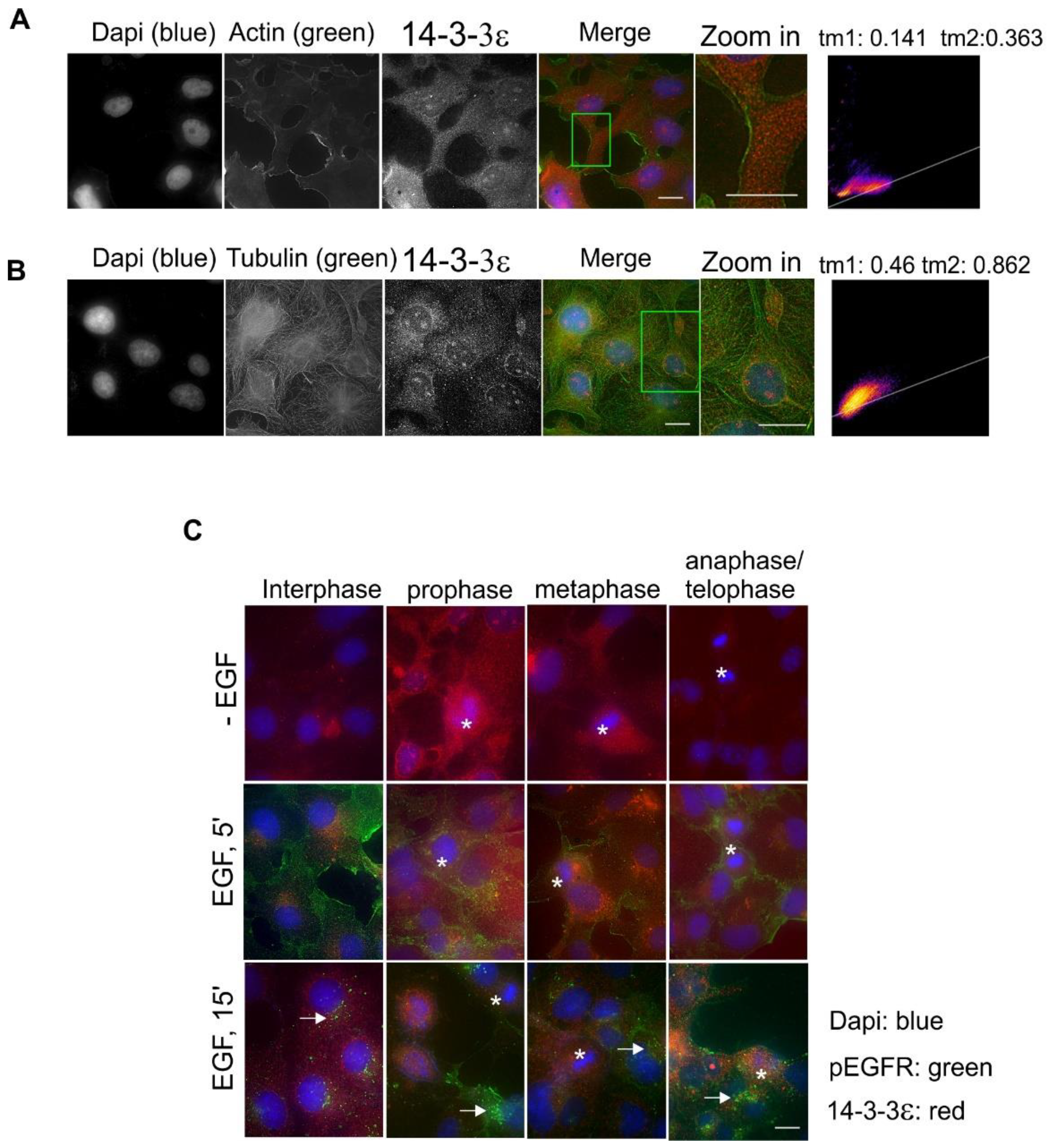
2.4. Subcellular Localization and Translocation of 14-3-3ε in Response to EGF and During the Cell Cycle
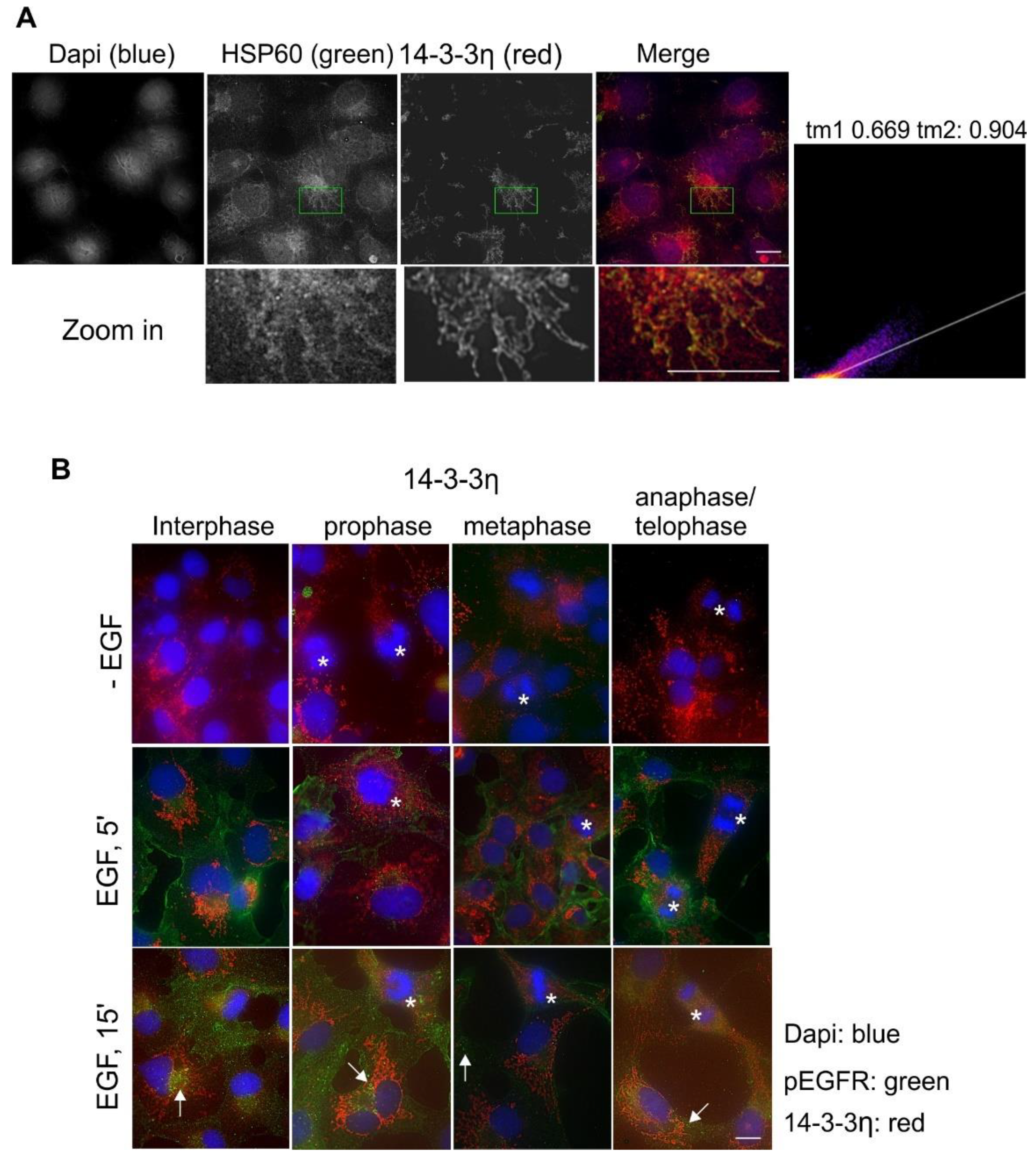
2.5. Subcellular Localization and Translocation of 14-3-3ƞ in Response to EGF and During the Cell Cycle
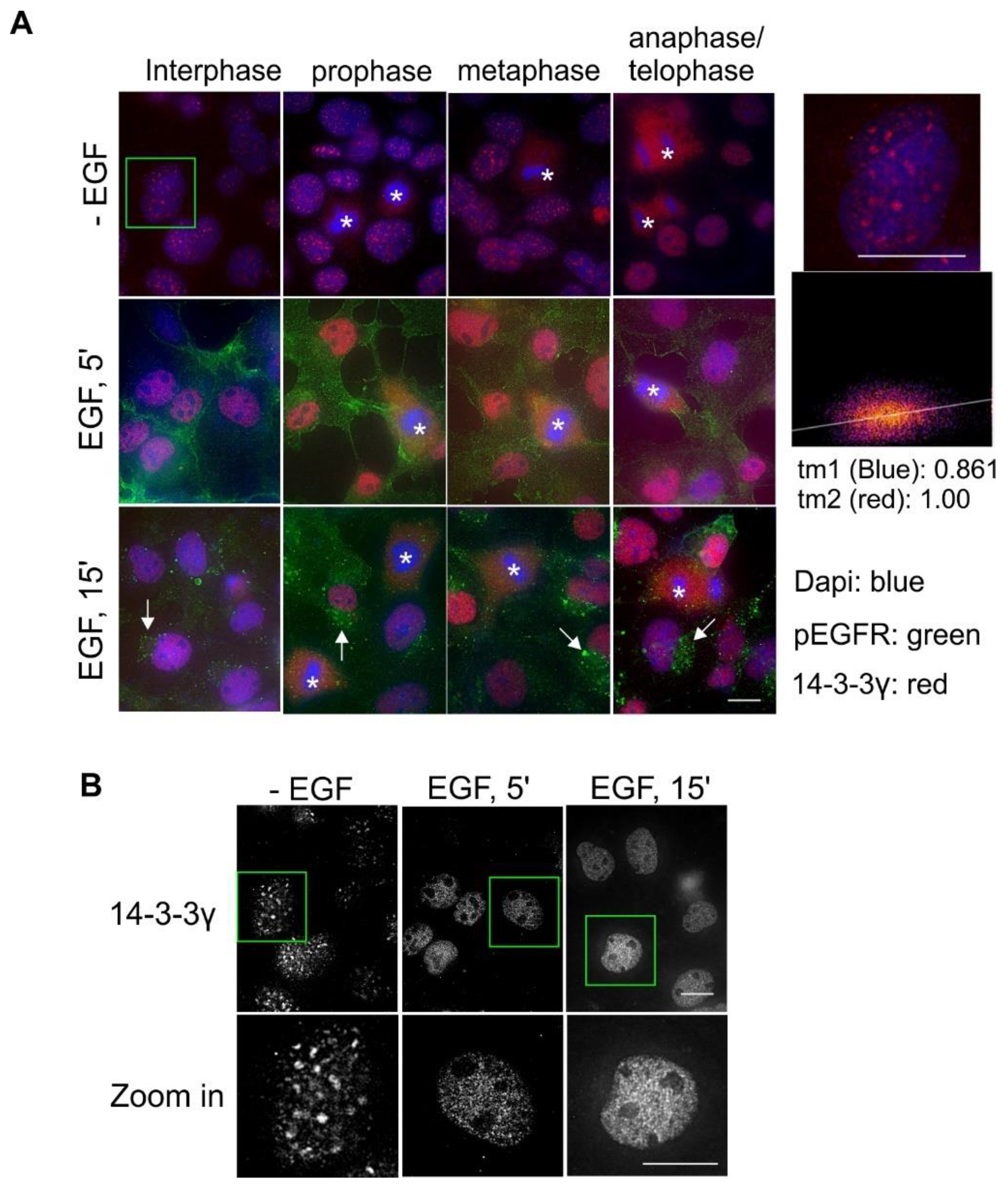
2.6. Subcellular Localization and Translocation of 14-3-3γ in Response to EGF and During the Cell Cycle
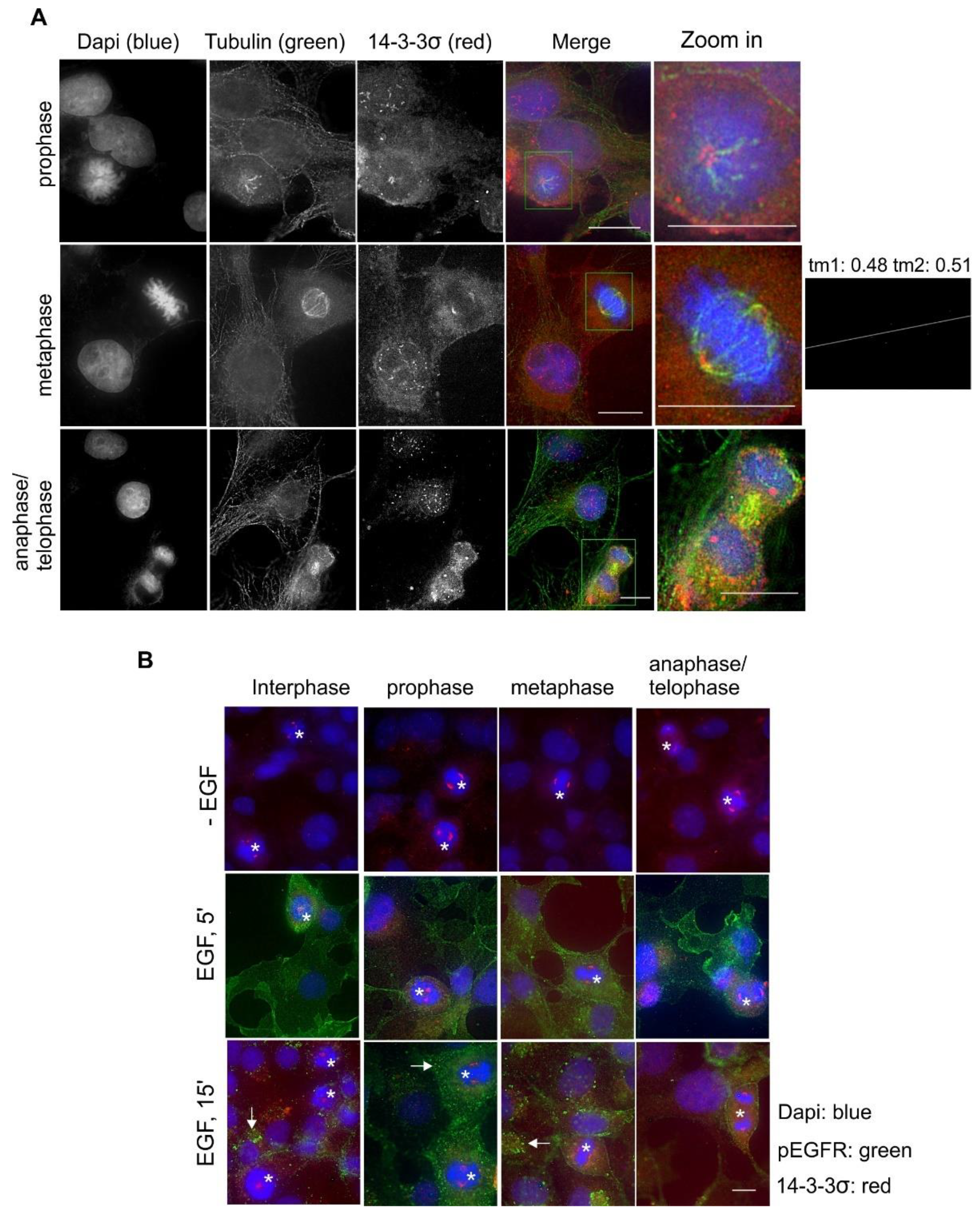
2.7. Subcellular Localization and Translocation of 14-3-3σ in Response to EGF and During the Cell Cycle
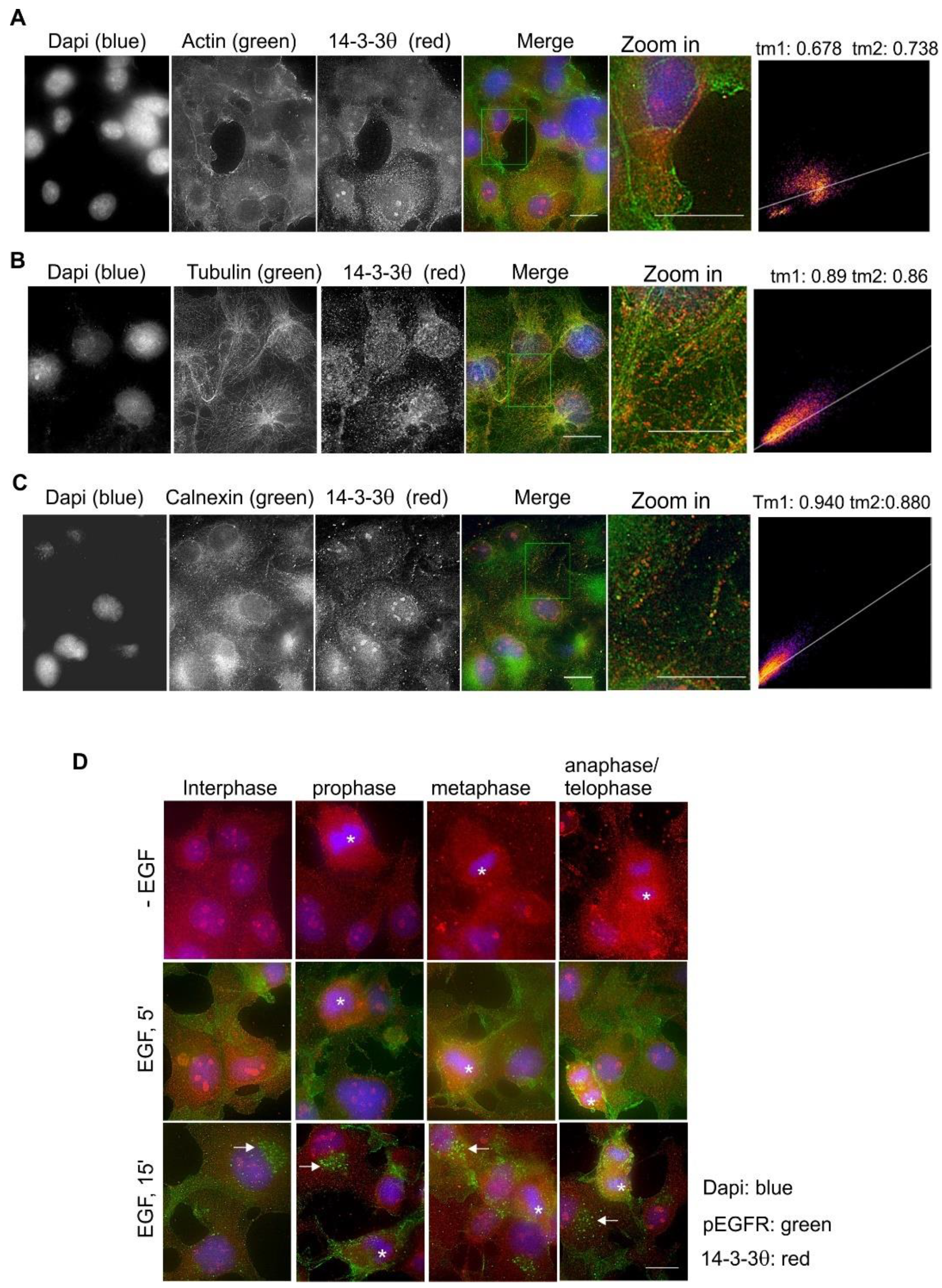
2.8. Subcellular Localization and Translocation of 14-3-3θ in response to EGF and During the Cell Cycle
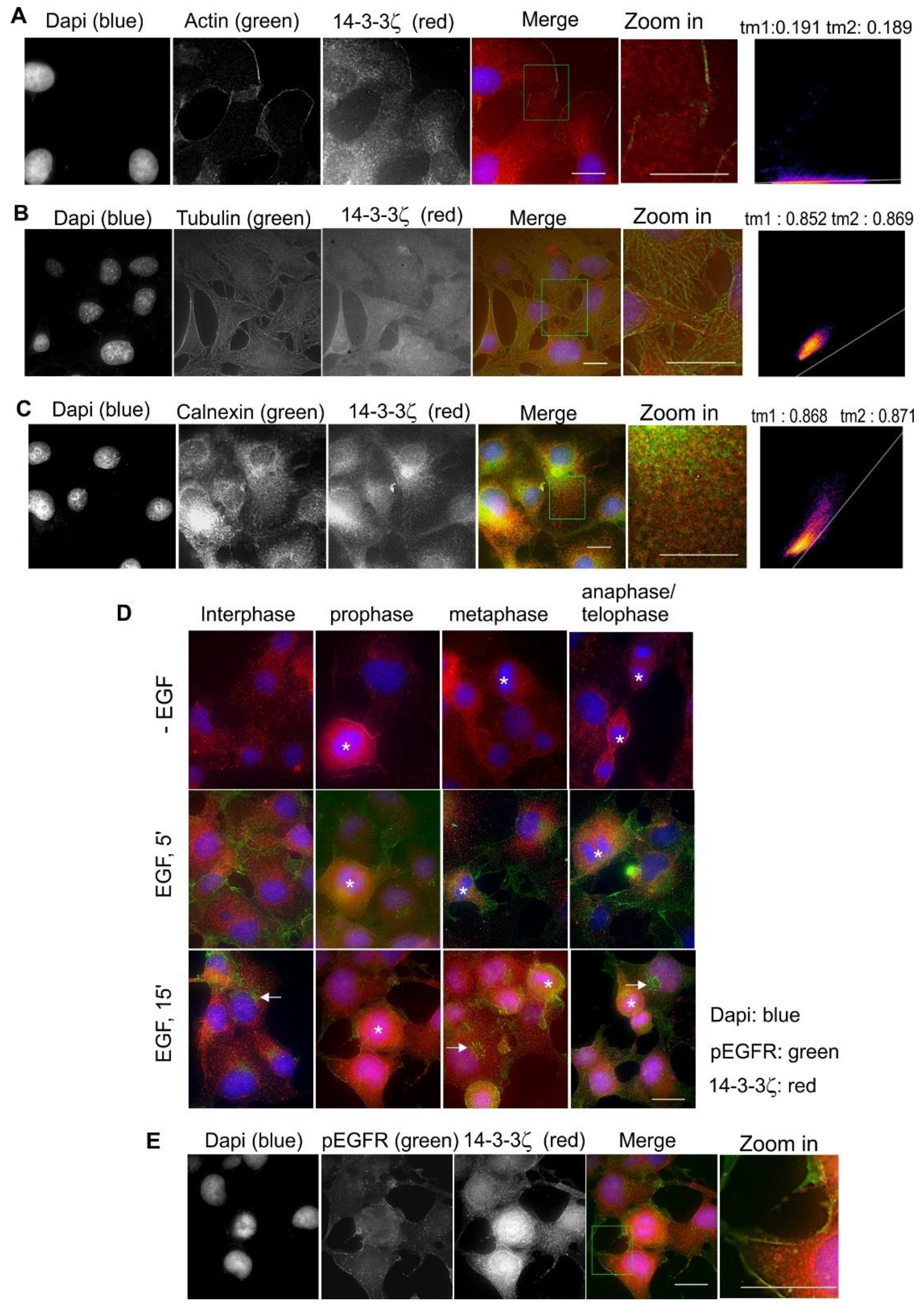
2.9. Subcellular Localization and Translocation of 14-3-3ζ in Response to EGF and During the Cell Cycle
2.10. The Expression and Subcellular Localization of Total 14-3-3 Protein and Each 14-3-3 Isoform in Human Cell Lines
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Culture and Treatment
5.2. Chemicals and Antibodies
5.3. Cell Lysates and Immunoblotting
5.4. Indirect Immunofluorescence
5.5. Co-Localization Analysis of IF Images by Mander′s Overlap Coefficient
5.6. Subcellular Fractionation
5.7. siRNA
5.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thomas, D.; Guthridge, M.; Woodcock, J.; Lopez, A. 14-3-3 protein Signal. in development and growth factor responses. Curr. Top. Dev. Biol. 2005, 67, 285–303. [Google Scholar] [PubMed]
- Ichimura, T.; Isobe, T.; Okuyama, T.; Takahashi, N.; Araki, K.; Kuwano, R.; Takahashi, Y. Molecular cloning of cDNA coding for brain-specific 14-3-3 protein, a protein kinase-dependent activator of tyrosine and tryptophan hydroxylases. Proc. Natl. Acad. Sci. USA 1988, 85, 7084–7088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toker, A.; Sellers, L.A.; Amess, B.; Patel, Y.; Harris, A.; Aitken, A. Multiple isoforms of a protein kinase C inhibitor (KCIP-1/14-3-3) from sheep brain. Amino acid sequence of phosphorylated forms. Eur. J. Biochem. 1992, 206, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Gardino, A.K.; Yaffe, M.B. 14-3-3 proteins as signaling integration points for cell cycle control and apoptosis. Semin. Cell Dev. Biol. 2011, 22, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh, Y.; Papadopoulos, V. The role of the 14-3-3 protein family in health, disease, and drug development. Drug Discov. Today 2016, 21, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Sluchanko, N.N.; Gusev, N.B. Moonlighting chaperone-like activity of the universal regulatory 14-3-3 proteins. FEBS J. 2017, 284, 1279–1295. [Google Scholar] [CrossRef] [Green Version]
- Freeman, A.K.; Morrison, D.K. 14-3-3 proteins: Diverse functions in cell proliferation and cancer progression. Semin. Cell Dev. Biol. 2011, 22, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Fan, Z.; Ding, M.; Zhang, H.; Mu, L.; Ding, Y.; Zhang, Y.; Jia, B.; Chen, L.; Chang, Z.; et al. An EGFR/PI3K/AKT axis promotes accumulation of the Rac1-GEF Tiam1 that is critical in EGFR-driven tumorigenesis. Oncogene 2015, 34, 5971–5982. [Google Scholar] [CrossRef]
- Moores, S.L.; Selfors, L.M.; Fredericks, J.; Breit, T.; Fujikawa, K.; Alt, F.W.; Brugge, J.S.; Swat, W. Vav family proteins couple to diverse cell surface receptors. Mol. Cell. Biol. 2000, 20, 6364–6373. [Google Scholar] [CrossRef]
- Cao, W.D.; Zhang, X.; Zhang, J.N.; Yang, Z.J.; Zhen, H.N.; Cheng, G.; Li, B.; Gao, D. Immunocytochemical detection of 14-3-3 in primary nervous system tumors. J. Neurooncol. 2006, 77, 125–130. [Google Scholar] [CrossRef]
- Lee, C.H.; Marino-Enriquez, A.; Ou, W.; Zhu, M.; Ali, R.H.; Chiang, S.; Amant, F.; Gilks, C.B.; van de Rijn, M.; Oliva, E.; et al. The clinicopathologic features of YWHAE-FAM22 endometrial stromal sarcomas: A histologically high-grade and clinically aggressive tumor. Am. J. Surg. Pathol. 2012, 36, 641–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masters, S.C.; Subramanian, R.R.; Truong, A.; Yang, H.; Fujii, K.; Zhang, H.; Fu, H. Survival-promoting functions of 14-3-3 proteins. Biochem. Soc. Trans. 2002, 30, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Hashizume, S.; Kato, M.; Honjoh, T.; Setoguchi, Y.; Yasumoto, K. Elevated expression levels of the 14-3-3 family of proteins in lung cancer tissues. Hum. Antibodies 1997, 8, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, V.M.; Martinez, J.D. 14-3-3gamma induces oncogenic transformation by stimulating MAP kinase and PI3K signaling. PLoS ONE 2010, 5, e11433. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Nesland, J.M.; Suo, Z.; Trope, C.G.; Holm, R. The prognostic value of 14-3-3 isoforms in vulvar squamous cell carcinoma cases: 14-3-3beta and epsilon are independent prognostic factors for these tumors. PLoS ONE 2011, 6, e24843. [Google Scholar] [CrossRef]
- Qi, W.; Martinez, J.D. Reduction of 14-3-3 proteins correlates with increased sensitivity to killing of human lung cancer cells by ionizing radiation. Radiat. Res. 2003, 160, 217–223. [Google Scholar] [CrossRef]
- Xiao, Y.; Lin, V.Y.; Ke, S.; Lin, G.E.; Lin, F.T.; Lin, W.C. 14-3-3tau promotes breast cancer invasion and metastasis by inhibiting RhoGDIalpha. Mol. Cell. Biol. 2014, 34, 2635–2649. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.Y.; Jeon, W.K.; Bae, E.J.; Kim, S.T.; Lee, H.J.; Kim, S.J.; Kim, B.C. 14-3-3 sigma and 14-3-3 zeta plays an opposite role in cell growth inhibition mediated by transforming growth factor-beta 1. Mol. Cells 2010, 29, 305–309. [Google Scholar] [CrossRef]
- Hermeking, H.; Lengauer, C.; Polyak, K.; He, T.C.; Zhang, L.; Thiagalingam, S.; Kinzler, K.W.; Vogelstein, B. 14-3-3sigma is a p53-regulated inhibitor of G2/M progression. Mol. Cell 1997, 1, 3–11. [Google Scholar] [CrossRef]
- Cheng, L.; Pan, C.X.; Zhang, J.T.; Zhang, S.; Kinch, M.S.; Li, L.; Baldridge, L.A.; Wade, C.; Hu, Z.; Koch, M.O.; et al. Loss of 14-3-3sigma in prostate cancer and its precursors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 3064–3068. [Google Scholar] [CrossRef] [Green Version]
- Moreira, J.M.; Gromov, P.; Celis, J.E. Expression of the tumor suppressor protein 14-3-3 sigma is down-regulated in invasive transitional cell carcinomas of the urinary bladder undergoing epithelial-to-mesenchymal transition. Mol. Cell. Proteom. 2004, 3, 410–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, D.; Chen, Y.; Karia, B.; Brown, A.; Gu, T.T.; Li, J.; Carey, M.S.; Hennessy, B.T.; Bishop, A.J. 14-3-3 sigma expression effects G2/M response to oxygen and correlates with ovarian cancer metastasis. PLoS ONE 2011, 6, e15864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goc, A.; Abdalla, M.; Al-Azayzih, A.; Somanath, P.R. Rac1 activation driven by 14-3-3zeta dimerization promotes prostate cancer cell-matrix interactions, motility and transendothelial migration. PLoS ONE 2012, 7, e40594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal, C.L.; Yu, D. 14-3-3zeta as a prognostic marker and therapeutic target for cancer. Expert Opin Ther. Targets 2010, 14, 1343–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mhawech, P. 14-3-3 proteins—An update. Cell Res. 2005, 15, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Tseng, H.C.; Yang, C.M. Bradykinin-mediated cell proliferation depends on transactivation of EGF receptor in corneal fibroblasts. J. Cell. Physiol. 2012, 227, 1367–1381. [Google Scholar] [CrossRef]
- Rittinger, K.; Budman, J.; Xu, J.; Volinia, S.; Cantley, L.C.; Smerdon, S.J.; Gamblin, S.J.; Yaffe, M.B. Structural analysis of 14-3-3 phosphopeptide complexes identifies a dual role for the nuclear export signal of 14-3-3 in ligand binding. Mol. Cell 1999, 4, 153–166. [Google Scholar] [CrossRef]
- Yaffe, M.B.; Rittinger, K.; Volinia, S.; Caron, P.R.; Aitken, A.; Leffers, H.; Gamblin, S.J.; Smerdon, S.J.; Cantley, L.C. The structural basis for 14-3-3: phosphopeptide binding specificity. Cell 1997, 91, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Coblitz, B.; Wu, M.; Shikano, S.; Li, M. C-terminal binding: An expanded repertoire and function of 14-3-3 proteins. FEBS Lett. 2006, 580, 1531–1535. [Google Scholar] [CrossRef] [Green Version]
- Shikano, S.; Coblitz, B.; Sun, H.; Li, M. Genetic isolation of transport signals directing cell surface expression. Nat. Cell Biol. 2005, 7, 985–992. [Google Scholar] [CrossRef]
- Paul, A.L.; Sehnke, P.C.; Ferl, R.J. Isoform-specific subcellular localization among 14-3-3 proteins in Arabidopsis seems to be driven by client interactions. Mol. Biol. Cell 2005, 16, 1735–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.T.; Parry, D.H.; Donahoe, B.; Chien, C.T.; O’Farrell, P.H.; Purdy, A. Cell cycle roles for two 14-3-3 proteins during Drosophila development. J. Cell Sci. 2001, 114, 3445–3454. [Google Scholar] [PubMed]
- Pietromonaco, S.F.; Seluja, G.A.; Aitken, A.; Elias, L. Association of 14-3-3 proteins with centrosomes. Blood Cells Mol. Dis. 1996, 22, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; Sehgal, L.; Bose, A.; Gulvady, A.; Senapati, P.; Thorat, R.; Basu, S.; Bhatt, K.; Hosing, A.S.; Balyan, R.; et al. 14-3-3gamma prevents centrosome amplification and neoplastic progression. Sci. Rep. 2016, 6, 26580. [Google Scholar] [CrossRef] [Green Version]
- Schindler, C.K.; Heverin, M.; Henshall, D.C. Isoform- and subcellular fraction-specific differences in hippocampal 14-3-3 levels following experimentally evoked seizures and in human temporal lobe epilepsy. J. Neurochem. 2006, 99, 561–569. [Google Scholar] [CrossRef]
- Lamba, S.; Ravichandran, V.; Major, E.O. Glial cell type-specific subcellular localization of 14-3-3 zeta: An implication for JCV tropism. Glia 2009, 57, 971–977. [Google Scholar] [CrossRef] [Green Version]
- Jerome, M.; Paudel, H.K. 14-3-3zeta regulates nuclear trafficking of protein phosphatase 1alpha (PP1alpha) in HEK-293 cells. Arch. Biochem. Biophys. 2014, 558, 28–35. [Google Scholar] [CrossRef]
- Chun, J.; Kwon, T.; Lee, E.J.; Kim, C.H.; Han, Y.S.; Hong, S.K.; Hyun, S.; Kang, S.S. 14-3-3 protein mediates phosphorylation of microtubule-associated protein tau by serum- and glucocorticoid-induced protein kinase 1. Mol. Cells 2004, 18, 360–368. [Google Scholar]
- Inamdar, S.M.; Lankford, C.K.; Laird, J.G.; Novbatova, G.; Tatro, N.; Whitmore, S.S.; Scheetz, T.E.; Baker, S.A. Analysis of 14-3-3 isoforms expressed in photoreceptors. Exp. Eye Res. 2018, 170, 108–116. [Google Scholar] [CrossRef]
- Aitken, A. Functional specificity in 14-3-3 isoform interactions through dimer formation and phosphorylation. Chromosome location of mammalian isoforms and variants. Plant Mol. Biol. 2002, 50, 993–1010. [Google Scholar] [CrossRef]
- Sluchanko, N.N.; Gusev, N.B. 14-3-3 proteins and regulation of cytoskeleton. Biochemistry (Moscow) 2010, 75, 1528–1546. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.; Holzmann, C.; Riess, O. 14-3-3 proteins in the nervous system. Nat. Rev. Neurosci. 2003, 4, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Aaron, J.S.; Taylor, A.B.; Chew, T.L. Image co-localization-co-occurrence versus correlation. J. Cell. Sci. 2018, 131, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamburg, J.R. Proteins of the ADF/cofilin family: Essential regulators of actin dynamics. Annu. Rev. Cell Dev. Biol. 1999, 15, 185–230. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Yang, Y.; Cai, X.; Liu, L.; Wu, K.; Yu, M. Down-regulation of 14-3-3 zeta inhibits TGF-beta1-induced actomyosin contraction in human trabecular meshwork cells through RhoA signaling pathway. Investig. Ophthalmol. Vis. Sci. 2016, 57, 719–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kligys, K.; Yao, J.; Yu, D.; Jones, J.C. 14-3-3zeta/tau heterodimers regulate Slingshot activity in migrating keratinocytes. Biochem. Biophys. Res. Commun. 2009, 383, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Krtkova, J.; Xu, J.; Lalle, M.; Steele-Ogus, M.; Alas, G.C.M.; Sept, D.; Paredez, A.R. 14-3-3 regulates actin filament formation in the deep-branching eukaryote Giardia lamblia. mSphere 2017, 2, e00248-17. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, H.Y.; Han, D.; MacDonald, R.; Paudel, H.K. Overexpression of 14-3-3z promotes tau phosphorylation at Ser262 and accelerates proteosomal degradation of synaptophysin in rat primary hippocampal neurons. PLoS ONE 2013, 8, e84615. [Google Scholar] [CrossRef] [Green Version]
- Cornell, B.; Wachi, T.; Zhukarev, V.; Toyo-Oka, K. Regulation of neuronal morphogenesis by 14-3-3epsilon (Ywhae) via the microtubule binding protein, doublecortin. Hum. Mol. Genet. 2016, 25, 4405–4418. [Google Scholar] [CrossRef] [Green Version]
- Park, G.Y.; Han, J.Y.; Han, Y.K.; Kim, S.D.; Kim, J.S.; Jo, W.S.; Chun, S.H.; Jeong, D.H.; Lee, C.W.; Yang, K.; et al. 14-3-3 eta depletion sensitizes glioblastoma cells to irradiation due to enhanced mitotic cell death. Cancer Gene Ther. 2014, 21, 158–163. [Google Scholar] [CrossRef]
- Joo, Y.; Schumacher, B.; Landrieu, I.; Bartel, M.; Smet-Nocca, C.; Jang, A.; Choi, H.S.; Jeon, N.L.; Chang, K.A.; Kim, H.S.; et al. Involvement of 14-3-3 in tubulin instability and impaired axon development is mediated by Tau. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 4133–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, S.; Melkova, K.; Trosanova, Z.; Hanakova, K.; Zachrdla, M.; Novacek, J.; Zupa, E.; Zdrahal, Z.; Hritz, J.; Zidek, L. Quantitative mapping of microtubule-associated protein 2c (MAP2c) phosphorylation and regulatory protein 14-3-3zeta-binding sites reveals key differences between MAP2c and its homolog Tau. J. Biol. Chem. 2017, 292, 6715–6727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Kee, Y.S.; Poirier, C.C.; Jelinek, C.; Osborne, J.; Divi, S.; Surcel, A.; Will, M.E.; Eggert, U.S.; Muller-Taubenberger, A.; et al. 14-3-3 coordinates microtubules, Rac, and myosin II to control cell mechanics and cytokinesis. Curr. Biol. 2010, 20, 1881–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, D.N. 14-3-3, an integrator of cell mechanics and cytokinesis. Small GTPases 2010, 1, 165–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Yang, L.; Huang, J.; Xu, P.; Zhang, Z.; Yin, D.; Liu, J.; He, H.; He, M. Luteoloside attenuates anoxia/reoxygenation-induced cardiomyocytes injury via mitochondrial pathway mediated by 14-3-3eta protein. Phytother. Res. 2018, 32, 1126–1134. [Google Scholar] [CrossRef]
- Sreedhar, R.; Arumugam, S.; Thandavarayan, R.A.; Giridharan, V.V.; Karuppagounder, V.; Pitchaimani, V.; Afrin, R.; Miyashita, S.; Nomoto, M.; Harima, M.; et al. Myocardial 14-3-3eta protein protects against mitochondria mediated apoptosis. Cell. Signal. 2015, 27, 770–776. [Google Scholar] [CrossRef]
- Huang, J.; Liu, Z.; Xu, P.; Zhang, Z.; Yin, D.; Liu, J.; He, H.; He, M. Capsaicin prevents mitochondrial damage, protects cardiomyocytes subjected to anoxia/reoxygenation injury mediated by 14-3-3eta/Bcl-2. Eur. J. Pharmacol. 2018, 819, 43–50. [Google Scholar] [CrossRef]
- Zhang, Z.; He, H.; Qiao, Y.; Huang, J.; Wu, Z.; Xu, P.; Yin, D.; He, M. Tanshinone IIa pretreatment protects H9c2 cells against anoxia/reoxygenation injury: Involvement of the translocation of Bcl-2 to mitochondria mediated by 14-3-3eta. Oxidative Med. Cell. Longev. 2018, 2018, 3583921. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; You, J.; Qiao, Y.; Wu, Z.; Liu, D.; Yin, D.; He, H.; He, M. Tetramethylpyrazine attenuates lipopolysaccharide-induced cardiomyocyte injury via improving mitochondrial function mediated by 14-3-3gamma. Eur. J. Pharmacol. 2018, 832, 67–74. [Google Scholar] [CrossRef]
- He, H.; Luo, Y.; Qiao, Y.; Zhang, Z.; Yin, D.; Yao, J.; You, J.; He, M. Curcumin attenuates doxorubicin-induced cardiotoxicity via suppressing oxidative stress and preventing mitochondrial dysfunction mediated by 14-3-3gamma. Food Funct. 2018, 9, 4404–4418. [Google Scholar] [CrossRef]
- Tollenaere, M.A.; Villumsen, B.H.; Blasius, M.; Nielsen, J.C.; Wagner, S.A.; Bartek, J.; Beli, P.; Mailand, N.; Bekker-Jensen, S. P38- and MK2-dependent signalling promotes stress-induced centriolar satellite remodelling via 14-3-3-dependent sequestration of CEP131/AZI1. Nat. Commun. 2015, 6, 10075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhri, M.; Scarabel, M.; Aitken, A. Mammalian and yeast 14-3-3 isoforms form distinct patterns of dimers in vivo. Biochem. Biophys. Res. Commun. 2003, 300, 679–685. [Google Scholar] [CrossRef]
- Mao, Y.S.; Zhang, B.; Spector, D.L. Biogenesis and function of nuclear bodies. Trends Genet. 2011, 27, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z. ErbB receptors and cancer. Methods Mol. Biol. 2017, 1652, 3–35. [Google Scholar] [PubMed]
- Abdrabou, A.; Wang, Z. Post-translational modification and subcellular distribution of Rac1: An update. Cells 2018, 7, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Pennock, S.; Chen, X.; Wang, Z. Endosomal signaling of epidermal growth factor receptor stimulates signal transduction pathways leading to cell survival. Mol. Cell. Biol. 2002, 22, 7279–7290. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 14-3-3 isoform | Subcellular Localization |
|---|---|
| Beta | Cytoplasmic, ER and Mitochondria |
| Zeta | Cytoplasmic and Nuclear |
| Gamma | Nucleus |
| Epsilon | Cytoplasmic |
| Eta | Mitochondria |
| Tau/Theta | Cytoplasmic and ER |
| Sigma | Centrosome |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdrabou, A.; Brandwein, D.; Wang, Z. Differential Subcellular Distribution and Translocation of Seven 14-3-3 Isoforms in Response to EGF and During the Cell Cycle. Int. J. Mol. Sci. 2020, 21, 318. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010318
Abdrabou A, Brandwein D, Wang Z. Differential Subcellular Distribution and Translocation of Seven 14-3-3 Isoforms in Response to EGF and During the Cell Cycle. International Journal of Molecular Sciences. 2020; 21(1):318. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010318
Chicago/Turabian StyleAbdrabou, Abdalla, Daniel Brandwein, and Zhixiang Wang. 2020. "Differential Subcellular Distribution and Translocation of Seven 14-3-3 Isoforms in Response to EGF and During the Cell Cycle" International Journal of Molecular Sciences 21, no. 1: 318. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010318