MicroRNA-204 Deficiency in Human Aortic Valves Elevates Valvular Osteogenic Activity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
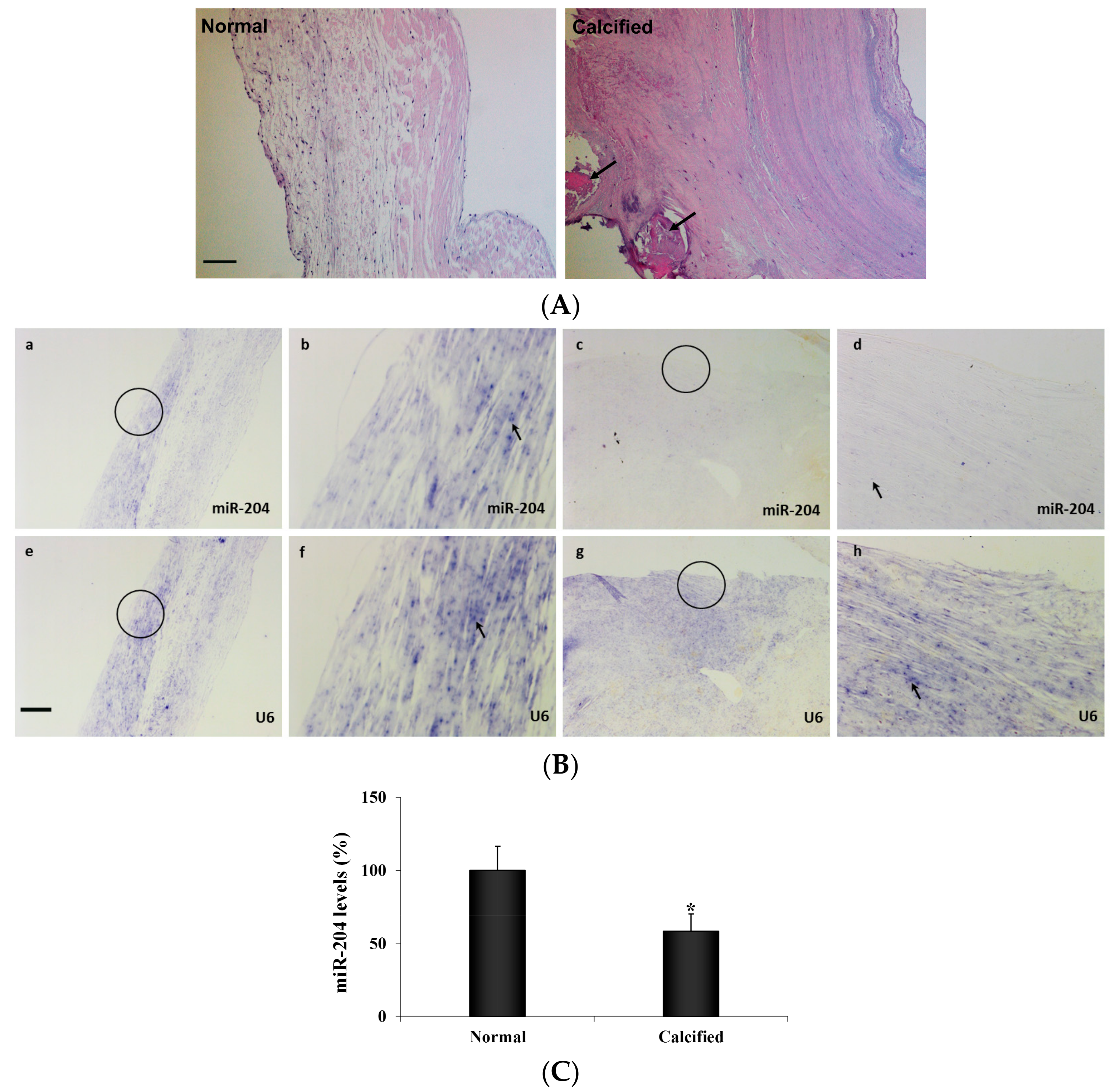
2.1. Diseased Human Aortic Valves are Deficient in MiR-204
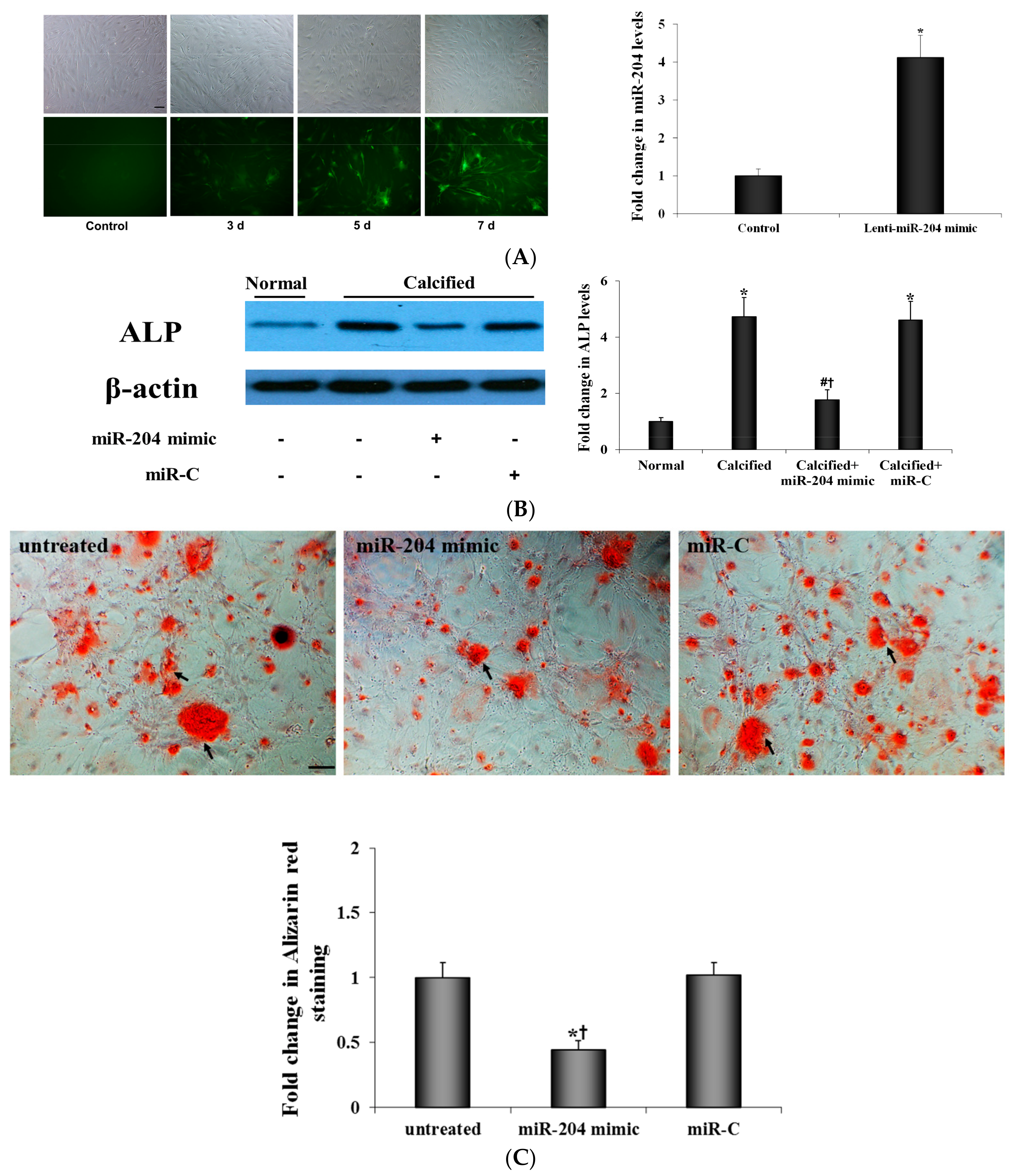
2.2. MiR-204 Mimic Suppresses the Osteogenic Activity in AVICs from Diseased Valves
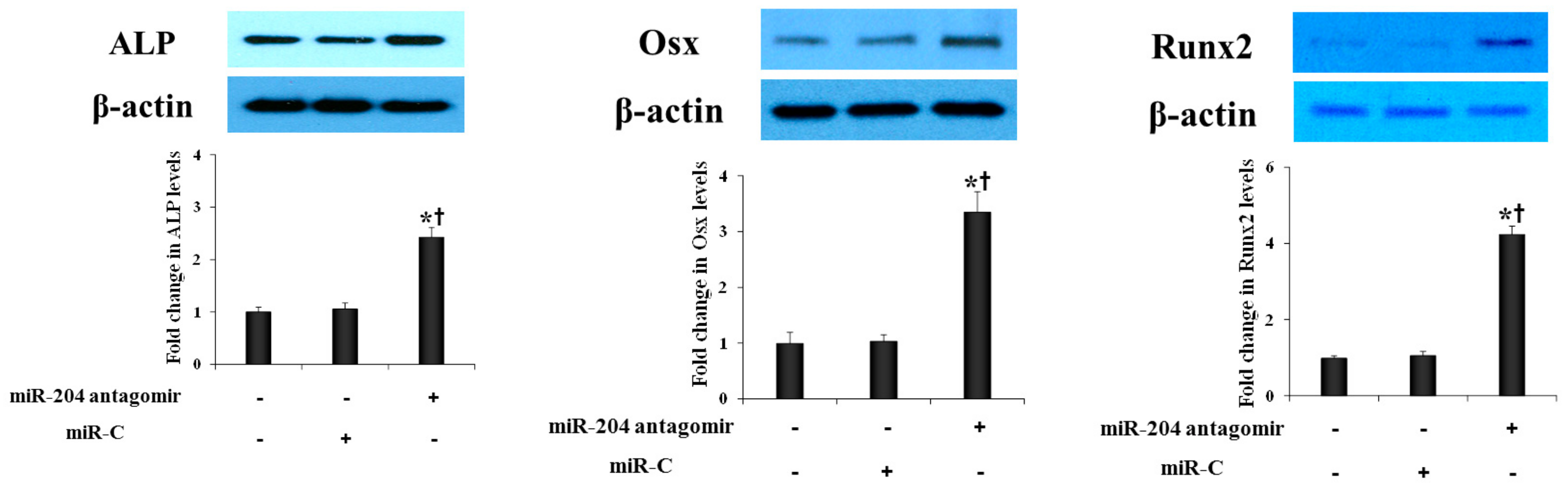
2.3. MiR-204 Antagomir Enhances the Expression of Osteogenic Biomarkers in AVICs from Normal Human Aortic Valves
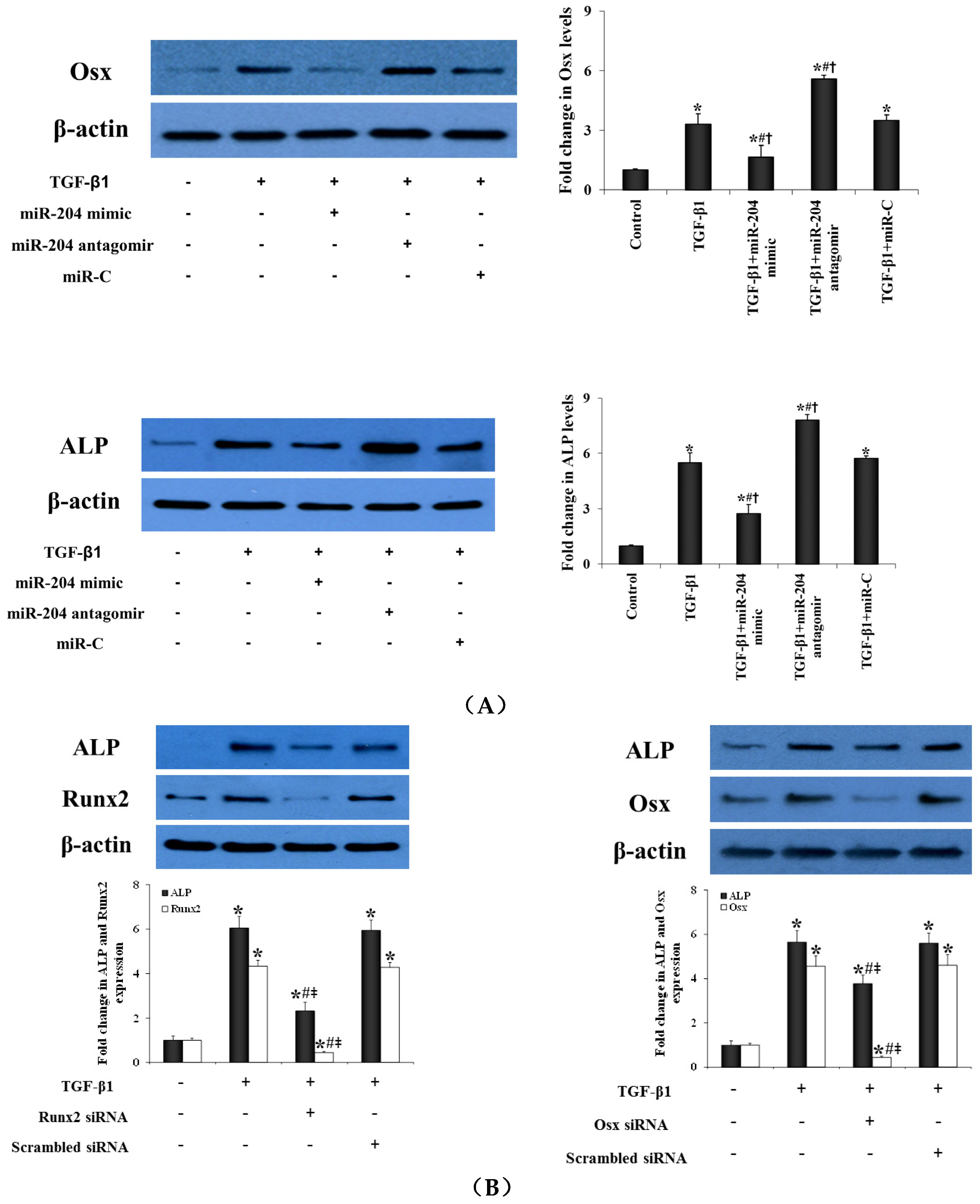
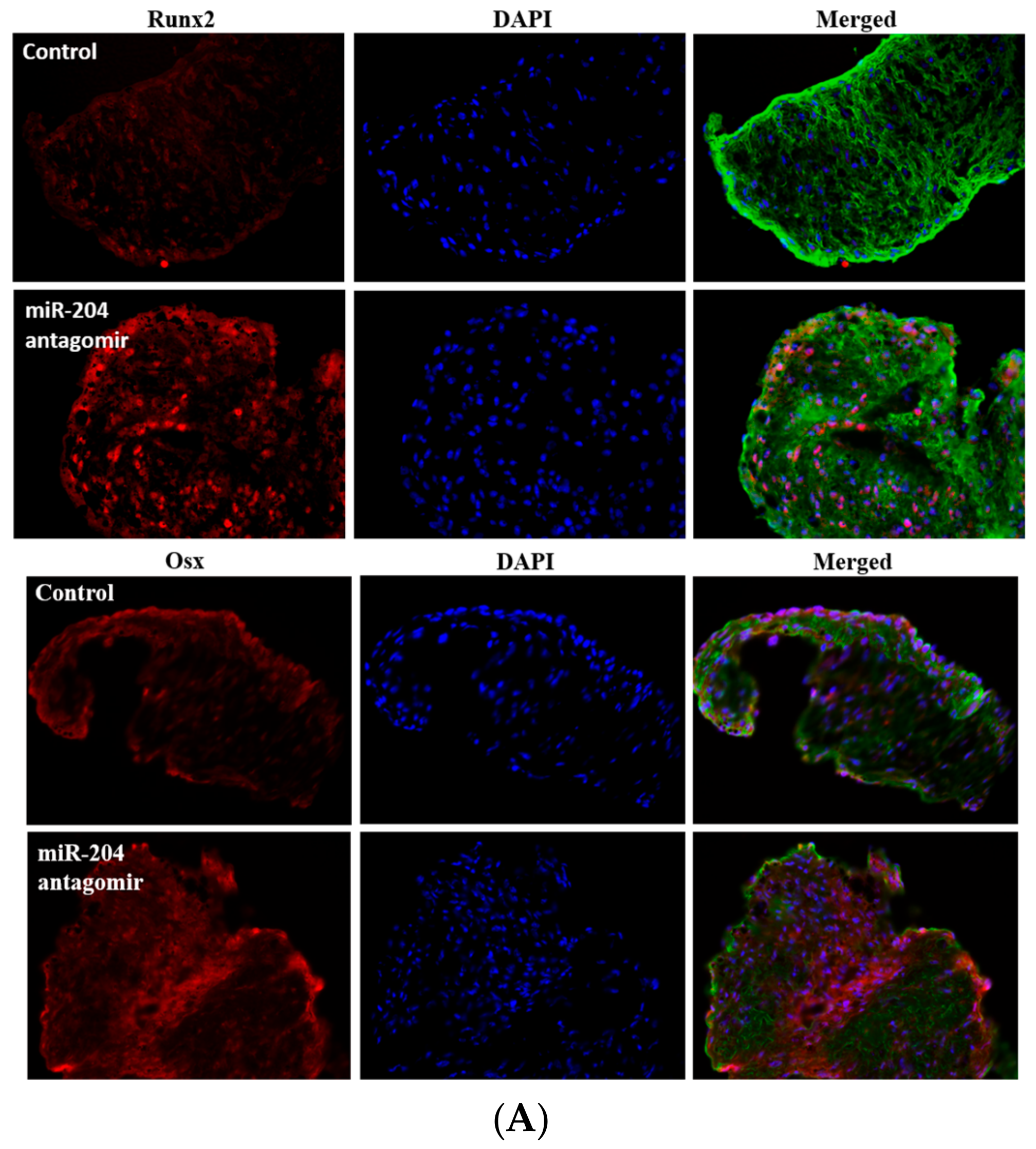
2.4. MiR-204 Up-Regulates the Expression of Runx2 and Osx in Human AVICs to Induce the Transition to Pro-Osteogenic Phenotype
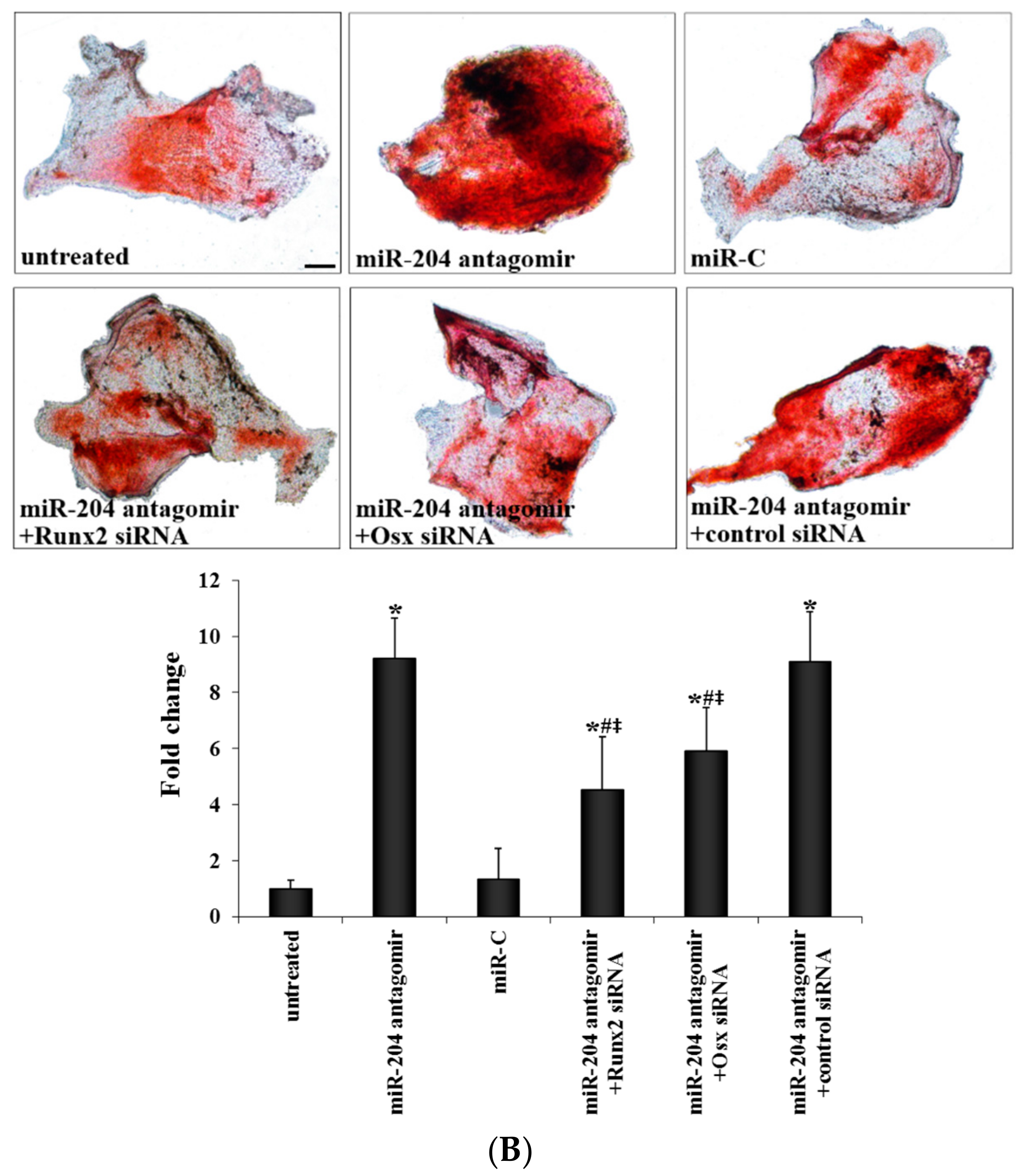
2.5. Expression of MiR-204 Antagomir Promotes Calcium Deposit Formation in Aortic Valves
3. Discussion
3.1. Reduced Expression of MiR-204 in Diseased Aortic Valves Contributes to the Mechanism Underlying AVIC Pro-Osteogenic Phenotype
3.2. MiR-204 Suppresses Valvular Osteogenic Activity through Down-Regulation of Runx2 and Osx
3.3. Summary and Limitations
4. Materials and Methods
4.1. Materials
4.2. Isolation, Culture and Treatment of Human AVICs
4.3. Mouse Aortic Valve Culture
4.4. Preparation of Recombinant Lentiviral Supernatants and Lentiviral Transduction
4.5. Immunoblotting
4.6. Real-Time RT-PCR Analysis
4.7. In Situ Hybridization
4.8. Histology and Immunofluorescent Staining
4.9. Staining for Calcium Deposits
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Messika-Zeitoun, D.; Bielak, L.F.; Peyser, P.A.; Sheedy, P.F.; Turner, S.T.; Nkomo, V.T.; Breen, J.F.; Maalouf, J.; Scott, C.; Tajik, A.J.; et al. Aortic valve calcification: Determinants and progression in the population. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 642–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dweck, M.R.; Boon, N.A.; Newby, D.E. Calcific aortic stenosis: A disease of the valve and the myocardium. J. Am. Coll. Cardiol. 2012, 60, 1854–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohukainen, P.; Ruskoaho, H.; Rysa, J. Cellular mechanisms of valvular thickening in early and intermediate calcific aortic valve disease. Curr. Cardiol. Rev. 2018, 14, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Kostyunin, A.E.; Yuzhalin, A.E.; Ovcharenko, E.A.; Kutikhin, A.G. Development of calcific aortic valve disease: Do we know enough for new clinical trials? J. Mol. Cell. Cardiol. 2019, 132, 189–209. [Google Scholar] [CrossRef]
- Mohler, E.R., III. Mechanisms of aortic valve calcification. Am. J. Cardiol. 2004, 94, 1396–1402. [Google Scholar] [CrossRef]
- New, S.E.; Aikawa, E. Molecular imaging insights into early inflammatory stages of arterial and aortic valve calcification. Circ. Res. 2011, 108, 1381–1391. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.C.; Joag, V.R.; Gotlieb, A.I. The emerging role of valve interstitial cell phenotypes in regulating heart valve pathobiology. Am. J. Pathol. 2007, 171, 1407–1418. [Google Scholar] [CrossRef] [Green Version]
- Bogdanova, M.; Zabirnyk, A.; Malashicheva, A.; Enayati, K.Z.; Karlsen, T.A.; Kaljusto, M.-L.; Kvitting, J.-P.E.; Dissen, E.; Sullivan, G.J.; Kostareva, A. Interstitial cells in calcified aortic valves have reduced differentiation potential and stem cell-like properties. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Hafizi, S.; Taylor, P.M.; Chester, A.H.; Allen, S.P.; Yacoub, M.H. Mitogenic and secretory responses of human valve interstitial cells to vasoactive agents. J. Heart Valve Dis. 2000, 9, 454–458. [Google Scholar]
- Chester, A.H.; Taylor, P.M. Molecular and functional characteristics of heart-valve interstitial cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1437–1443. [Google Scholar] [CrossRef] [Green Version]
- Osman, L.; Chester, A.H.; Amrani, M.; Yacoub, M.H.; Smolenski, R.T. A novel role of extracellular nucleotides in valve calcification: A potential target for atorvastatin. Circulation 2006, 114 (Suppl. 1), I566–I572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.H.; Simmons, C.A. Cell-matrix interactions in the pathobiology of calcific aortic valve disease: Critical roles for matricellular, matricrine, and matrix mechanics cues. Circ. Res. 2011, 108, 1510–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yip, C.Y.; Blaser, M.C.; Mirzaei, Z.; Zhong, X.; Simmons, C.A. Inhibition of pathological differentiation of valvular interstitial cells by C-type natriuretic peptide. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1881–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, A.; Kiss, D.; Giri, J.; Anwaruddin, S.; Siddiqi, H.; Guerraty, M.; Chirinos, J.A.; Ferrari, G.; Rader, D.J. Biomarkers of calcific aortic valve disease. Atertio. Thromb. Vasc. Biol. 2017, 37, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Towler, D.A. Molecular and cellular aspects of calcific aortic valve disease. Circul. Res. 2013, 113, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Krohn, J.B.; Hutcheson, J.D.; Martínez-Martínez, E.; Irvin, W.S.; Bouten, C.V.; Bertazzo, S.; Bendeck, M.P.; Aikawa, E. Discoidin domain receptor-1 regulates calcific extracellular vesicle release in vascular smooth muscle cell fibrocalcific response via transforming growth factor-β signaling. Atertio. Thromb. Vasc. Biol. 2016, 36, 525–533. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Chang, Q.; Xin, M.; Wang, Q.; Li, H.; Qian, J. Endogenous bone morphogenetic protein 2 plays a role in vascular smooth muscle cell calcification induced by interleukin 6 in vitro. Int. J. Immunopathol. Pharmacol. 2017, 30, 227–237. [Google Scholar] [CrossRef]
- Luong, T.T.; Estepa, M.; Boehme, B.; Pieske, B.; Lang, F.; Eckardt, K.-U.; Voelkl, J.; Alesutan, I. Inhibition of vascular smooth muscle cell calcification by vasorin through interference with TGFβ1 signaling. Cell. Signal. 2019, 64, 109414. [Google Scholar] [CrossRef]
- Jian, B.; Narula, N.; Li, Q.Y.; Mohler, E.R., III; Levy, R.J. Progression of aortic valve stenosis: TGF-beta1 is present in calcified aortic valve cusps and promotes aortic valve interstitial cell calcification via apoptosis. Ann. Thorac. Surg. 2003, 75, 457–466. [Google Scholar] [CrossRef]
- Osman, L.; Yacoub, M.H.; Latif, N.; Amrani, M.; Chester, A.H. Role of human valve interstitial cells in valve calcification and their response to atorvastatin. Circulation 2006, 114 (Suppl. 1), I547–I552. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Meng, X.; Su, X.; Mauchley, D.C.; Ao, L.; Cleveland, J.C., Jr.; Fullerton, D.A. Bone morphogenic protein 2 induces Runx2 and osteopontin expression in human aortic valve interstitial cells: Role of Smad1 and extracellular signal-regulated kinase 1/2. J. Thorac. Cardiovasc. Surg. 2009, 138, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, R.; Fullerton, D.A.; Ao, L.; Zheng, D.; Zhao, K.-s.; Meng, X. BMP-2 and TGF-β1 mediate biglycan-induced pro-osteogenic reprogramming in aortic valve interstitial cells. J. Mol. Med. 2015, 93, 403–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bostrom, K.I.; Rajamannan, N.M.; Towler, D.A. The regulation of valvular and vascular sclerosis by osteogenic morphogens. Circ. Res. 2011, 109, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, R.; Hata, K.; Matsubara, T.; Wakabayashi, M.; Yoneda, T. Regulation of bone and cartilage development by network between BMP signalling and transcription factors. J. Biochem. 2012, 151, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, E.M.; Frost, R.J.; Olson, E.N. MicroRNAs add a new dimension to cardiovascular disease. Circulation 2010, 121, 1022–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Liu, X.; Zhang, X.; Song, Z.; Han, L.; He, Y.; Xu, Z. MicroRNA-30b is a multifunctional regulator of aortic valve interstitial cells. J. Thorac. Cardiovasc. Surg. 2014, 147, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Nigam, V.; Sievers, H.H.; Jensen, B.C.; Sier, H.A.; Simpson, P.C.; Srivastava, D.; Mohamed, S.A. Altered microRNAs in bicuspid aortic valve: A comparison between stenotic and insufficient valves. J. Heart Valve Dis. 2010, 19, 459–465. [Google Scholar]
- Petrkova, J.; Borucka, J.; Kalab, M.; Klevcova, P.; Michalek, J.; Taborsky, M.; Petrek, M. Increased expression of miR-146a in valvular tissue from patients with aortic valve stenosis. Front Cardiovasc Med 2019, 6, 86. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, S.; Deng, C.; Li, F.; Wang, Y.; Hu, X.; Shi, F.; Dong, N. MicroRNA-204 targets runx2 to attenuate BMP-2-induced osteoblast differentiation of human aortic valve interstitial cells. J. Cardiovasc. Pharmacol. 2015, 66, 63–71. [Google Scholar] [CrossRef]
- Toshima, T.; Watanabe, T.; Narumi, T.; Otaki, Y.; Shishido, T.; Aono, T.; Goto, J.; Watanabe, K.; Sugai, T.; Takahashi, T. Therapeutic inhibition of microRNA-34a ameliorates aortic valve calcification via modulation of Notch1-Runx2 signalling. Cardiovasc. Res. 2019. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, L.; Xing, L.; Chen, D. MicroRNA-204 regulates Runx2 protein expression and mesenchymal progenitor cell differentiation. Stem Cells 2010, 28, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, R.; Fullerton, D.A.; Ao, L.; Zhao, K.s.; Reece, T.B.; Cleveland, J.C.; Meng, X. Altered microRNA expression is responsible for the pro-osteogenic phenotype of interstitial cells in calcified human aortic valves. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Miron, R.; Zhang, Y. Osteoinduction: A review of old concepts with new standards. J. Dent. Res. 2012, 91, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Foster, L.J.; Zeemann, P.A.; Li, C.; Mann, M.; Jensen, O.N.; Kassem, M. Differential expression profiling of membrane proteins by quantitative proteomics in a human mesenchymal stem cell line undergoing osteoblast differentiation. Stem Cells 2005, 23, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Benton, J.A.; Kern, H.B.; Anseth, K.S. Substrate properties influence calcification in valvular interstitial cell culture. J. Heart Valve Dis. 2008, 17, 689–699. [Google Scholar] [PubMed]
- Clark-Greuel, J.N.; Connolly, J.M.; Sorichillo, E.; Narula, N.R.; Rapoport, H.S.; Mohler, E.R., III; Gorman, J.H., III; Gorman, R.C.; Levy, R.J. Transforming growth factor-beta1 mechanisms in aortic valve calcification: Increased alkaline phosphatase and related events. Ann. Thorac. Surg. 2007, 83, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Fullerton, D.A.; Su, X.; Ao, L.; Cleveland, J.C.; Meng, X. Pro-osteogenic phenotype of human aortic valve interstitial cells is associated with higher levels of Toll-like receptors 2 and 4 and enhanced expression of bone morphogenetic protein 2. J. Am. Coll. Cardiol. 2009, 53, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Ducy, P. Cbfa1: A molecular switch in osteoblast biology. Dev. Dyn. 2000, 219, 461–471. [Google Scholar] [CrossRef]
- Yoon, D.S.; Kim, Y.H.; Lee, S.; Lee, K.M.; Park, K.H.; Jang, Y.; Lee, J.W. Interleukin-6 induces the lineage commitment of bone marrow-derived mesenchymal multipotent cells through down-regulation of Sox2 by osteogenic transcription factors. FASEB J. 2014, 28, 3273–3286. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, Y.; Fu, Q.; Dong, Y. Osterix is required for sonic hedgehog-induced osteoblastic MC3T3-E1 cell differentiation. Cell Biochem. Biophys. 2012, 64, 169–176. [Google Scholar] [CrossRef]
- Meng, X.; Ao, L.; Song, Y.; Babu, A.; Yang, X.; Wang, M.; Weyant, M.J.; Dinarello, C.A.; Cleveland, J.C., Jr.; Fullerton, D.A. Expression of functional Toll-like receptors 2 and 4 in human aortic valve interstitial cells: Potential roles in aortic valve inflammation and stenosis. Am. J. Physiol. Cell Physiol. 2008, 294, C29–C35. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Jin, C.; Ao, L.; Cleveland, J.C., Jr.; Song, R.; Xu, D.; Fullerton, D.A.; Meng, X. Cross-talk between the toll-like receptor 4 and notch1 pathways augments the inflammatory response in the interstitial cells of stenotic human aortic valves. Circulation 2012, 126 (Suppl. 1), S222–S230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, J.D.; Levay, A.K.; Gillaspie, D.B.; Tao, G.; Lincoln, J. Reduced sox9 function promotes heart valve calcification phenotypes in vivo. Circ. Res. 2010, 106, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ao, L.; Zou, N.; Cleveland, J.C.J.; Fullerton, D.A.; Meng, X. Myocardial TLR4 is a determinant of neutrophil infiltration after global myocardial ischemia: Mediating KC and MCP-1 expression induced by extracellular HSC70. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H21–H28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Meng, X.; Brown, J.M.; Ao, L.; Banerjee, A.; Harken, A.H. Norepinephrine induces cardiac heat shock protein and delayed cardioprotection in the rat through a1-adrenoceptors. Cardiovasc. Res. 1996, 32, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Song, R.; Zeng, Q.; Ao, L.; Jessica, A.Y.; Cleveland, J.C.; Zhao, K.-s.; Fullerton, D.A.; Meng, X. Biglycan induces the expression of osteogenic factors in human aortic valve interstitial cells via Toll-like receptor-2. Atertio. Thromb. Vasc. Biol. 2012, 32, 2711–2720. [Google Scholar] [CrossRef] [Green Version]
- Cowan, C.M.; Zhang, X.; James, A.W.; Mari Kim, T.; Sun, N.; Wu, B.; Ting, K.; Soo, C. NELL-1 increases pre-osteoblast mineralization using both phosphate transporter Pit1 and Pit2. Biochem. Biophys. Res. Commun. 2012, 422, 351–357. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, R.; Zhai, Y.; Ao, L.; Fullerton, D.A.; Meng, X. MicroRNA-204 Deficiency in Human Aortic Valves Elevates Valvular Osteogenic Activity. Int. J. Mol. Sci. 2020, 21, 76. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010076
Song R, Zhai Y, Ao L, Fullerton DA, Meng X. MicroRNA-204 Deficiency in Human Aortic Valves Elevates Valvular Osteogenic Activity. International Journal of Molecular Sciences. 2020; 21(1):76. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010076
Chicago/Turabian StyleSong, Rui, Yufeng Zhai, Lihua Ao, David A. Fullerton, and Xianzhong Meng. 2020. "MicroRNA-204 Deficiency in Human Aortic Valves Elevates Valvular Osteogenic Activity" International Journal of Molecular Sciences 21, no. 1: 76. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010076