Comparative Effects and Mechanisms of Chitosan and Its Derivatives on Hypercholesterolemia in High-Fat Diet-Fed Rats


Abstract
:1. Introduction
2. Results and Discussion
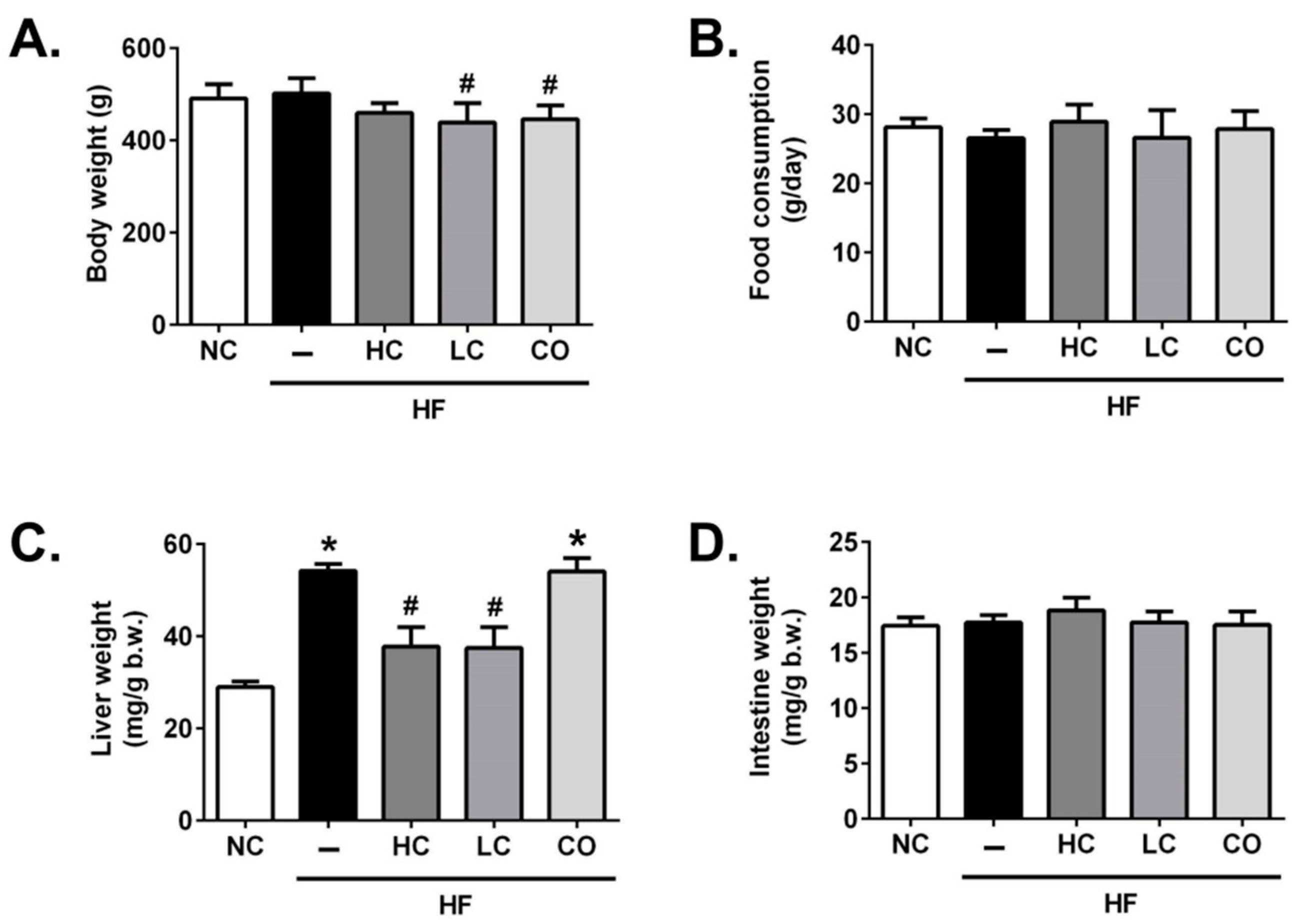
2.1. Effects of High Molecular Weight Chitosan (HC), Low Molecular Weight Chitosan (LC), and Chitosan Oligosaccharide (CO) on Body Weight, Organ Weights, and Food Intake in High-Fat (HF) Diet-Fed Rats
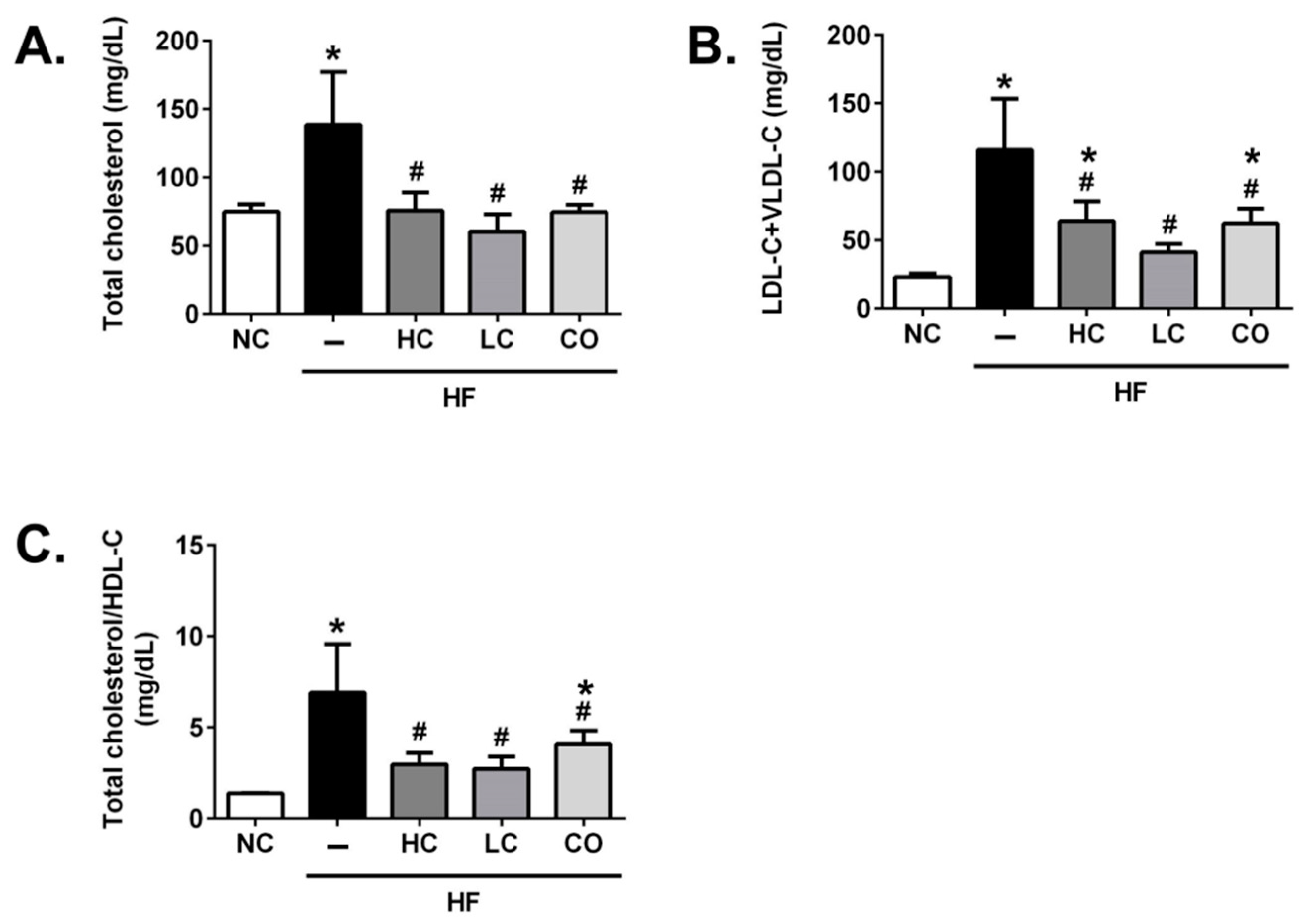
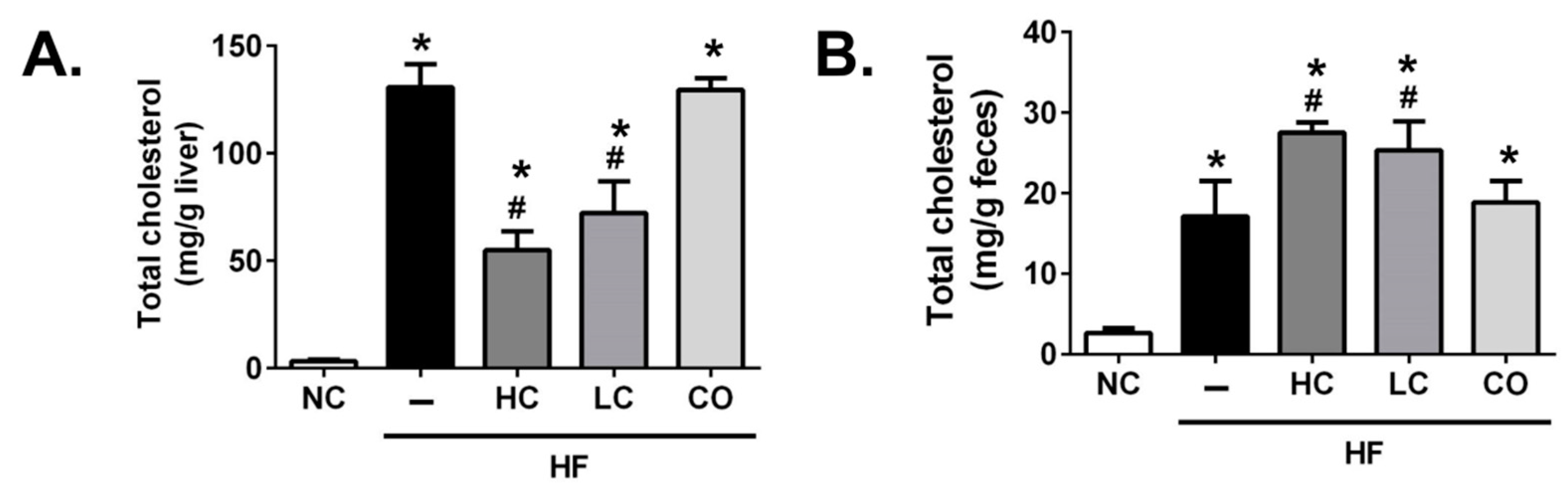
2.2. Effects of HC, LC, and CO on Hepatic and Fecal Lipid Responses in HF Diet-Fed Rats
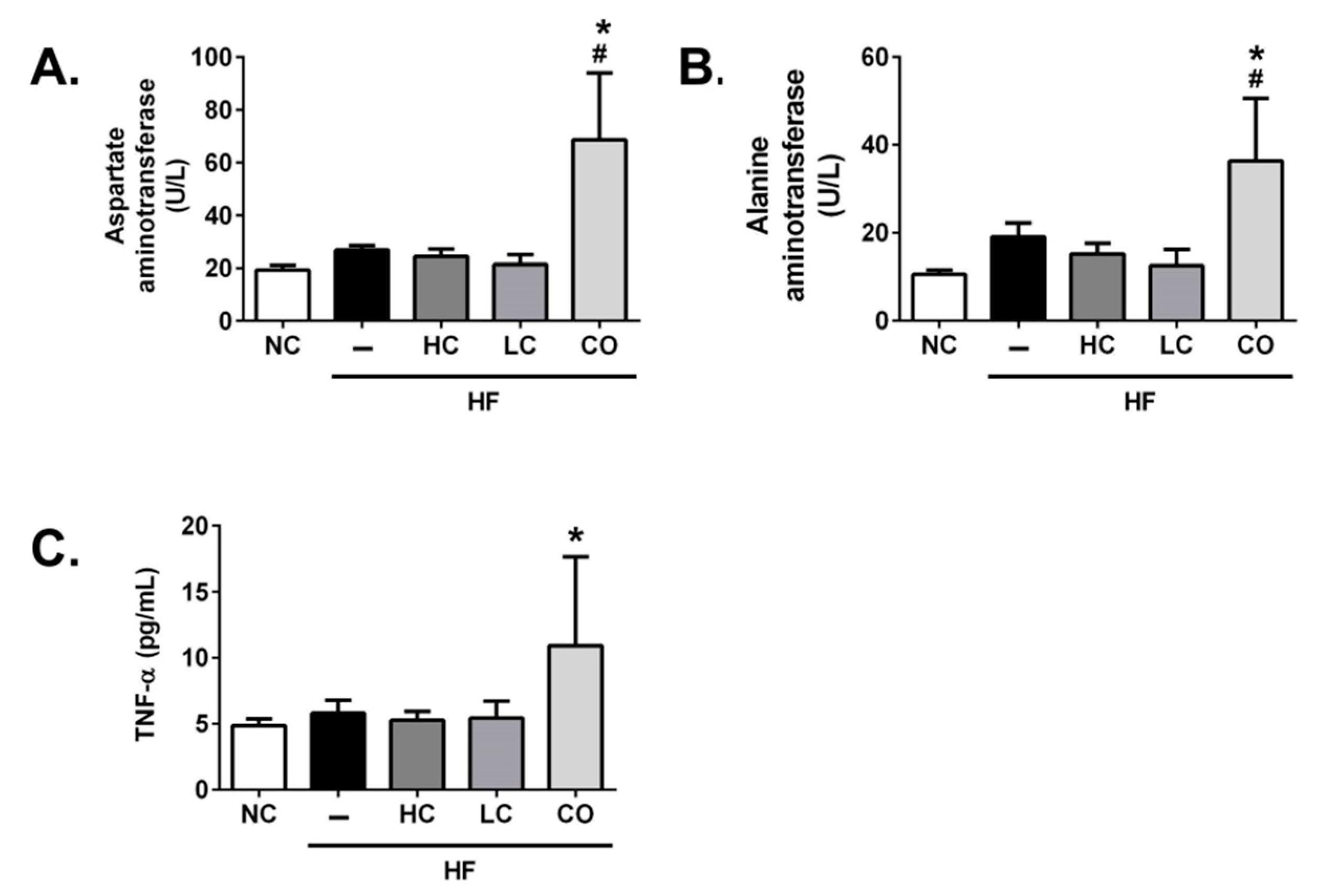
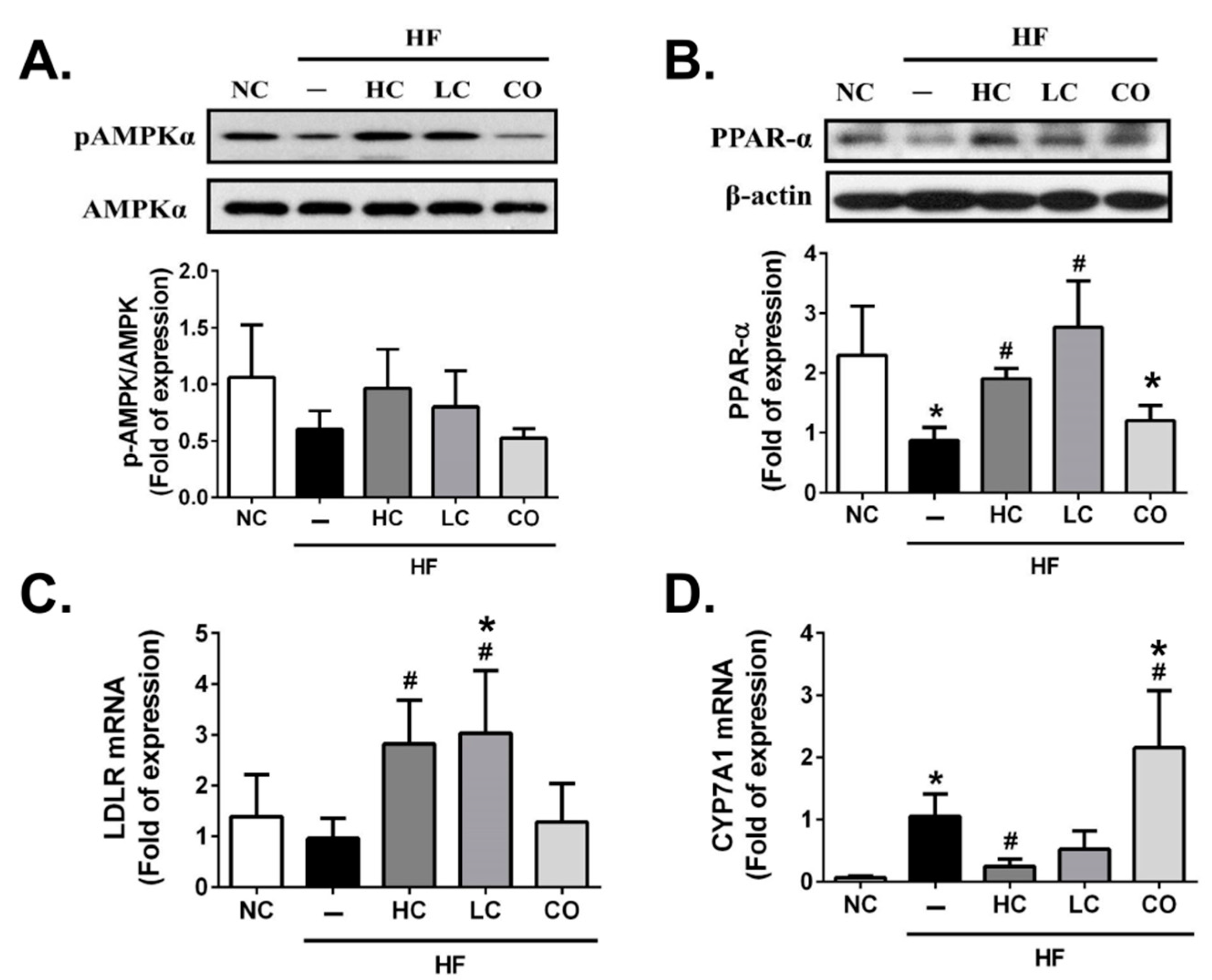
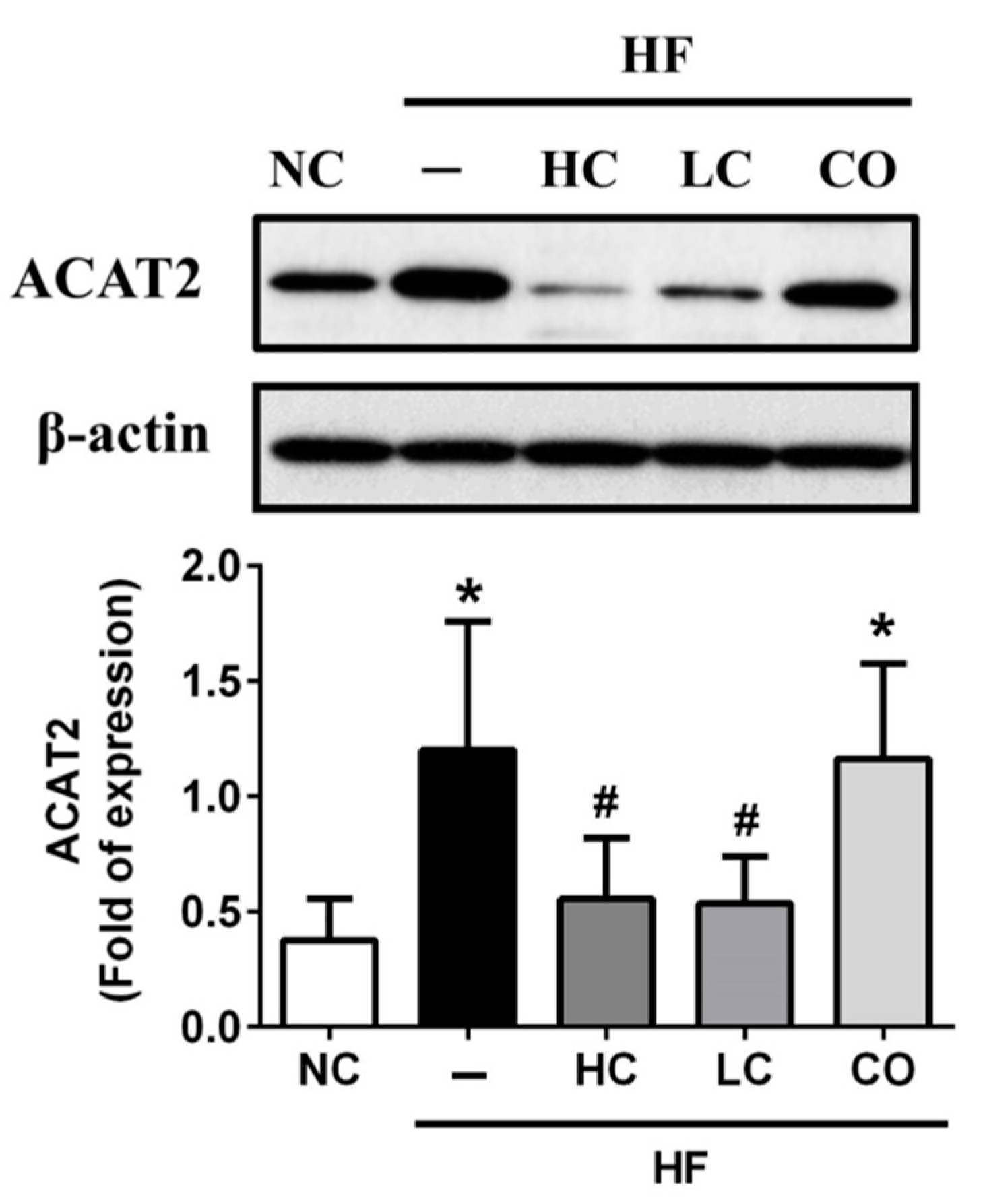
2.3. The Mechanism of HC, LC, and CO in the Livers and Intestines of HF Diet-Fed Rats
3. Materials and Methods
3.1. Materials
3.2. Animals and Diets
3.3. Determination of Total Cholesterol (TC) and Plasma Lipoprotein Cholesterol
3.4. Determination of Activities of Aspartate Aminotransferase (AST) and Alanine Aminotransferase (ALT)
3.5. Determination of Plasma Tumor Necrosis Factor-α (TNF-α)
3.6. Western Blot Analysis
3.7. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
3.8. Statistical Evaluation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Du, S.; Wang, C.; Jiang, W.; Li, C.; Li, Y.; Feng, R.; Sun, C. The impact of body weight gain on nonalcoholic fatty liver disease and metabolic syndrome during earlier and later adulthood. Diabetes Res. Clin. Pract. 2016, 116, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.N.; Du, S.S.; Wang, C.; Li, Y.C.; Liu, L.Y.; Guo, F.C.; Sun, C.H. Lean-non-alcoholic fatty liver disease increases risk for metabolic disorders in a normal weight Chinese population. World J. Gastroenterol. 2014, 20, 17932–17940. [Google Scholar] [CrossRef] [PubMed]
- Targher, G.; Marra, F.; Marchesini, G. Increased risk of cardiovascular disease in non-alcoholic fatty liver disease: Causal effect or epiphenomenon? Diabetologia 2008, 51, 1947–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nseir, W.; Hellou, E.; Assy, N. Role of diet and lifestyle changes in nonalcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 9338–9344. [Google Scholar] [PubMed]
- Choi, J.H.; Rhee, E.J.; Bae, J.C.; Park, S.E.; Park, C.Y.; Cho, Y.K.; Oh, K.W.; Park, S.W.; Lee, W.Y. Increased risk of type 2 diabetes in subjects with both elevated liver enzymes and ultrasonographically diagnosed nonalcoholic fatty liver disease: A 4-year longitudinal study. Arch. Med. Res. 2013, 44, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Bunnoy, A.; Saenphet, K.; Lumyong, S.; Saenphet, S.; Chomdej, S. Monascus purpureus-fermented Thai glutinous rice reduces blood and hepatic cholesterol and hepatic steatosis concentrations in diet-induced hypercholesterolemic rats. BMC Complement. Altern. Med. 2015, 15, 88. [Google Scholar] [CrossRef] [Green Version]
- Senthilkumar, K.; Kim, S.K. Marine invertebrate natural products for anti-inflammatory and chronic diseases. Evid. Based Complement. Altern. Med. 2013, 2013, 572859. [Google Scholar] [CrossRef]
- Xia, W.; Liu, P.; Zhang, J.; Chen, J. Biological activities of chitosan and chitooligosaccharides. Food Hydrocoll. 2011, 25, 170–179. [Google Scholar] [CrossRef]
- Anitha, A.; Sowmya, S.; Sudheesh Kumar, P.T.; Deepthi, S.; Chennazhi, K.P.; Ehrlich, H.; Tsurkan, M.; Jayakumar, R. Chitin and chitosan in selected biomedical applications. Prog. Polym. Sci. 2014, 39, 1644–1667. [Google Scholar] [CrossRef]
- Laurienzo, P. Marine polysaccharides in pharmaceutical applications: An overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef] [Green Version]
- Kerch, G. The potential of chitosan and its derivatives in prevention and treatment of age-related diseases. Mar. Drugs 2015, 13, 2158–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Chen, J.; Cao, P.; Pan, H.; Ding, C.; Xiao, T.; Zhang, P.; Guo, J.; Su, Z. Anti-obese effect of glucosamine and chitosan oligosaccharide in high-fat diet-induced obese rats. Mar. Drugs 2015, 13, 2732–2756. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, M.; Kimura, Y. Low molecular weight chitosan inhibits obesity induced by feeding a high-fat diet long-term in mice. J. Pharm. Pharmacol. 2006, 58, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Picchi, M.G.; Mattos, A.M.; Barbosa, M.R.; Duarte, C.P.; Gandini Mde, A.; Portari, G.V.; Jordão, A.A. A high-fat diet as a model of fatty liver disease in rats. Acta Cir. Bras. 2011, 26 (Suppl. 2), 25–30. [Google Scholar] [CrossRef] [Green Version]
- Teodoro, J.S.; Gomes, A.P.; Varela, A.T.; Duarte, F.V.; Rolo, A.P.; Palmeira, C.M. Hepatic and skeletal muscle mitochondrial toxicity of chitosan oligosaccharides of normal and diabetic rats. Toxicol. Mech. Methods 2016, 26, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.T.; Huang, S.Y.; Chiang, M.T. A comparative study on hypoglycemic and hypocholesterolemic effects of high and low molecular weight chitosan in streptozotocin-induced diabetic rats. Food Chem. Toxicol. 2008, 46, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Yang, Q.; Huang, G.; Ding, C.; Cao, P.; Huang, L.; Xiao, T.; Guo, J.; Su, Z. Hypolipidemic effects of chitosan and its derivatives in hyperlipidemic rats induced by a high-fat diet. Food Nutr. Res. 2016, 60, 31137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakino, S.; Ohki, T.; Nakayama, H.; Yuan, X.; Otabe, S.; Hashinaga, T.; Wada, N.; Kurita, Y.; Tanaka, K.; Hara, K.; et al. Pivotal Role of TNF-α in the Development and Progression of Nonalcoholic Fatty Liver Disease in a Murine Model. Horm. Metab. Res. 2018, 50, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.H.; Yang, H.P.; Chun, H.S. Chitooligosaccharide ameliorates diet-induced obesity in mice and affects adipose gene expression involved in adipogenesis and inflammation. Nutr. Res. 2012, 32, 218–228. [Google Scholar] [CrossRef]
- Yao, H.T.; Luo, M.N.; Li, C.C. Chitosan oligosaccharides reduce acetaminophen-induced hepatotoxicity by suppressing CYP-mediated bioactivation. J. Funct. Foods 2015, 12, 262–270. [Google Scholar] [CrossRef]
- Loh, K.; Tam, S.; Murray-Segal, L.; Huynh, K.; Meikle, P.J.; Scott, J.W.; van Denderen, B.; Chen, Z.; Steel, R.; LeBlond, N.D.; et al. Inhibition of Adenosine Monophosphate-Activated Protein Kinase-3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase Signaling Leads to Hypercholesterolemia and Promotes Hepatic Steatosis and Insulin Resistance. Hepatol. Commun. 2018, 3, 84–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khera, A.V.; Millar, J.S.; Ruotolo, G.; Wang, M.D.; Rader, D.J. Potent peroxisome proliferator-activated receptor-α agonist treatment increases cholesterol efflux capacity in humans with the metabolic syndrome. Eur. Heart J. 2015, 36, 3020–3022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemper, M.F.; Srivastava, S.; Todd King, M.; Clarke, K.; Veech, R.L.; Pawlosky, R.J. An ester of β-Hydroxybutyrate regulates cholesterol biosynthesis in rats and a cholesterol biomarker in humans. Lipids 2015, 50, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Levinson, S.S.; Wagner, S.G. Implications of reverse cholesterol transport: Recent studies. Clin. Chim. Acta 2015, 439, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Bieghs, V.; Van Gorp, P.J.; Wouters, K.; Hendrikx, T.; Gijbels, M.J.; van Bilsen, M.; Bakker, J.; Binder, C.J.; Lütjohann, D.; Staels, B.; et al. LDL receptor knock-out mice are a physiological model particularly vulnerable to study the onset of inflammation in non-alcoholic fatty liver disease. PLoS ONE 2012, 7, e30668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, M.H.; Houng, S.J.; Jun, H.J.; Lee, J.H.; Choi, J.W.; Kim, S.H.; Kim, Y.R.; Lee, S.J. Barley intake induces bile acid excretion by reduced expression of intestinal ASBT and NPC1L1 in C57BL/6J mice. J. Agric. Food Chem. 2011, 59, 6798–6805. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Cassader, M. Cholesterol metabolism and the pathogenesis of non-alcoholic steatohepatitis. Prog. Lipid Res. 2013, 52, 175–191. [Google Scholar] [CrossRef] [Green Version]
- Ferrell, J.M.; Boehme, S.; Li, F.; Chiang, J.Y. Cholesterol 7α-hydroxylase-deficient mice are protected from high-fat/high-cholesterol diet-induced metabolic disorders. J. Lipid Res. 2016, 57, 1144–1154. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.G.; Willingham, M.C.; Davis, M.A.; Skinner, K.A.; Rudel, L.L. Differential expression of ACAT1 and ACAT2 among cells within liver, intestine, kidney, and adrenal of nonhuman primates. J. Lipid Res. 2000, 41, 1991–2001. [Google Scholar]
- Zhou, X.; Ren, F.; Wei, H.; Liu, L.; Shen, T.; Xu, S.; Wei, J.; Ren, J.; Ni, H. Combination of berberine and evodiamine inhibits intestinal cholesterol absorption in high fat diet induced hyperlipidemic rats. Lipids Health Dis. 2017, 16, 239. [Google Scholar] [CrossRef] [Green Version]
- Institute of Laboratory Animal Resources. Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 2011. [Google Scholar]
- Takehisa, F.; Suzuki, Y. Effect of guar gum and cholestyramine on plasma lipoprotein cholesterol in rats. J. Jap. Soc. Nutr. Food Sci. 1990, 43, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.Y.; Wang, L.P.; Liu, S.H.; Chiang, M.T. Fish Oil Supplementation Alleviates the Altered Lipid Homeostasis in Blood, Liver, and Adipose Tissues in High-Fat Diet-Fed Rats. J. Agric. Food Chem. 2018, 66, 4118–4128. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Chang, T.C.; Liu, S.H.; Chiang, M.T. The regulatory effects of fish oil and chitosan on hepatic lipogenic signals in high-fat diet-induced obese rats. J. Food Drug Anal. 2017, 25, 919–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (%) | NC | HF | HC | LC | CO |
|---|---|---|---|---|---|
| Corn starch | 63.8 | 56.2 | 56.2 | 56.2 | 56.2 |
| Casein | 20 | 20 | 20 | 20 | 20 |
| Lard | 3 | 10 | 10 | 10 | 10 |
| Soybean oil | 2 | 2 | 2 | 2 | 2 |
| Vitamin 1 | 1 | 1 | 1 | 1 | 1 |
| Mineral 2 | 5 | 5 | 5 | 5 | 5 |
| Cholesterol | 0.5 | 0.5 | 0.5 | 0.5 | |
| Cholic acid | 0.1 | 0.1 | 0.1 | 0.1 | |
| Choline chloride | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Cellulose | 5 | 5 | |||
| High-MW chitosan | 5 | ||||
| Low-MW chitosan | 5 | ||||
| Chitooligosaccharide | 5 | ||||
| NC: Normal control diet | |||||
| HF: HF diet | |||||
| HC: HF diet + 5% high-MW chitosan | |||||
| LC: HF diet + 5% low-MW chitosan | |||||
| CO: HF diet + 5% chitooligosaccharide | |||||
| DD: Degrees of deacetylation |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-Y.; Yen, T.-E.; Liu, S.-H.; Chiang, M.-T. Comparative Effects and Mechanisms of Chitosan and Its Derivatives on Hypercholesterolemia in High-Fat Diet-Fed Rats. Int. J. Mol. Sci. 2020, 21, 92. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010092
Chiu C-Y, Yen T-E, Liu S-H, Chiang M-T. Comparative Effects and Mechanisms of Chitosan and Its Derivatives on Hypercholesterolemia in High-Fat Diet-Fed Rats. International Journal of Molecular Sciences. 2020; 21(1):92. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010092
Chicago/Turabian StyleChiu, Chen-Yuan, Tsai-En Yen, Shing-Hwa Liu, and Meng-Tsan Chiang. 2020. "Comparative Effects and Mechanisms of Chitosan and Its Derivatives on Hypercholesterolemia in High-Fat Diet-Fed Rats" International Journal of Molecular Sciences 21, no. 1: 92. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010092