Targeting NF-κB by the Cell-Permeable NEMO-Binding Domain Peptide Improves Albuminuria and Renal Lesions in an Experimental Model of Type 2 Diabetic Nephropathy

 ,
,
Abstract
:1. Introduction
2. Results
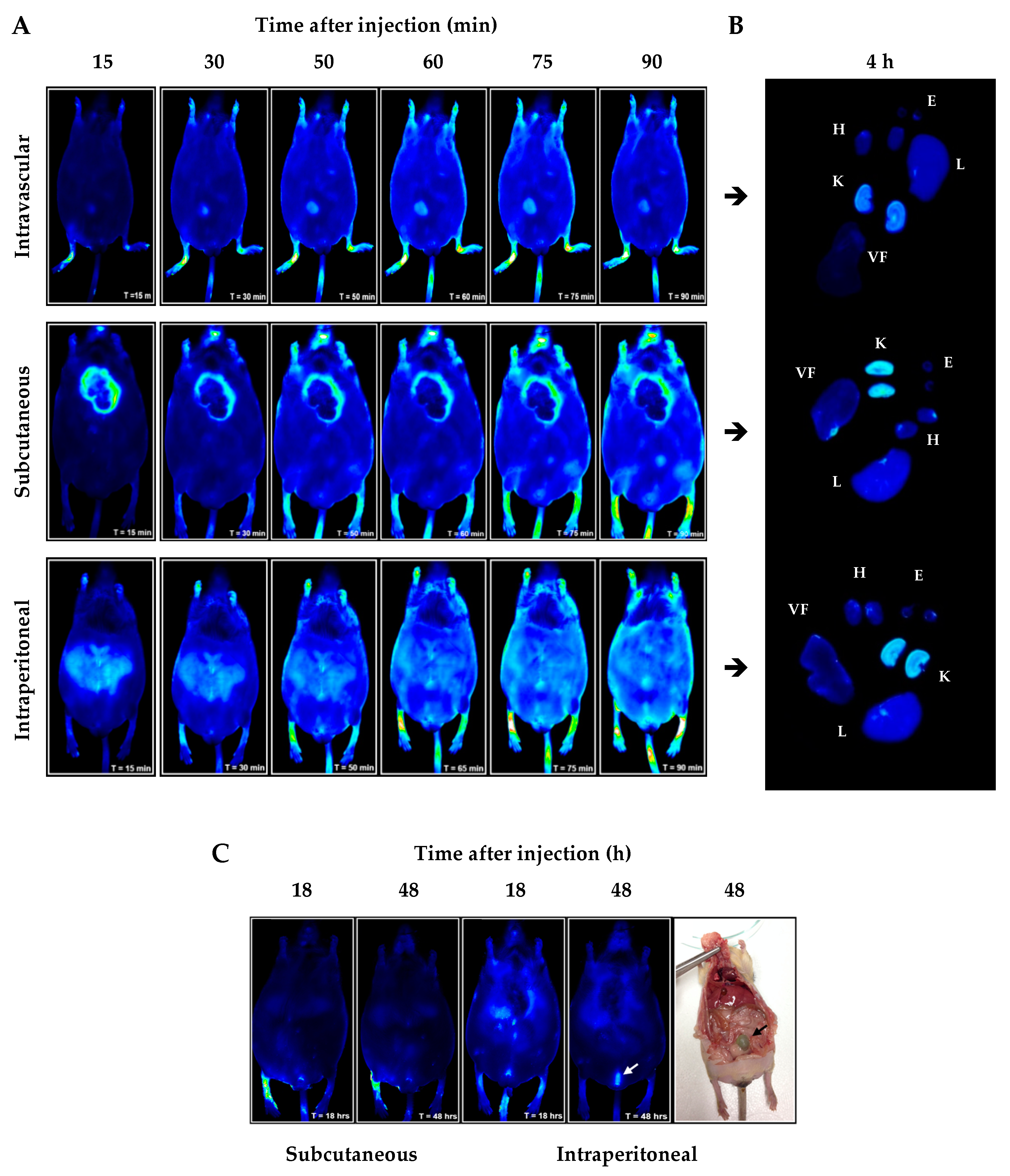
2.1. In Vivo/Ex Vivo Biodistribution of NBD Peptide in BTBR Ob/Ob Mice
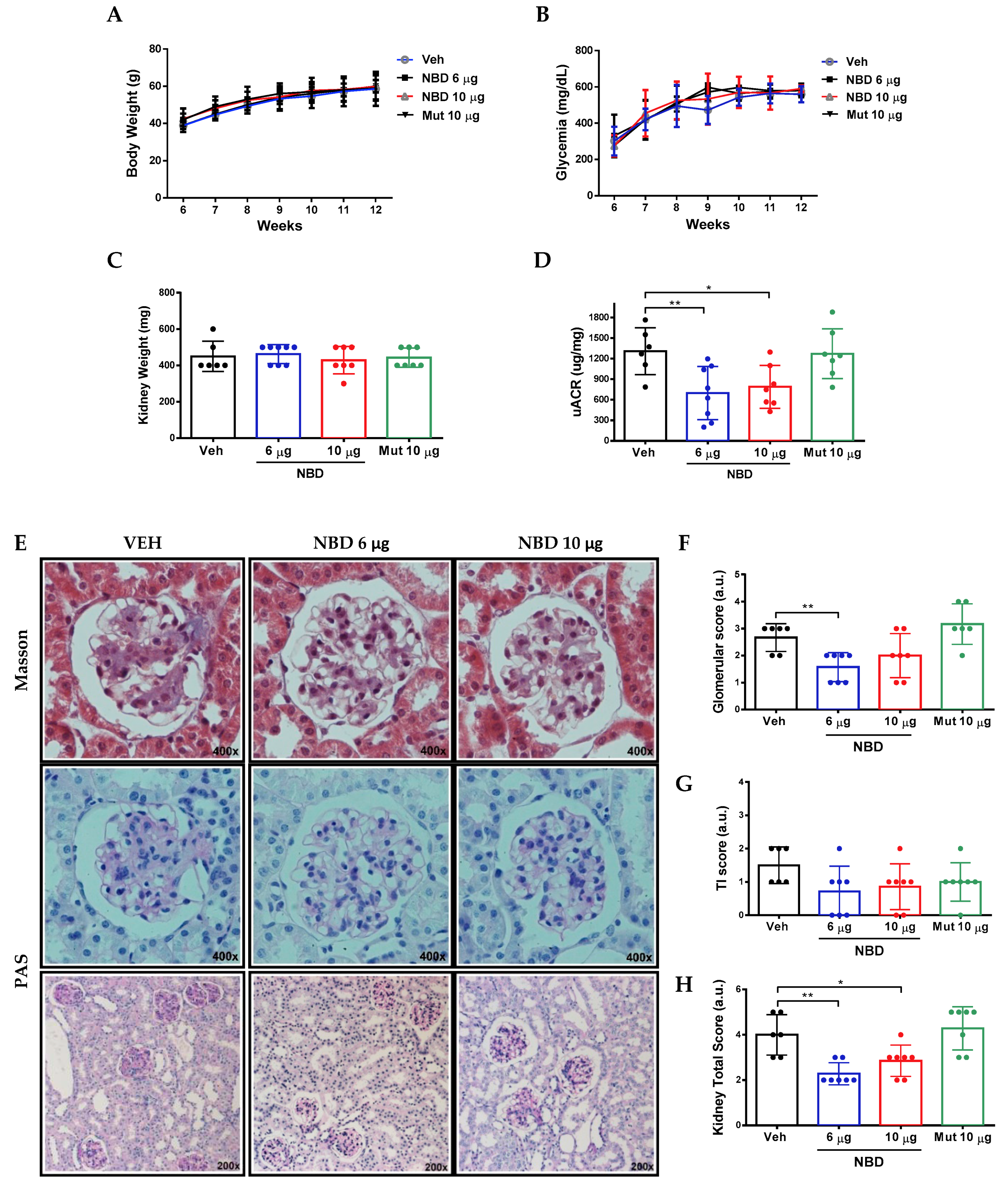
2.2. Cell-Permeable NBD Peptide Reduced Albuminuria and Morphologic Kidney Lesions in BTBR Ob/Ob Mice
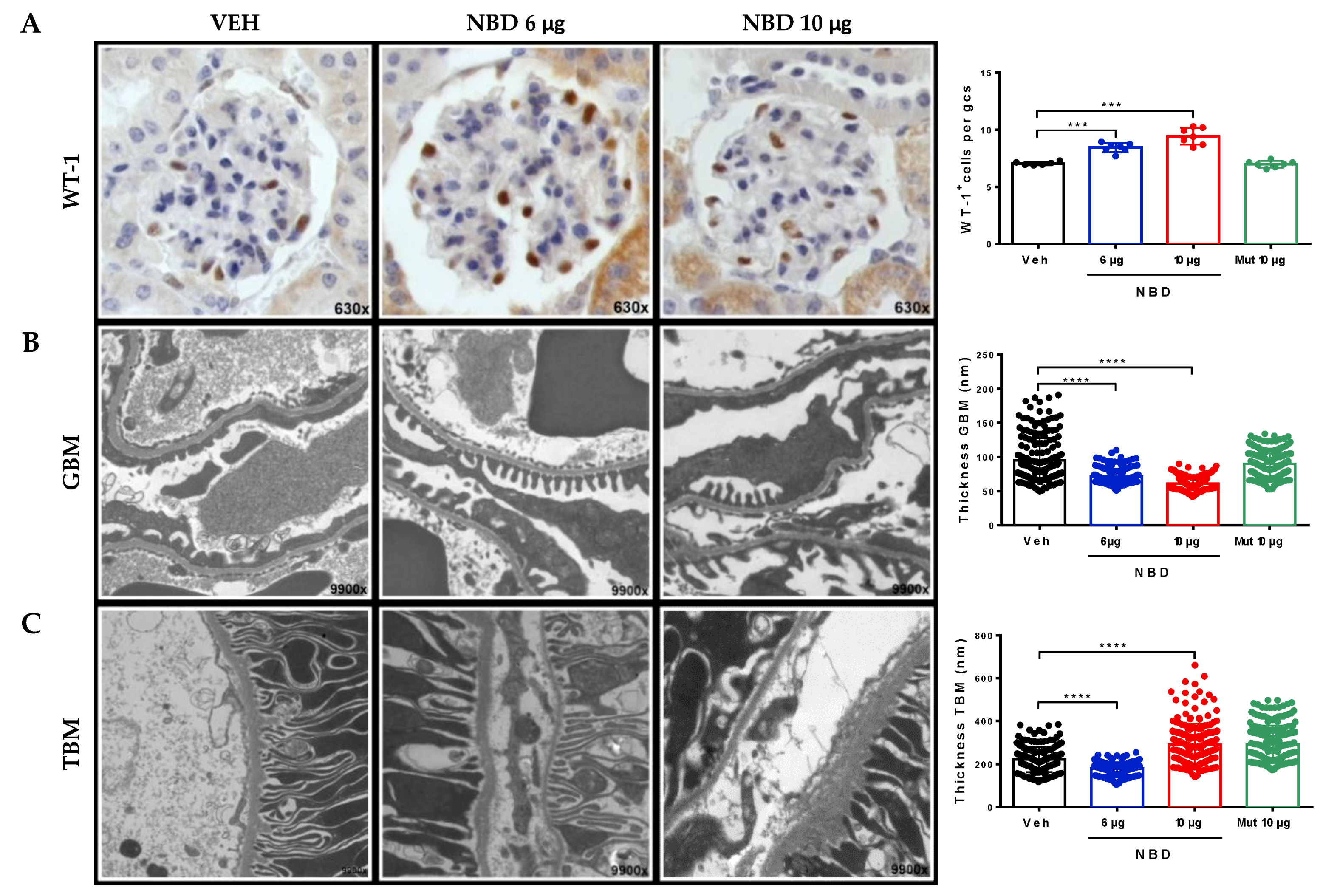
2.3. Cell-Permeable NBD Peptide Reduced Podocyte Damage and Basement Membrane Thickness in BTBR Ob/Ob Mouse Kidney
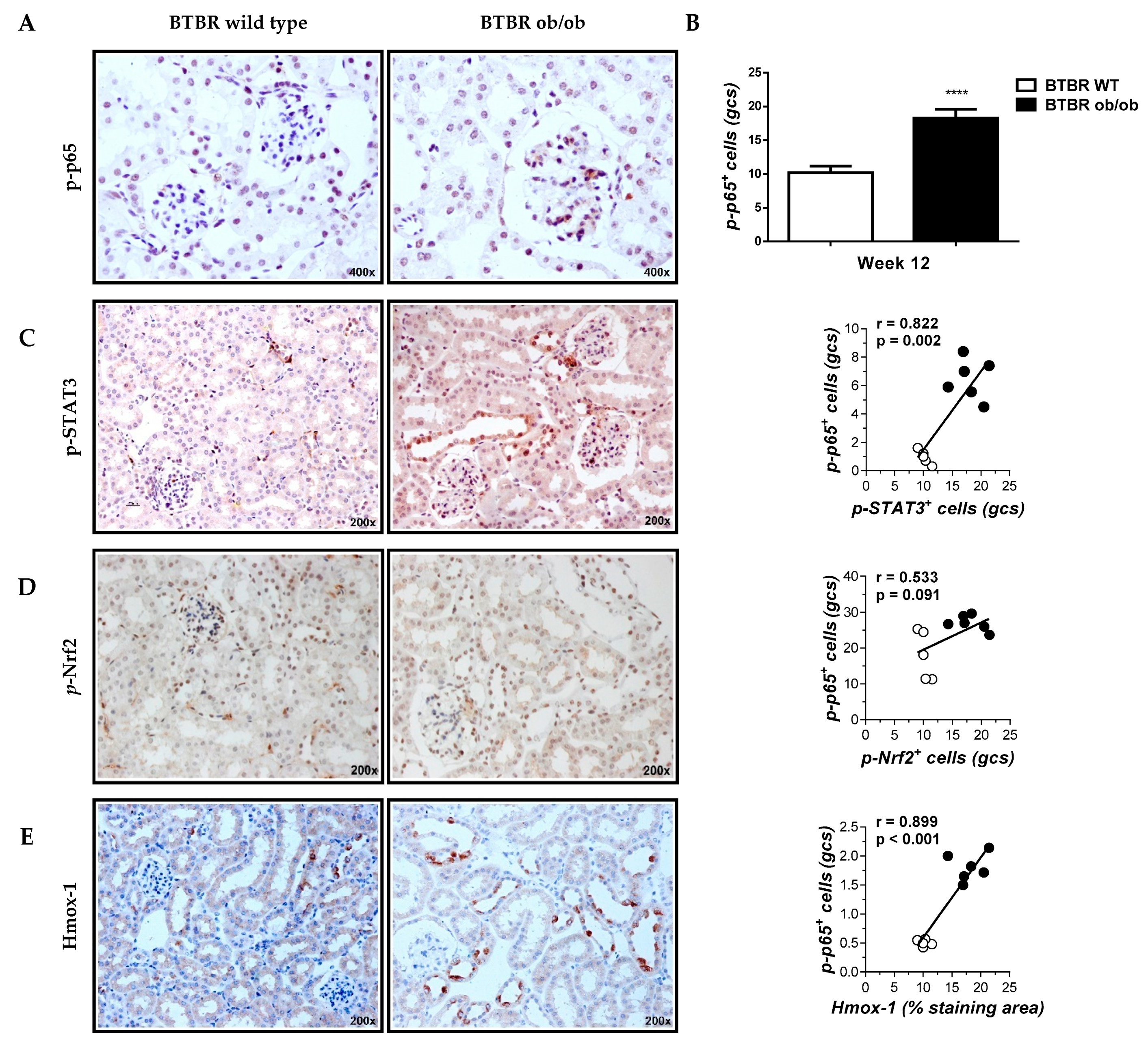
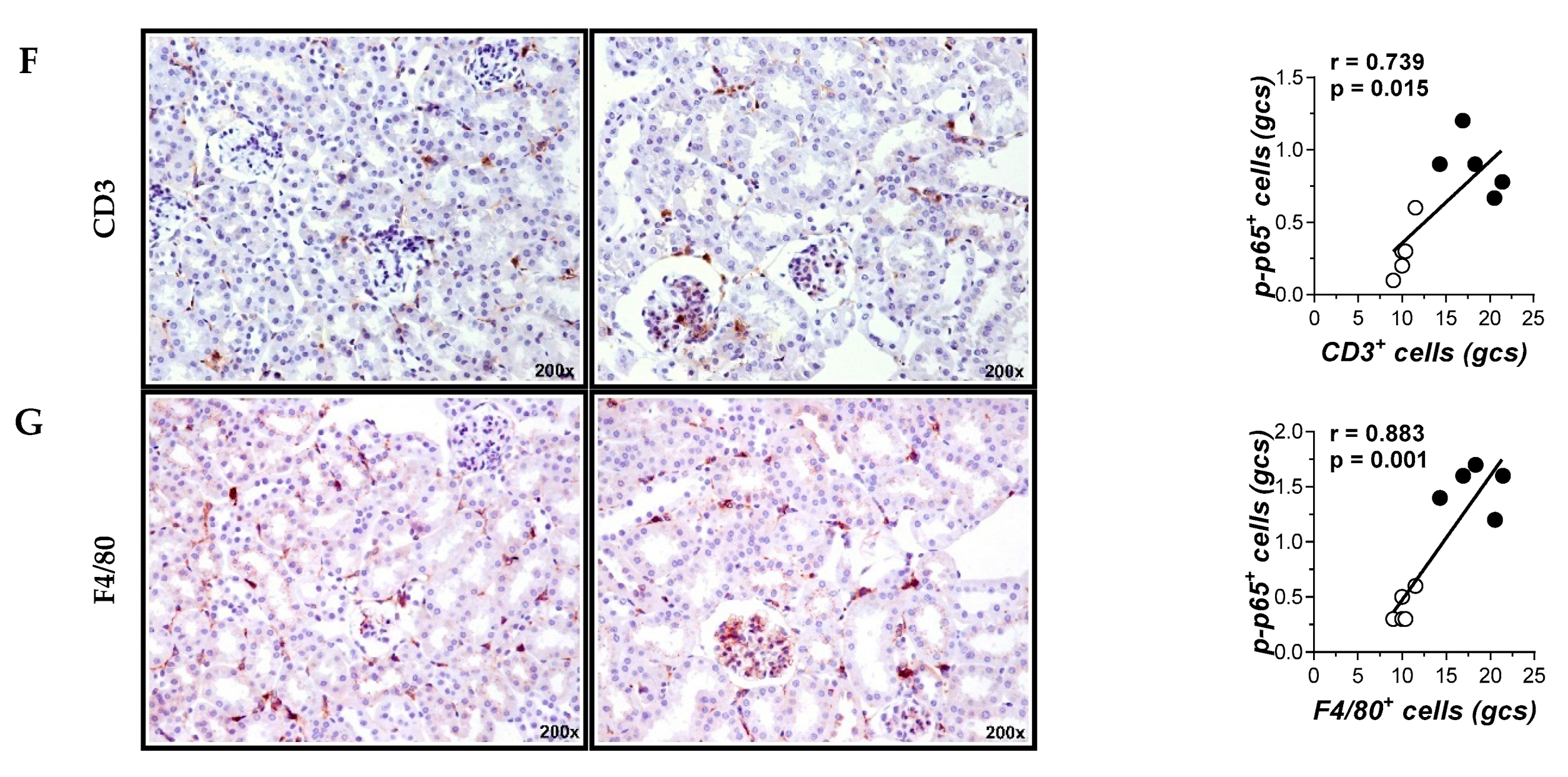
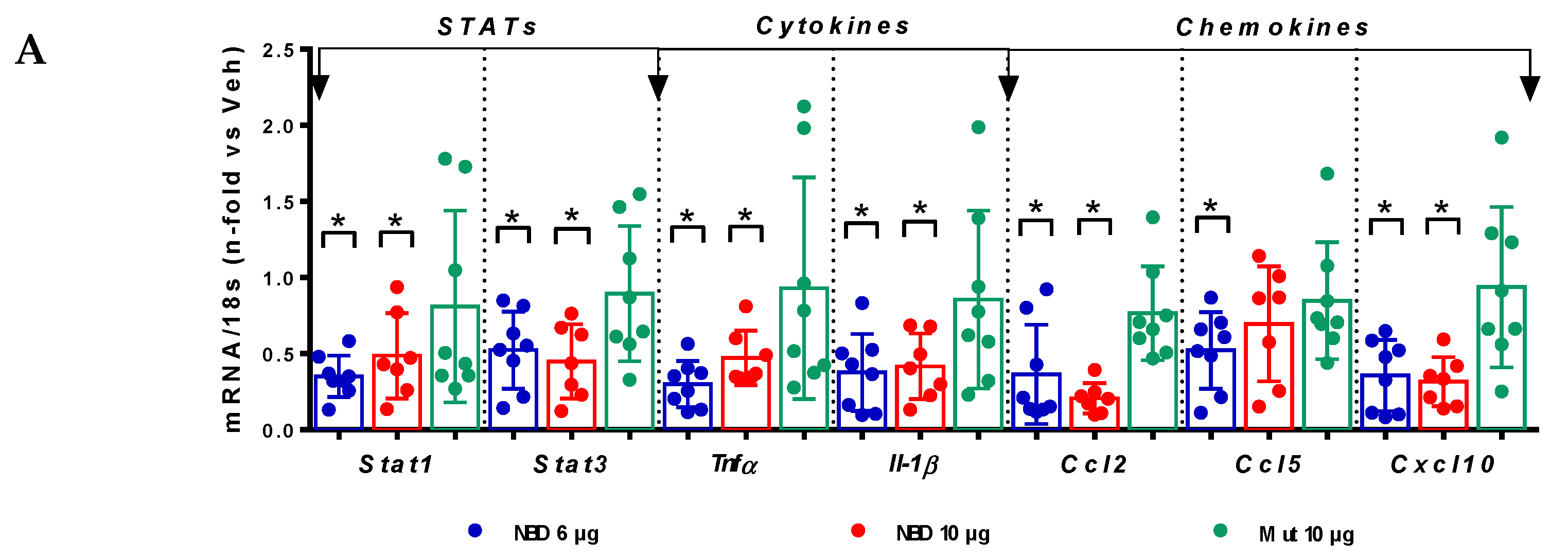
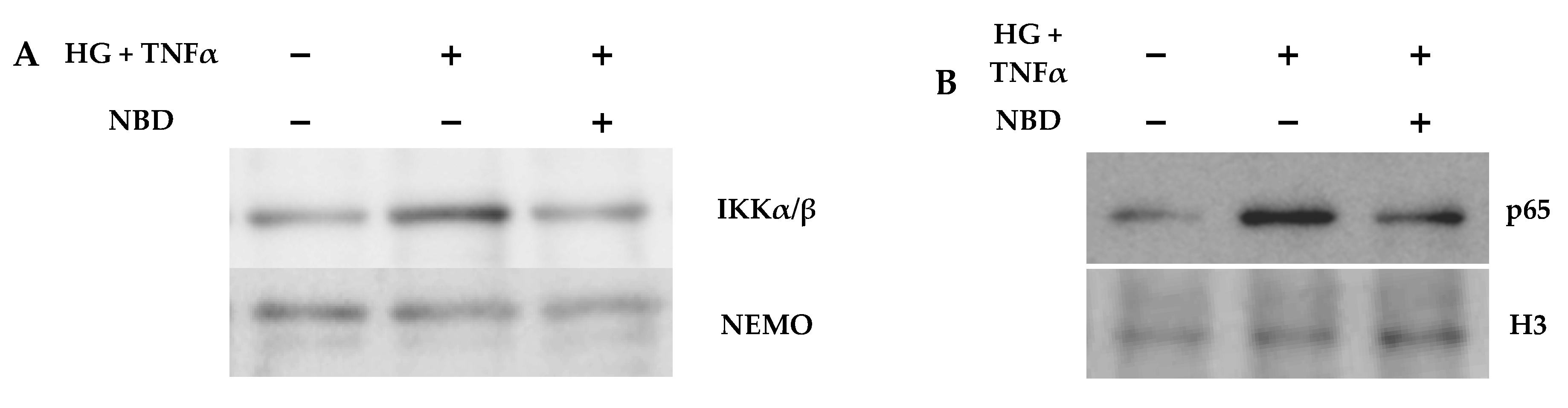
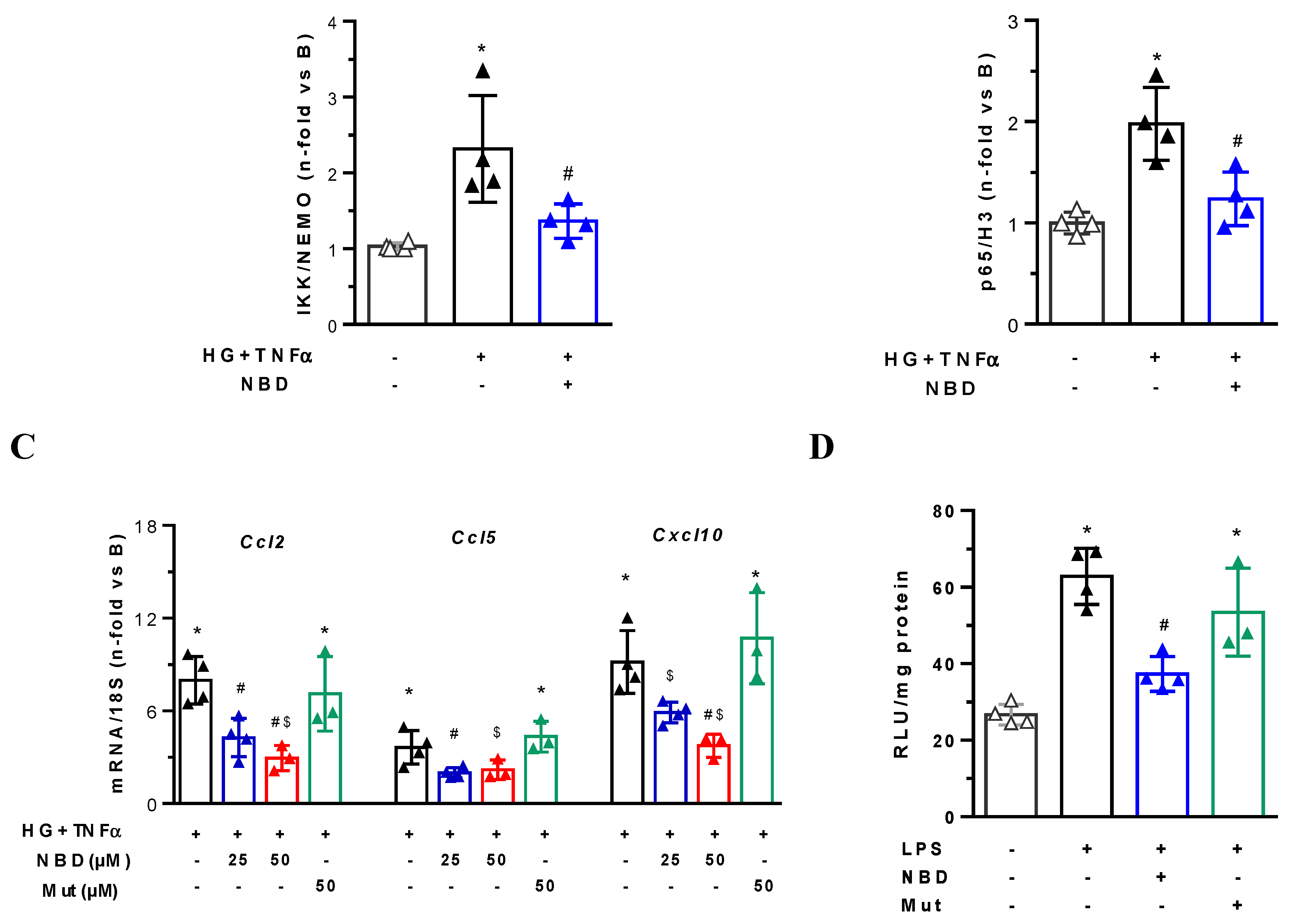
2.4. Cell-Permeable NBD Peptide Modulated the Proinflammatory and Oxidative Stress Markers in BTBR Ob/Ob Mice and Cultured Cells
3. Discussion
4. Materials and Methods
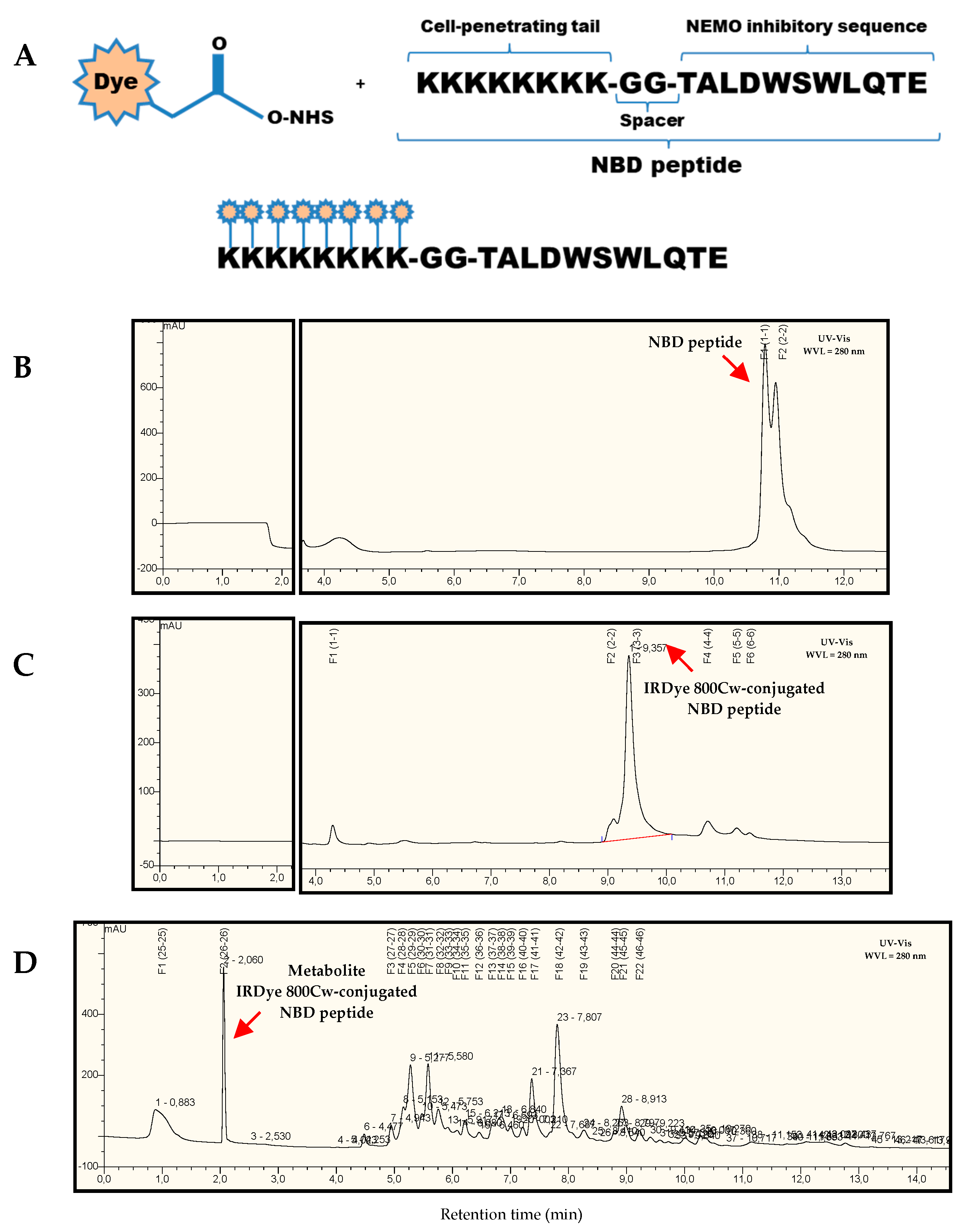
4.1. Synthesis of Cell-Permeable Peptides
4.2. In Vivo and Ex Vivo Optical Fluorescence Imaging
4.3. Histological Analysis and Immunohistochemistry
4.4. Cell Cultures
4.5. Western Blot and Immunoprecipitation
4.6. NOX Activity Assay
4.7. mRNA Expression
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| JAK | Janus Kinase 1 |
| STAT | Signal transducers and activators of transcription |
| TNFα | Tumor necrosis factor alpha |
| IL-1β | Interleukin 1β |
| CCL2/5 | C–C motif chemokine ligand 2/5 |
| CXCL10 | C–X–C motif chemokine 10 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| Hmox-1 | Heme-oxigenase-1 |
| NOX4 | NADPH oxidase 4 |
| SOD1 | Superoxide dismutase 1 |
| 18s | 18S ribosomal RNA |
References
- Unnikrishnan, R.; Pradeepa, R.; Joshi, S.R.; Mohan, V. Type 2 diabetes: Demystifying the global epidemic. Diabetes 2017, 66, 1432–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gassasse, Z.; Smith, D.; Finer, S.; Gallo, V. Association between urbanisation and type 2 diabetes: An ecological study. BMJ Glob. Heal. 2017, 2, e000473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Diabetes Federation. IDF Diabetes Atlas, 9th ed.; International Diabetes Federation: Brussels, Belgium, 2019. [Google Scholar]
- Satirapoj, B.; Adler, S.G. Comprehensive approach to diabetic nephropathy. Kidney Res. Clin. Pract. 2014, 33, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alicic, R.Z.; Rooney, M.T.; Tuttle, K.R. Diabetic kidney disease: Challenges, progress, and possibilities. Clin. J. Am. Soc. Nephrol. 2017, 12, 2032–2045. [Google Scholar] [CrossRef] [PubMed]
- Pichler, R.; Afkarian, M.; Dieter, B.P.; Tuttle, K.R. Immunity and inflammation in diabetic kidney disease: Translating mechanisms to biomarkers and treatment targets. Am. J. Physiol. Renal Physiol. 2017, 312, F716–F731. [Google Scholar] [CrossRef] [Green Version]
- Alicic, R.Z.; Johnson, E.J.; Tuttle, K.R. Inflammatory mechanisms as new biomarkers and therapeutic targets for diabetic kidney disease. Adv. Chronic Kidney Dis. 2018, 25, 181–191. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, B.; Ortiz, A.; Gomez-Guerrero, C.; Egido, J. Therapeutic approaches to diabetic nephropathy—Beyond the RAS. Nat. Rev. Nephrol. 2014, 10, 325–346. [Google Scholar] [CrossRef]
- Matoba, K.; Takeda, Y.; Nagai, Y.; Kawanami, D.; Utsunomiya, K.; Nishimura, R. Unraveling the role of inflammation in the pathogenesis of diabetic kidney disease. Int. J. Mol. Sci. 2019, 20, 3393. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.A.; Gomez-Guerrero, C.; Mas, S.; Sanz, A.B.; Lorenzo, O.; Ruiz-Ortega, M.; Opazo, L.; Mezzano, S.A.; Egido, J. Targeting inflammation in diabetic nephropathy: A tale of hope. Expert Opin. Investig. Drugs 2018, 27, 917–930. [Google Scholar] [CrossRef]
- Herman-Edelstein, M.; Scherzer, P.; Tobar, A.; Levi, M.; Gafter, U. Altered renal lipid metabolism and renal lipid accumulation in human diabetic nephropathy. J. Lipid Res. 2014, 55, 561. [Google Scholar] [CrossRef] [Green Version]
- Wada, J.; Hirofumi, M. Innate immunity in diabetes and diabetic nephropathy. Nat. Rev. Nephrol. 2016, 12, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Guijarro, C.; Egido, J. Transcription factor-kappa B (NF-kappa B) and renal disease. Kidney Int. 2001, 59, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, K.S.; Sethi, G.; Aggarwal, B.B. Nuclear factor-kappa B: From clone to clinic. Curr. Mol. Med. 2007, 7, 619–637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, S.C. NF-kB in inflammation and renal diseases. Cell Biosci. 2015, 5, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [Green Version]
- Sanz, A.B.; Sanchez-Niño, M.D.; Ramos, A.M.; Moreno, J.A.; Santamaria, B.; Ruiz-ortega, M.; Egido, J. NF-kappaB in Renal Inflammation. J. Am. Soc. Nephrol. 2010, 21, 1254–1262. [Google Scholar] [CrossRef] [Green Version]
- Mezzano, S.; Aros, C.; Droguett, A.; Burgos, M.E.; Ardiles, L.; Flores, C.; Schneider, H.; Ruiz-Ortega, M.; Egido, J. NF-kappaB activation and overexpression of regulated genes in human diabetic nephropathy. Nephrol. Dial. Transplant 2004, 19, 2505–2512. [Google Scholar] [CrossRef]
- Mezzano, S.A.; Droguett, M.A.; Burgos, M.E.; Ardiles, L.G.; Aros, C.A.; Caorsi, I.; Egido, J. Overexpression of chemokines, fibrogenic cytokines, and myofibroblasts in human membranous nephropathy. Kidney Int. 2000, 57, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Schmid, H.; Boucherot, A.; Yasuda, Y.; Henger, A.; Brunner, B.; Eichinger, F.; Nitsche, A.; Kiss, E.; Bleich, M.; Gröne, H.J.; et al. Modular activation of nuclear factor-κB transcriptional programs in human diabetic nephropathy. Diabetes 2006, 55, 2993–3003. [Google Scholar] [CrossRef] [Green Version]
- Yi, B.; Hu, X.; Zhang, H.; Huang, J.; Liu, J.; Hu, J.; Li, W.; Huang, L. Nuclear NF-κB p65 in peripheral blood mononuclear cells correlates with urinary MCP-1, RANTES and the severity of type 2 diabetic nephropathy. PLoS ONE 2014, 9, e99633. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, inflammation, and metabolic disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, H.; Mi, R.Y.; Yoon, J.C.; Kitamura, M.; Hi, B.L. Role of high glucose-induced nuclear factor-κB activation in monocyte chemoattractant protein-1 expression by mesangial cells. J. Am. Soc. Nephrol. 2002, 13, 894–902. [Google Scholar] [PubMed]
- Najafian, B.; Fogo, A.B.; Lusco, M.A.; Alpers, C.E. AJKD Atlas of renal pathology: Diabetic nephropathy. Am. J. Kidney Dis. 2015, 66, e37–e38. [Google Scholar] [CrossRef] [PubMed]
- Hinz, M.; Scheidereit, C. The IκB kinase complex in NF-κB regulation and beyond. EMBO Rep. 2014, 15, 46–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, M.J.; D’Acquisto, F.; Madge, L.A.; Glockner, J.; Pober, J.S.; Ghosh, S. Selective inhibition of NF-κB activation by a peptide that blocks the interaction of NEMO with the IκB kinase complex. Science 2000, 289, 1550–1554. [Google Scholar] [CrossRef]
- Strickland, I.; Ghosh, S. Use of cell permeable NBD peptides for suppression of inflammation. Ann. Rheum. Dis. 2006, 65, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhang, L.; Mu, X.; Doebelin, C.; Nguyen, W.; Wallace, C.; Reay, D.P.; McGowan, S.J.; Corbo, L.; Clemens, P.R.; et al. Development of novel NEMO-binding domain mimetics for inhibiting IKK/NF-κB activation. PLoS Biol. 2018, 16, 1–27. [Google Scholar] [CrossRef]
- Oguiza, A.; Recio, C.; Lazaro, I.; Mallavia, B.; Blanco, J.; Egido, J.; Gomez-Guerrero, C. Peptide-based inhibition of IκB kinase/nuclear factor-κB pathway protects against diabetes-associated nephropathy and atherosclerosis in a mouse model of type 1 diabetes. Diabetologia 2015, 58, 1656–1667. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Li, D.; Bao, B.; Wan, X.; Pan, B.; Tu, J.; Wang, H.; Ouyang, Y.; Chen, Z.; Yin, X. NEMO-binding domain peptides alleviate perihematomal inflammation injury after experimental intracerebral hemorrhage. Neuroscience 2019, 409, 43–57. [Google Scholar] [CrossRef]
- Sun, Y.; Li, X.; Zhang, L.; Liu, X.; Jiang, B.; Long, Z.; Jiang, Y. Cell permeable NBD Peptide—Modified liposomes by hyaluronic acid coating for the synergistic targeted therapy of metastatic inflammatory breast cancer. Mol. Pharm. 2019, 16, 1140–1155. [Google Scholar] [CrossRef]
- Habineza Ndikuyeze, G.; Gaurnier-Hausser, A.; Patel, R.; Baldwin, A.S.; May, M.J.; Flood, P.; Krick, E.; Propert, K.J.; Mason, N.J. A phase I clinical trial of systemically delivered NEMO binding domain peptide in dogs with spontaneous activated B-cell like diffuse large B-cell lymphoma. PLoS ONE 2014, 9, e95404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudkins, K.L.; Pichaiwong, W.; Wietecha, T.; Kowalewska, J.; Banas, M.C.; Spencer, M.W.; Mu, A.; Koelling, M.; Pippin, J.W.; Shankland, S.J.; et al. BTBR Ob/Ob Mutant mice model progressive diabetic nephropathy. J. Am. Soc. Nephrol. 2010, 21, 1533–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichaiwong, W.; Hudkins, K.L.; Wietecha, T.; Nguyen, T.Q.; Tachaudomdach, C.; Li, W.; Askari, B.; Kobayashi, T.; O’Brien, K.D.; Pippin, J.W.; et al. Reversibility of structural and functional damage in a model of advanced diabetic nephropathy. J. Am. Soc. Nephrol. 2013, 24, 1088–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warram, J.M.; De Boer, E.; Korb, M.; Hartman, Y.; Kovar, J.; Markert, J.M.; Gillespie, G.Y.; Rosenthal, E.L. Fluorescence-guided resection of experimental malignant glioma using cetuximab-IRDye 800CW. Br. J. Neurosurg. 2015, 29, 850–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.E.; Tummers, W.S.; Teraphongphom, N.; van den Berg, N.S.; Hasan, A.; Ertsey, R.D.; Nagpal, S.; Recht, L.D.; Plowey, E.D.; Vogel, H.; et al. First in human intraoperative near infrared fluorescence imaging of glioblastoma using cetuximab-IRDye800. J. Neurooncol. 2018, 139, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavoz, C.; Sanchez, Y.; Orejudo, M.; Carpio, J.D.; Droguett, A.; Egido, J.; Mezzano, S.; Ruiz-Ortega, M. Interleukin-17A blockade reduces albuminuria and kidney injury in an accelerated model of diabetic nephropathy 1. Kidney Int. 2019, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Diez-Sampedro, A.; Lenz, O.; Fornoni, A. Podocytopathy in diabetes: A metabolic and endocrine disorder. Am. J. Kidney Dis. 2011, 58, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Wiggins, R.C. The spectrum of podocytopathies: A unifying view of glomerular diseases. Kidney Int. 2007, 71, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- Persson, F.; Rossing, P. Diagnosis of diabetic kidney disease: State of the art and future perspective. Kidney Int. Suppl. 2018, 8, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Porrini, E.; Ruggenenti, P.; Mogensen, C.E.; Barlovic, D.P.; Praga, M.; Cruzado, J.M.; Hojs, R.; Abbate, M.; de Vries, A.P.J. Non-proteinuric pathways in loss of renal function in patients with type 2 diabetes. Lancet Diabetes Endocrinol. 2015, 3, 382–391. [Google Scholar] [CrossRef]
- MacIsaac, R.J.; Jerums, G. Diabetic kidney disease with and without albuminuria. Curr. Opin. Nephrol. Hypertens. 2011, 20, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Yamanouchi, M.; Furuichi, K.; Hoshino, J.; Ubara, Y.; Wada, T. Nonproteinuric diabetic kidney disease. Clin. Exp. Nephrol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gohda, T.; Tomino, Y. Novel biomarkers for the progression of diabetic nephropathy: Soluble TNF receptors. Curr. Diab. Rep. 2013, 13, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Azushima, K.; Gurley, S.B.; Coffman, T.M. Modelling diabetic nephropathy in mice. Nat. Rev. Nephrol. 2017, 14, 48–56. [Google Scholar] [CrossRef]
- Singer, K.; Lumeng, C.N. The initiation of metabolic inflammation in childhood obesity. J. Clin. Invest. 2017, 127, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Reilly, S.M.; Saltiel, A.R. Adapting to obesity with adipose tissue inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef]
- Lamhamedi-Cherradi, S.-E.; Zheng, S.; Hilliard, B.A.; Xu, L.; Sun, J.; Alsheadat, S.; Liou, H.-C.; Chen, Y.H. Transcriptional regulation of type I diabetes by NF-κB. J. Immunol. 2003, 171, 4886–4892. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Li, L.; Yuan, W.; Zheng, L.; Guo, Z.; Huang, W. NEMO-Binding domain peptide attenuates lipopolysaccharide-induced acute lung injury by inhibiting the NF-κB signaling pathway. Mediators Inflamm. 2016, 2016, 734960. [Google Scholar] [CrossRef]
- Roy, A.; Mondal, S.; Kordower, J.H.; Pahan, K. Attenuation of microglial RANTES by NEMO-binding domain peptide inhibits the infiltration of CD8+ T cells in the nigra of hemiparkinsonian monkey. Neuroscience 2015, 302, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Rabadi, M.M.; Han, S.J.; Kim, M.; D’agati, V.; Lee, H.T. Peptidyl arginine deiminase-4 exacerbates ischemic AKI by finding NEMO. Am. J. Physiol. Ren. Physiol. 2019, 316, F1180–F1190. [Google Scholar] [CrossRef]
- Zoja, C.; Corna, D.; Camozzi, D.; Cattaneo, D.; Rottoli, D.; Batani, C.; Zanchi, C.; Abbate, M.; Remuzzi, G. How to fully protect the kidney in a severe model of progressive nephropathy: A multidrug approach. J. Am. Soc. Nephrol. 2002, 13, 2898–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallavia, B.; Recio, C.; Oguiza, A.; Ortiz-Muñoz, G.; Lazaro, I.; Lopez-Parra, V.; Lopez-Franco, O.; Schindler, S.; Depping, R.; Egido, J.; et al. Peptide inhibitor of NF-κB translocation ameliorates experimental atherosclerosis. Am. J. Pathol. 2013, 182, 1910–1921. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group (n) | Creatinine (mg/dL) | Total Cholesterol (mg/dL) | Triglycerides (mg/dL) | Uric Acid (mg/dL) |
|---|---|---|---|---|
| Vehicle (6) | 0.80 ± 0.08 | 158.0 ± 9.7 | 214.6 ± 69.1 | 3.90 ± 0.36 |
| NBD 6 μg (8) | 0.82 ± 0.08 | 177.4 ± 9.8 | 284.6 ± 35.4 | 3.30 ± 0.50 |
| NBD 10 μg (7) | 0.86 ± 0.04 | 207.9 ± 10.9 * | 289.7 ± 34.1 | 4.04 ± 0.29 |
| Mut 10 μg (7) | 0.74 ± 0.06 | 165.3 ± 10.7 | 239.1± 27.0 | 3.17 ± 0.22 |
| Gene | 5′-3′ Forward | 5′-3′ Reverse |
|---|---|---|
| Stat1 | TGAGATGTCCCGGATAGTGG | CGCCAGAGAGAAATTCGTGT |
| Stat3 | GTCTGCAGAGTTCAAGCACCT | TCCTCAGTCACGATCAAGGAG |
| Tnfα | ATGGCCTCCCTCTCATCAG | TTTGCTACGACGTGGGCTAC |
| Il-1β | GCTGAAAGCTCTCCACCTCA | CTTGGGATCCACACTCTCCAG |
| Ccl2 | AGCTCTCTCTTCCTCCACCA | GGCGTTAACTGCATCTGGCT |
| Ccl5 | AGAGGACTCTGAGACAGCACA | CGAGCCATATGGTGAGGCAG |
| Cxcl10 | ACTCCCCTTTACCCAGTGGA | CCACTTGAGCGAGGACTCAG |
| Nfe2l2 | GATCCGCCAGCTACTCCCAGGTTG | CAGGGCAAGCGACTCATGGTCATC |
| Nox4 | CCCTCCTGGCTGCATTAGTC | AACCCTCGAGGCAAAGATCC |
| Sod1 | GGAACCATCCACTTCGAGCA | CTGCACTGGTACAGCCTTGT |
| Catalase | GGTGCCCCCAACTATTACCC | GAATGTCCGCACCTGAGTGA |
| 18s | CCGTCGTAGTTCCGACCATAA | CAGCTTTGCAACCATACTCCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Opazo-Ríos, L.; Plaza, A.; Sánchez Matus, Y.; Bernal, S.; Lopez-Sanz, L.; Jimenez-Castilla, L.; Carpio, D.; Droguett, A.; Mezzano, S.; Egido, J.; et al. Targeting NF-κB by the Cell-Permeable NEMO-Binding Domain Peptide Improves Albuminuria and Renal Lesions in an Experimental Model of Type 2 Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 4225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124225
Opazo-Ríos L, Plaza A, Sánchez Matus Y, Bernal S, Lopez-Sanz L, Jimenez-Castilla L, Carpio D, Droguett A, Mezzano S, Egido J, et al. Targeting NF-κB by the Cell-Permeable NEMO-Binding Domain Peptide Improves Albuminuria and Renal Lesions in an Experimental Model of Type 2 Diabetic Nephropathy. International Journal of Molecular Sciences. 2020; 21(12):4225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124225
Chicago/Turabian StyleOpazo-Ríos, Lucas, Anita Plaza, Yenniffer Sánchez Matus, Susana Bernal, Laura Lopez-Sanz, Luna Jimenez-Castilla, Daniel Carpio, Alejandra Droguett, Sergio Mezzano, Jesús Egido, and et al. 2020. "Targeting NF-κB by the Cell-Permeable NEMO-Binding Domain Peptide Improves Albuminuria and Renal Lesions in an Experimental Model of Type 2 Diabetic Nephropathy" International Journal of Molecular Sciences 21, no. 12: 4225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124225