Signaling Modulations of miR-206-3p in Tooth Morphogenesis

 ,
,
Abstract
:1. Introduction
2. Results
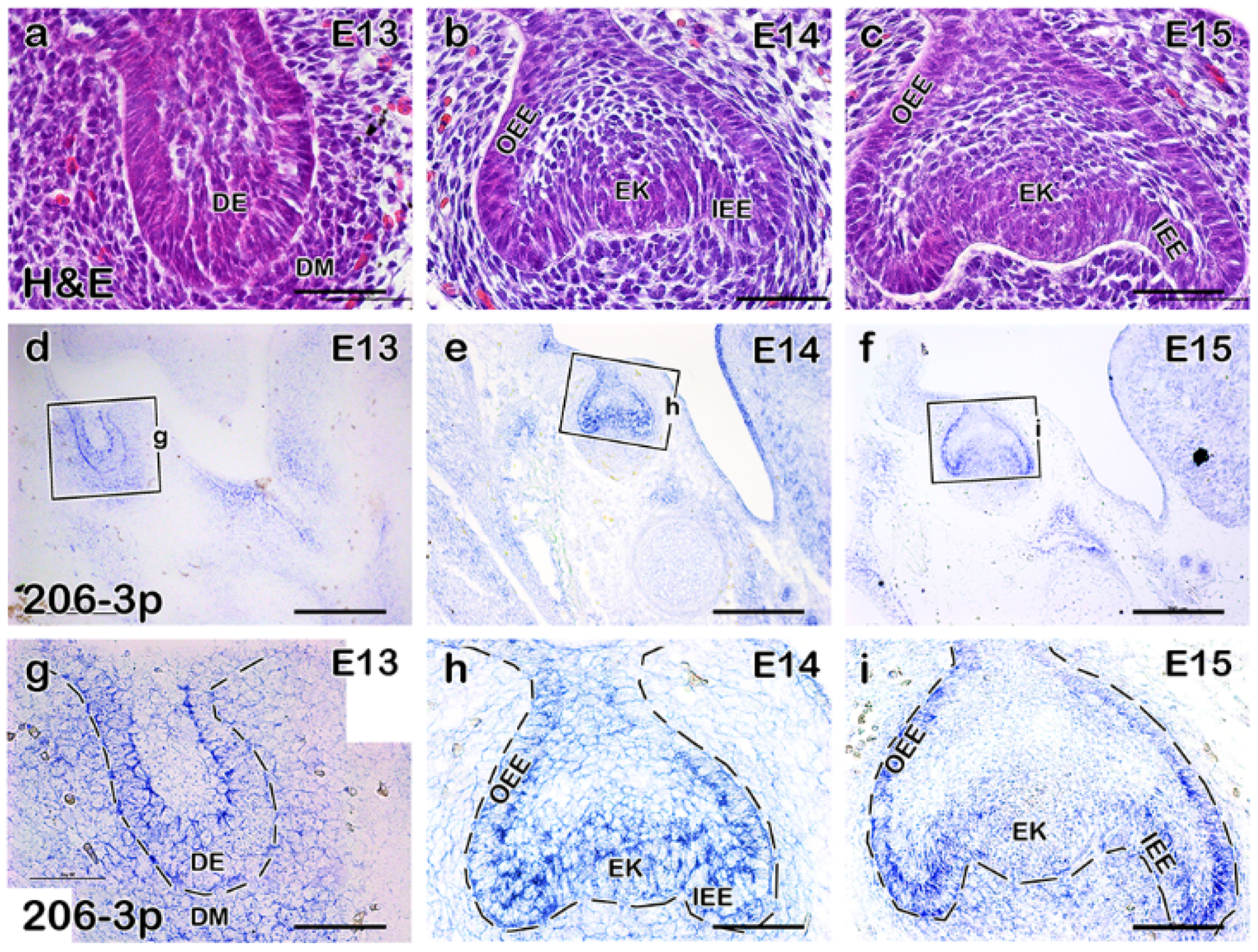
2.1. miR-206-3p Is Moderately Expressed in the Developing Molar
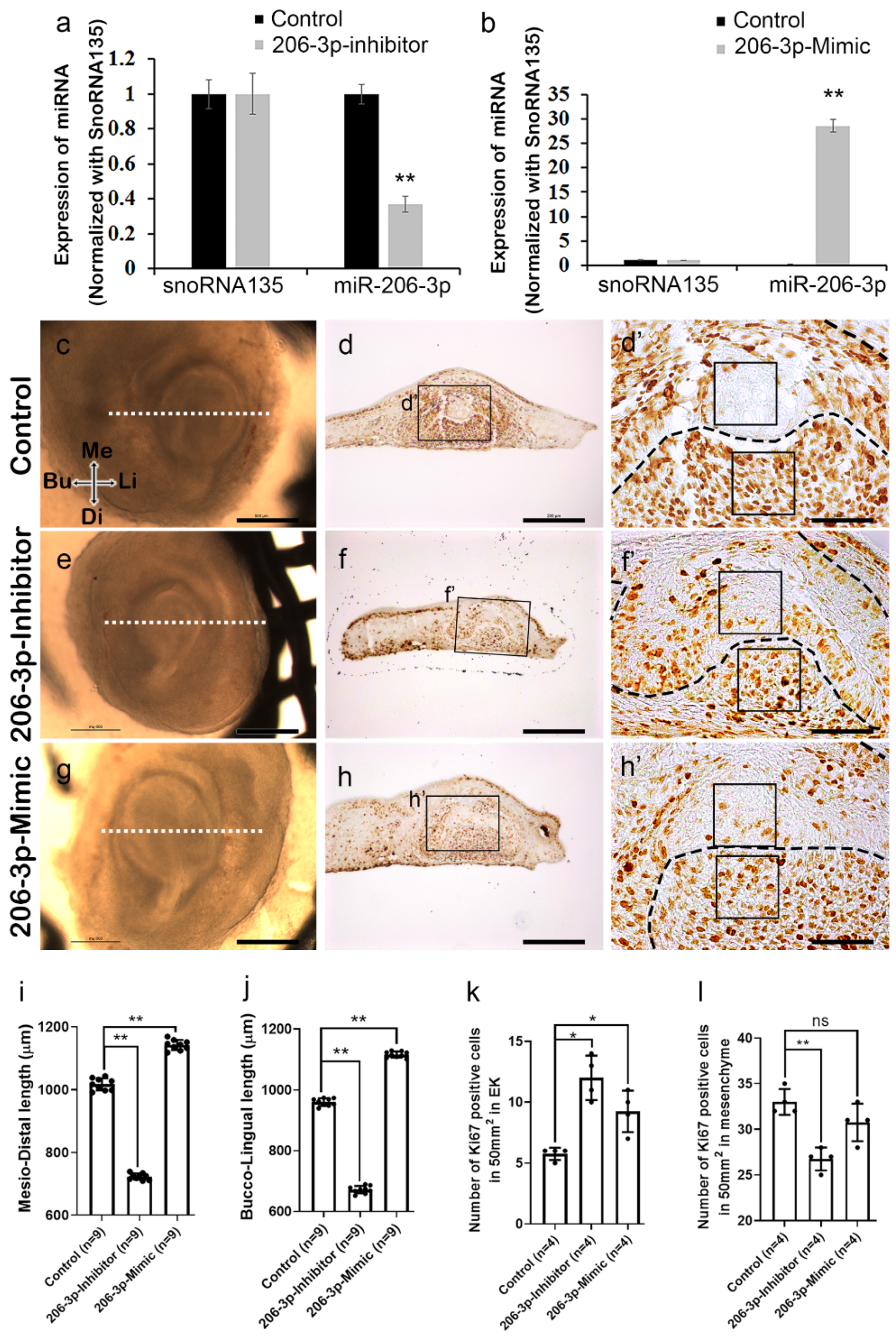
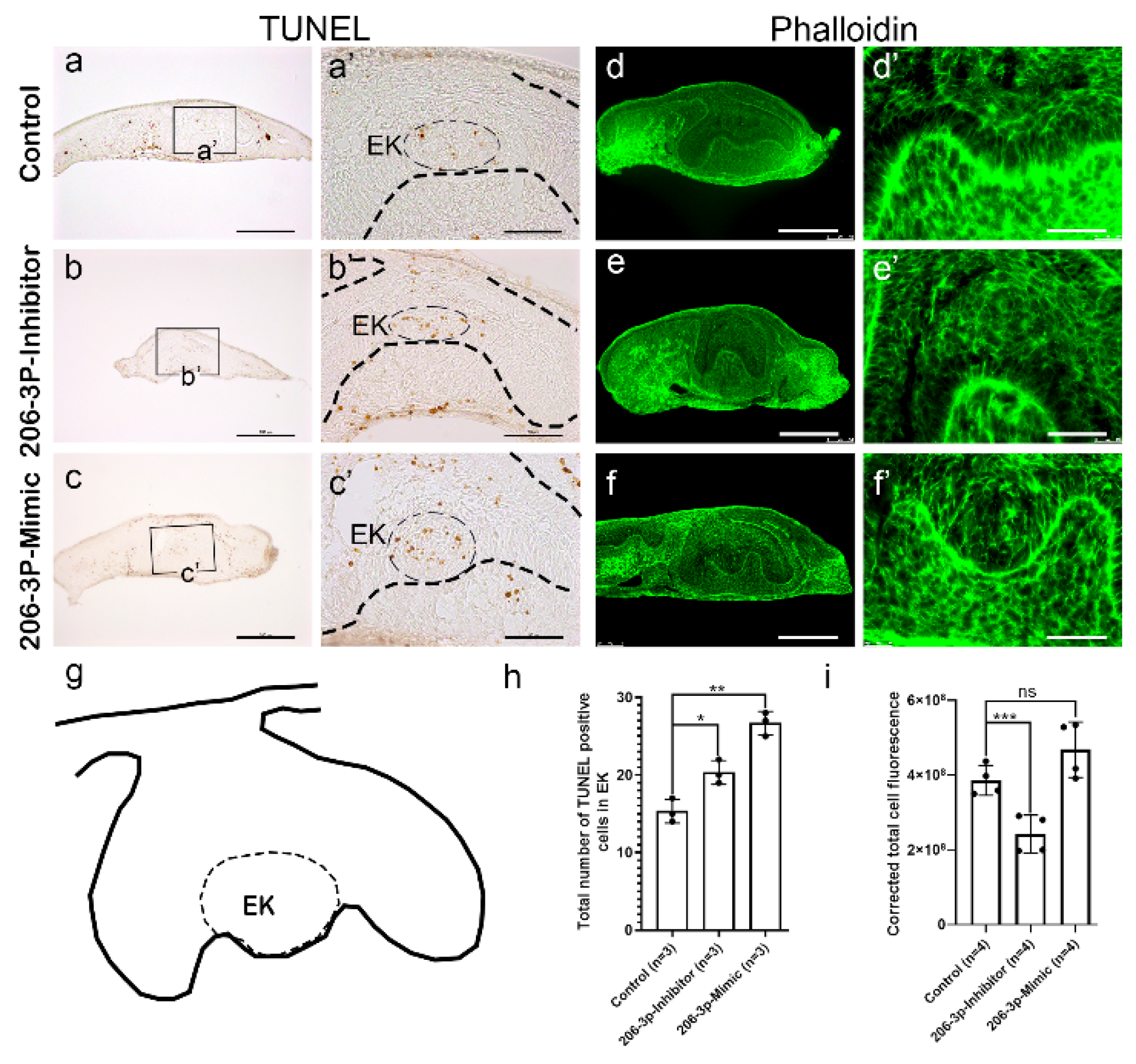
2.2. miR-206-3p Regulates Cell Dynamics Ex Vivo
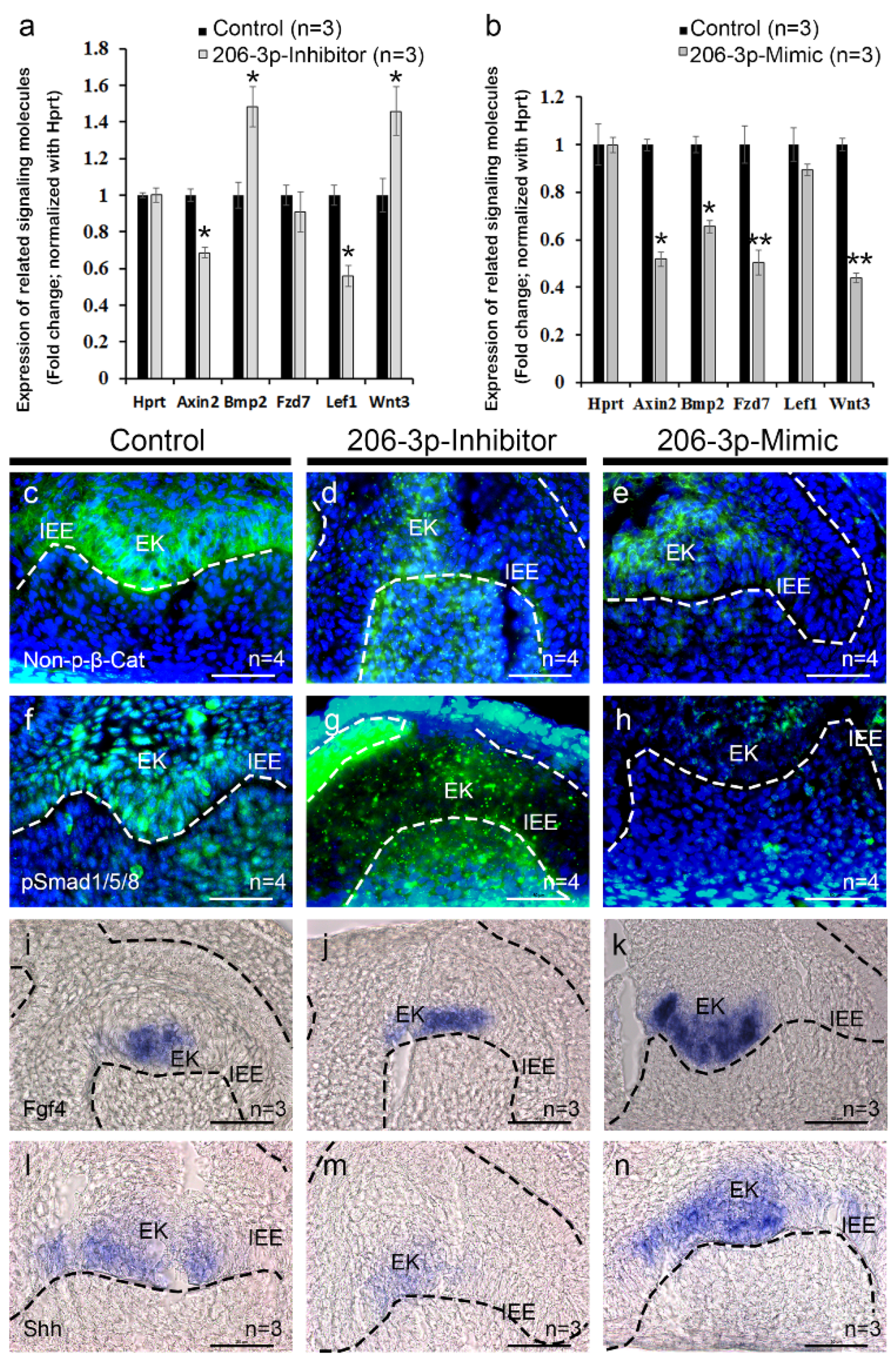
2.3. miR-206-3p Modulates Tooth Specific Signaling
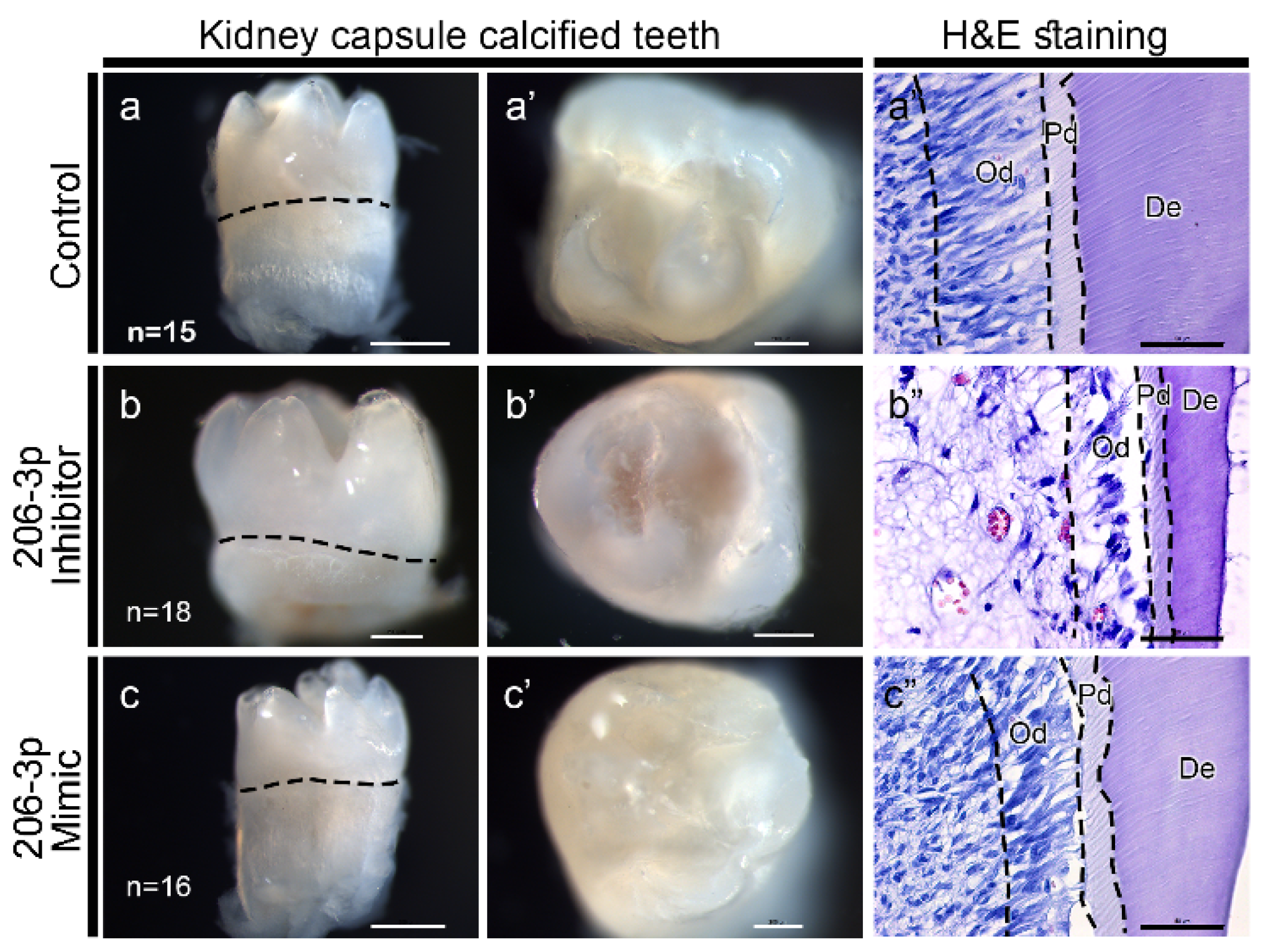
2.4. miR-206-3p Regulates Tooth Pulp Morphogenesis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. In Situ Hybridization
4.3. In Vitro Organ Cultivation and Renal Capsule Transplantations
4.4. Histology and Immunohistochemistry
4.5. TUNEL Assay
4.6. Phalloidin Staining
4.7. Quantitative PCR (qPCR)
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Balic, A.; Thesleff, I. Tissue Interactions Regulating Tooth Development and Renewal. Curr. Top. Dev. Biol. 2015, 115, 157–186. [Google Scholar] [CrossRef] [PubMed]
- Aurrekoetxea, M.; Irastorza, I.; García-Gallastegui, P.; Jiménez-Rojo, L.; Nakamura, T.; Yamada, Y.; Ibarretxe, G.; Unda, F.J. Wnt/β-Catenin Regulates the Activity of Epiprofin/Sp6, SHH, FGF, and BMP to Coordinate the Stages of Odontogenesis. Front. Cell Dev. Biol. 2016, 4, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Li, H.; Liu, Y.; Lin, X.; Lin, Y.; Wang, Y.; Hu, X.; Zhang, Y. Expression patterns of WNT/β -CATENIN signaling molecules during human tooth development. J. Mol. Histol. 2014, 45, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, L.; Sharpe, P.T. Expression of Wnt signalling pathway genes during tooth development. Mech. Dev. 1999, 85, 197–200. [Google Scholar] [CrossRef]
- Liu, F.; Chu, E.Y.; Watt, B.; Zhang, Y.; Gallant, N.M.; Andl, T.; Yang, S.H.; Lu, M.M.; Piccolo, S.; Schmidt-Ullrich, R.; et al. Wnt/β-catenin signaling directs multiple stages of tooth morphogenesis. Dev. Biol. 2008, 313, 210–224. [Google Scholar] [CrossRef] [Green Version]
- Järvinen, E.; Shimomura-Kuroki, J.; Balic, A.; Jussila, M.; Thesleff, I. Mesenchymal Wnt/β-catenin signaling limits tooth number. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Kratochwil, K.; Galceran, J.; Tontsch, S.; Roth, W.; Grosschedl, R. FGF4, a direct target of LEF1 and Wnt signaling, can rescue the arrest of tooth organogenesis in Lef1−/− mice. Genes Dev. 2002, 16, 3173–3185. [Google Scholar] [CrossRef] [Green Version]
- Yi, R.; O’Carroll, D.; Pasolli, H.A.; Zhang, Z.; Dietrich, F.S.; Tarakhovsky, A.; Fuchs, E. Morphogenesis in skin is governed by discrete sets of differentially expressed microRNAs. Nat. Genet. 2006, 38, 356–362. [Google Scholar] [CrossRef]
- Andl, T.; Murchison, E.P.; Liu, F.; Zhang, Y.; Yunta-Gonzalez, M.; Tobias, J.W.; Andl, C.D.; Seykora, J.T.; Hannon, G.J.; Millar, S.E. The miRNA-Processing Enzyme Dicer Is Essential for the Morphogenesis and Maintenance of Hair Follicles. Curr. Biol. 2006, 16, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Huang, Z.; Amendt, B.A.; Venugopalan, S.R.; Martin, J.F.; Elangovan, S.; Skobe, Z.; Florez, S.; Margolis, H.C.; Wang, J.; et al. MicroRNAs Play a Critical Role in Tooth Development. J. Dent. Res. 2010, 89, 779–784. [Google Scholar] [CrossRef]
- Oommen, S.; Otsuka-Tanaka, Y.; Imam, N.; Kawasaki, M.; Kawasaki, K.; Jalani-Ghazani, F.; Anderegg, A.; Awatramani, R.; Hindges, R.; Sharpe, P.T.; et al. Distinct roles of MicroRNAs in epithelium and mesenchyme during tooth development. Dev. Dyn. 2012, 241, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Sehic, A.; Tulek, A.; Khuu, C.; Nirvani, M.; Sand, L.P.; Utheim, T.P. Regulatory roles of microRNAs in human dental tissues. Gene 2017, 596, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Khuu, C.; Nirvani, M.; Utheim, T.P.; Sehic, A. MicroRNAs: Modulators of Tooth Development. MicroRNA 2016, 5, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Lee, M.J.; Li, L.; Yoon, K.S.; Kim, K.S.; Jung, H.S. Failure of tooth formation mediated by miR-135a overexpression via BMP signaling. J. Dent. Res. 2014, 93, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Jheon, A.H.; Li, C.Y.; Wen, T.; Michon, F.; Klein, O.D. Expression of micrornas in the stem cell niche of the adult mouse incisor. PLoS ONE 2011, 6, e24536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Lin, H.; Zhang, L.; Sun, Q.; Yuan, G.; Zhang, L.; Chen, S.; Chen, Z. miR-145 and miR-143 Regulate Odontoblast Differentiation through Targeting Klf4 and Osx Genes in a Feedback Loop. J. Biol. Chem. 2013, 288, 9261–9271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, X.; Ren, J.; Wu, X.; Li, G.; Fan, Z.; Zhang, C.; Li, A.; Wang, S. Mandible exosomal ssc-mir-133b regulates tooth development in miniature swine via endogenous apoptosis. Bone Res. 2018, 6. [Google Scholar] [CrossRef]
- Tang, R.; Ma, F.; Li, W.; Ouyang, S.; Liu, Z.; Wu, J. mir-206-3p inhibits 3T3-L1 cell adipogenesis via the c-Met/Pi3k/Akt pathway. Int. J. Mol. Sci. 2017, 18, 1510. [Google Scholar] [CrossRef] [Green Version]
- Mcconechy, M.K.; Ding, J.; Senz, J.; Yang, W.; Melnyk, N.; Tone, A.A.; Prentice, L.M.; Wiegand, K.; Mcalpine, J.N.; Sohrab, P.; et al. Ovarian and endometrial endometrioid carcinomas have distinct CTNNB1 and PTEN mutation profiles. Mod. Pathol. 2014, 27, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Jukka Jernvall, I.T. Reiterative Signaling and Patterning during Mammalian/Rtooth Morphogenesis; Elsevier: Amsterdam, The Netherlands, 2000; pp. 19–29. [Google Scholar]
- Cho, S.W.; Lee, H.A.; Cai, J.; Lee, M.J.; Kim, J.Y.; Ohshima, H.; Jung, H.S. The primary enamel knot determines the position of the first buccal cusp in developing mice molars. Differentiation 2007, 75, 441–451. [Google Scholar] [CrossRef]
- Cai, J.; Cho, S.W.; Kim, J.Y.; Lee, M.J.; Cha, Y.G.; Jung, H.S. Patterning the size and number of tooth and its cusps. Dev. Biol. 2007, 304, 499–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.; Zhang, H.; Zhou, X.; Han, X.; Ren, Y.; Gao, T.; Xiao, Y.; De Crombrugghe, B.; Somerman, M.J.; Feng, J.Q. Genetic evidence for the vital function of osterix in cementogenesis. J. Bone Miner. Res. 2012, 27, 1080–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obara, N.; Lesot, H. Asymmetrical growth, differential cell proliferation, and dynamic cell rearrangement underlie epithelial morphogenesis in mouse molar development. Cell Tissue Res. 2007, 330, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Kim, J.; Hwang, J.J.; Jeong, H.G.; Han, S.S.; Park, W.; Ryu, K.; Seomun, H.; Kim, J.Y.; Cho, E.S.; et al. Regulation of root patterns in mammalian teeth. Sci. Rep. 2017, 7, 12714. [Google Scholar] [CrossRef] [Green Version]
- Hoon, J.; Tan, M.; Koh, C.-G. The Regulation of Cellular Responses to Mechanical Cues by Rho GTPases. Cells 2016, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Kim, T.H.; Bae, C.H.; Lee, J.C.; Ko, S.O.; Yang, X.; Jiang, R.; Cho, E.S. β-catenin is Required in Odontoblasts for Tooth Root Formation. J. Dent. Res. 2013, 92, 215–221. [Google Scholar] [CrossRef]
- Andl, T.; Reddy, S.T.; Gaddapara, T.; Millar, S.E. WNT signals are required for the initiation of hair follicle development. Dev. Cell 2002, 2, 643–653. [Google Scholar] [CrossRef]
- Lan, Y.; Jia, S.; Jiang, R. Molecular patterning of the mammalian dentition. Semin. Cell Dev. Biol. 2014, 25–26, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Jeong, L.; Han, J.; Ito, Y.; Bringas, P.; Chai, Y. Developmental expression of Smad1-7 suggests critical function of TGF-β/BMP signaling in regulating epithelial-mesenchymal interaction during tooth morphogenesis. Int. J. Dev. Biol. 2003, 47, 31–39. [Google Scholar] [CrossRef]
- Liu, W.; Selever, J.; Lu, M.-F.; Martin, J.F. Genetic dissection of Pitx2 in craniofacial development uncovers new functions in branchial arch morphogenesis, late aspects of tooth morphogenesis and cell migration. Development 2003, 130, 6375–6385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Ning, G.; Meng, A.; Wang, Q. MicroRNA-206 Regulates Cell Movements during Zebrafish Gastrulation by Targeting prickle1a and Regulating c-Jun N-Terminal Kinase 2 Phosphorylation. Mol. Cell. Biol. 2012, 32, 2934–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsu, K.; Kishigami, R.; Fujiwara, N.; Ishizeki, K.; Harada, H. Functional role of rho-kinase in ameloblast differentiation. J. Cell. Physiol. 2011, 226, 2527–2534. [Google Scholar] [CrossRef] [PubMed]
- Mendonsa, A.M.; Na, T.Y.; Gumbiner, B.M. E-cadherin in contact inhibition and cancer. Oncogene 2018, 37, 4769–4780. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [Green Version]
- Thesleff, I. Signalling regulating tooth morphogenesis Signalling In Tooth Development. J. Cell Sci. 2003, 116, 1647–1648. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Kuang-Hsien Hu, J.; Du, W.; Klein, O.D. Lineage tracing of epithelial cells in developing teeth reveals two strategies for building signaling centers. J. Biol. Chem. 2017, 292, 15062–15069. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.-J.E.; Li, L.; Jung, H.-S. Hippo pathway/Yap regulates primary enamel knot and dental cusp patterning in tooth morphogenesis. Cell Tissue Res. 2015, 362, 447–451. [Google Scholar] [CrossRef]
- Chen, J.; Lan, Y.; Baek, J.A.; Gao, Y.; Jiang, R. Wnt/beta-catenin signaling plays an essential role in activation of odontogenic mesenchyme during early tooth development. Dev. Biol. 2009, 334, 174–185. [Google Scholar] [CrossRef] [Green Version]
- Gluhak-Heinrich, J.; Guo, D.; Yang, W.; Harris, M.A.; Lichtler, A.; Kream, B.; Zhang, J.; Feng, J.Q.; Smith, L.C.; Dechow, P.; et al. New roles and mechanism of action of BMP4 in postnatal tooth cytodifferentiation. Bone 2010, 46, 1533–1545. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Ito, Y.; Xu, X.; Han, J.; Bringas, P.; Maeda, T.; Slavkin, H.C.; Grosschedl, R.; Chai, Y. LEF1 is a critical epithelial survival factor during tooth morphogenesis. Dev. Biol. 2005, 278, 130–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Harris, M.A.; Cui, Y.; Mishina, Y.; Harris, S.E.; Gluhak-Heinrich, J. Bmp2 is required for odontoblast differentiation and pulp vasculogenesis. J. Dent. Res. 2012, 91, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Zheng, Y.; Li, R.; Li, X.; Zhou, M.; Niu, Y.; Zhang, Q. β-catenin enhances odontoblastic differentiation of dental pulp cells through activation of Runx2. PLoS ONE 2014, 9, e88890. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seppala, M.; Fraser, G.J.; Birjandi, A.A.; Xavier, G.M.; Cobourne, M.T.; Roelink, H.; Conway, S.J. Sonic Hedgehog Signaling and Development of the Dentition. J. Dev. Biol. 2017, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Ahn, Y.; Adasooriya, D.; Woo, E.J.; Kim, H.J.; Hu, K.S.; Krumlauf, R.; Cho, S.W. Shh Plays an Inhibitory Role in Cusp Patterning by Regulation of Sostdc1. J. Dent. Res. 2019, 98, 98–106. [Google Scholar] [CrossRef]
- Lu, C.P.; Polak, L.; Keyes, B.E.; Fuchs, E. Spatiotemporal antagonism in mesenchymal-epithelial signaling in sweat versus hair fate decision. Science 2016, 354, aah6102. [Google Scholar] [CrossRef] [Green Version]
- Michon, F.; Tummers, M.; Kyyrönen, M.; Frilander, M.J.; Thesleff, I. Tooth morphogenesis and ameloblast differentiation are regulated by micro-RNAs. Dev. Biol. 2010, 340, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Jheon, A.; Li, X.; Sun, Z.; Wang, J.; Florez, S.; Zhang, Z.; McManus, M.T.; Klein, O.D.; Amendt, B.A. The Pitx2:miR-200c/141:noggin pathway regulates Bmp signaling and ameloblast differentiation. Development 2013, 140, 3348–3359. [Google Scholar] [CrossRef] [Green Version]
- Neupane, S.; Sohn, W.J.; Rijal, G.; Lee, Y.J.; Lee, S.; Yamamoto, H.; An, C.H.; Cho, S.W.; Lee, Y.; Shin, H.I.; et al. Developmental regulations of Perp in mice molar morphogenesis. Cell Tissue Res. 2014, 358, 109–121. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, M.J.; Cho, K.W.; Lee, J.M.; Kim, Y.J.; Kim, J.Y.; Jung, H.I.; Cho, J.Y.; Cho, S.W.; Jung, H.S. Shh and ROCK1 modulate the dynamic epithelial morphogenesis in circumvallate papilla development. Dev. Biol. 2009, 325, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Lombaert, I.M.A.; Hauser, B.R.; Patel, V.N.; Hoffman, M.P. Exosomal MicroRNA Transport from Salivary Mesenchyme Regulates Epithelial Progenitor Expansion during Organogenesis. Dev. Cell 2017, 40, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, N.; Neupane, S.; Gwon, G.J.; Kim, J.Y.; An, C.H.; Lee, S.; Sohn, W.J.; Lee, Y.; Kim, J.Y. Grhl3 modulates epithelial structure formation of the circumvallate papilla during mouse development. Histochem. Cell Biol. 2017, 147, 5–16. [Google Scholar] [CrossRef]
- Busk, P.K. A tool for design of primers for microRNA-specific quantitative RT-qPCR. BMC Bioinform. 2014, 15, 29. [Google Scholar] [CrossRef] [Green Version]
- Neupane, S.; Sohn, W.J.; Gwon, G.J.; Kim, K.R.; Lee, S.; An, C.H.; Suh, J.Y.; Shin, H.I.; Yamamoto, H.; Cho, S.W.; et al. The role of APCDD1 in epithelial rearrangement in tooth morphogenesis. Histochem. Cell Biol. 2015, 144, 377–387. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession | Primer Sequence | References | Product Size (bp) | Remark | |
|---|---|---|---|---|---|---|
| Axin2 | BC057338.1 | Forward | TGAAGAAGAGGAGTGGACGT | [56] | 115 | Wnt signaling |
| Reverse | AGCTGTTTCCGTGGATCTCA | |||||
| Bmp2 | NM_007553.3 | Forward | AAGTGGCCCATTTAGAGGAG | [56] | 104 | Bmp signaling |
| Reverse | CAATGGCCTTATCTGTGACC | |||||
| Fzd7 | NM_008057.3 | Forward | AAGGGGGAAACTGCGGTATG | 203 | Wnt Signaling | |
| Reverse | TCAAAACCATCTCTCGCCCC | |||||
| Lef1 | NM_010703.4 | Forward | ACAGCGACGAGCACTTTTCT | [56] | 82 | EK signaling |
| Reverse | TGTCTGGACATGCCTTGCTT | |||||
| Wnt3 | NM_009521.2 | Forward | TCGAGGCTTACCTTTGCACAT | 114 | Wnt signaling | |
| Reverse | TGGTCTTGTCCTTCCCACCA | |||||
| Hprt | NM_013556.1 | Forward | CCTAAGATGATCGCAAGTTG | [56] | 86 | Internal standard |
| Reverse | CCACAGGGACTAGAACACCTGCTAA | |||||
| Primer Sequences for miRNA qPCR | ||||||
| Gene | Accession | Primer Sequence | ||||
| miR-206-3p | MI0000249 (miRBase) | Forward | GCAGTGGAATGTAAGGAAGT | |||
| Reverse | CCAGTTTTTTTTTTTTTTTCCACACA | |||||
| snoRNA135 | AF357323 (NCBI) | Forward | TGGAATTACCGGCAGATTGGTAGTGGTGAGCCTATGGT | |||
| Reverse | TCCAGTTTTTTTTTTTTTTTCTTCAGA | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neupane, S.; Aryal, Y.P.; Kim, T.-Y.; Yeon, C.-Y.; An, C.-H.; Kim, J.-Y.; Yamamoto, H.; Lee, Y.; Sohn, W.-J.; Kim, J.-Y. Signaling Modulations of miR-206-3p in Tooth Morphogenesis. Int. J. Mol. Sci. 2020, 21, 5251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155251
Neupane S, Aryal YP, Kim T-Y, Yeon C-Y, An C-H, Kim J-Y, Yamamoto H, Lee Y, Sohn W-J, Kim J-Y. Signaling Modulations of miR-206-3p in Tooth Morphogenesis. International Journal of Molecular Sciences. 2020; 21(15):5251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155251
Chicago/Turabian StyleNeupane, Sanjiv, Yam Prasad Aryal, Tae-Young Kim, Chang-Yeol Yeon, Chang-Hyeon An, Ji-Youn Kim, Hitoshi Yamamoto, Youngkyun Lee, Wern-Joo Sohn, and Jae-Young Kim. 2020. "Signaling Modulations of miR-206-3p in Tooth Morphogenesis" International Journal of Molecular Sciences 21, no. 15: 5251. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155251