IDH2 Deficiency Is Critical in Myogenesis and Fatty Acid Metabolism in Mice Skeletal Muscle

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
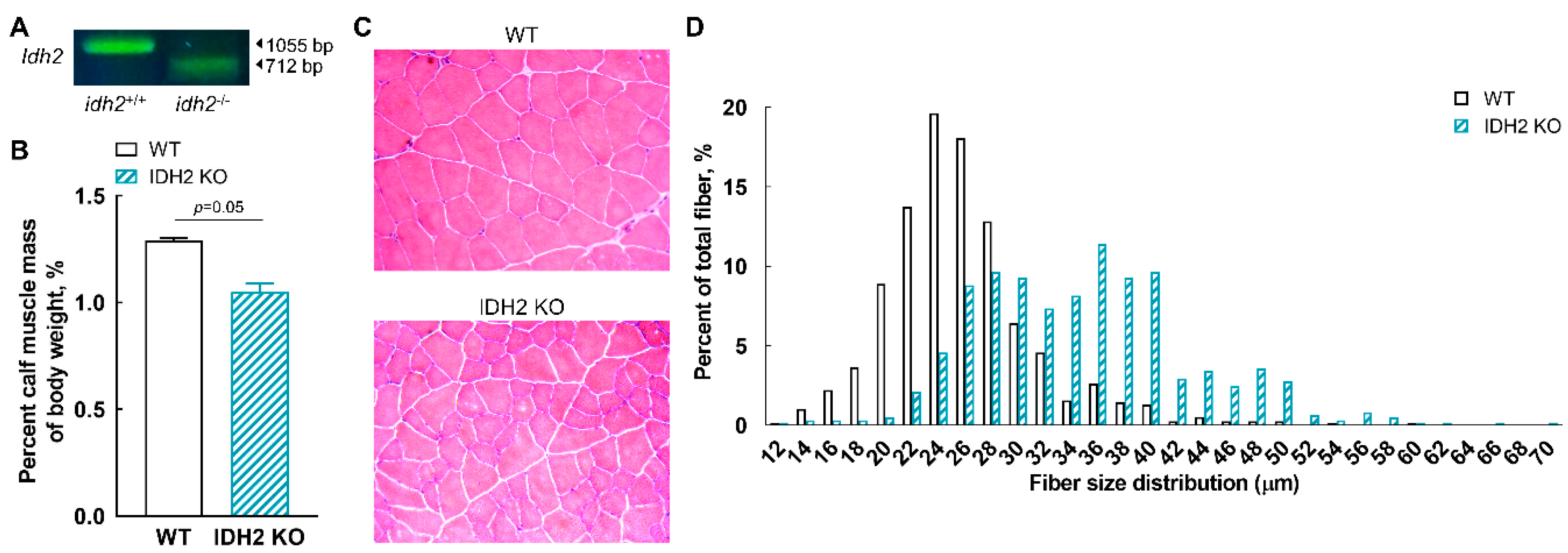
2.1. Genotype and Phenotype of IDH2 KO Mice
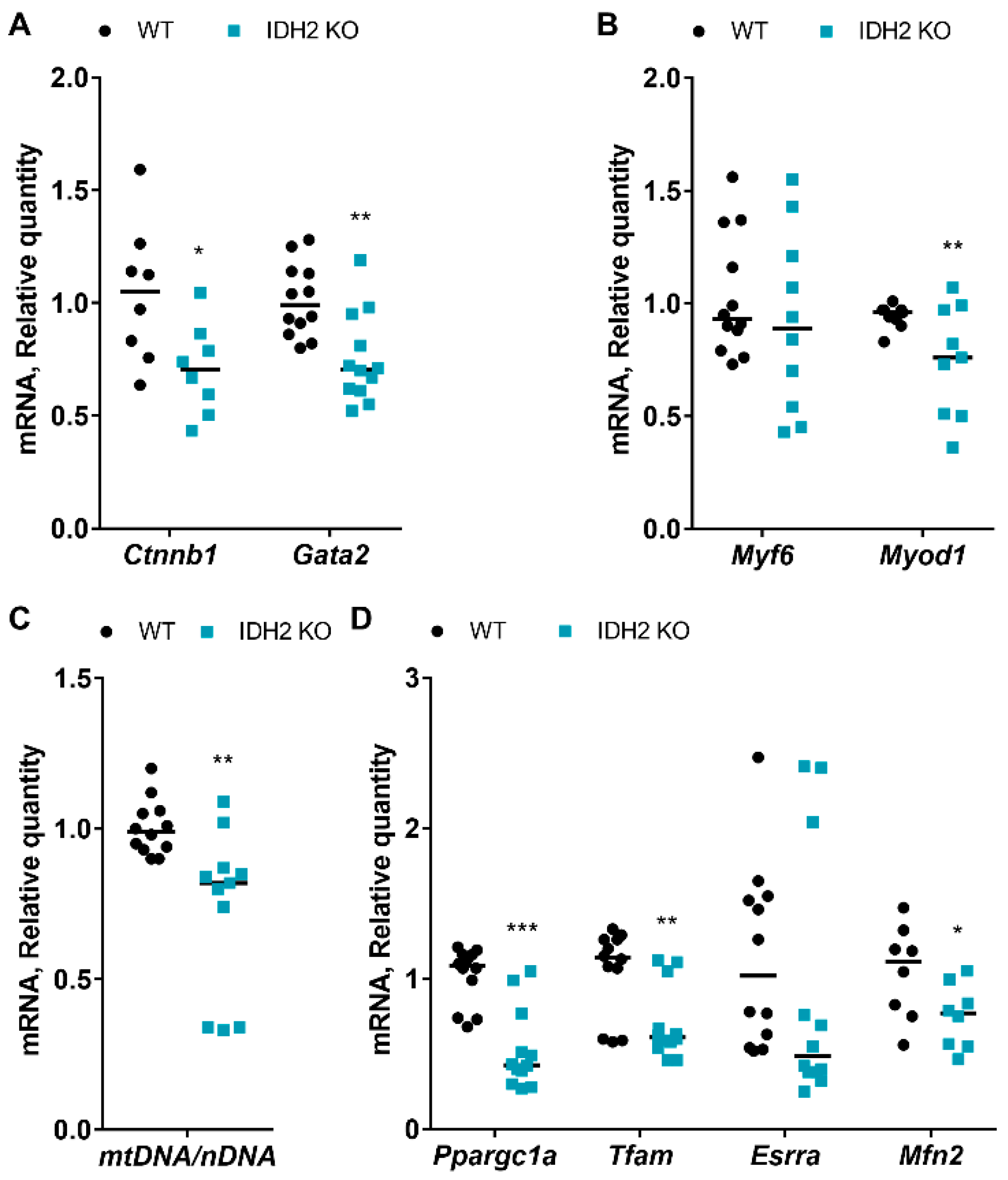
2.2. Effects of IDH2 on Myogenesis and Mitochondria Biogenesis
2.3. Effects of IDH2 on Adipogenesis, β-Oxidation, and Thermogenesis
3. Materials and Methods
3.1. Animal Care
3.2. Genotype Identification and Muscle Tissue Collection
3.3. RNA Isolation and cDNA Synthesis
3.4. Quantitative RT-PCR Analysis
3.5. Immunoblot Analysis
3.6. Measurement of Rectal Temperature in Response to Cold Stress
3.7. Histological Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 18S | 18S ribosomal RNA |
| Actb | beta-actin |
| Bmp7 | bone morphogenetic protein 7 |
| Cebpa | CCAAT/enhancer-binding protein alpha |
| Cebpb | CCAAT/enhancer-binding protein beta |
| Ctnnb1 | beta-catenin |
| Esrra | estrogen-related receptor alpha |
| Fabp4 | fatty acid binding protein 4 |
| Fat1 | protocadherin FAT 1 |
| Gata2 | GATA-binding factor 2 |
| IDH2 | mitochondrial NADP+-dependent isocitrate dehydrogenase |
| KO | knockout |
| Mfn2 | mitofusin 2 |
| mtDNA | mitochondria DNA |
| Myf6 | myogenic factor 6 |
| Myod1 | myogenic differentiation 1 |
| nDNA | nuclear DNA |
| Pparg | peroxisome proliferator-activated receptor gamma |
| Pppargc1a | proliferator-activated receptor gamma coactivator 1-alpha |
| Prdm16 | PR domain containing 16 |
| Tfam | mitochondrial transcription factor A |
| Ucp | uncoupling protein, mRNA |
| UCP | uncoupling protein, protein |
| WT | wild-type |
| Znf423 | zinc finger protein 423 |
References
- Lowell, B.B.; Shulman, G.I. Mitochondrial dysfunction and type 2 diabetes. Science 2005, 307, 384–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Yan, X.; Zhu, M.J.; McCormick, R.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Enhanced transforming growth factor-beta signaling and fibrogenesis in ovine fetal skeletal muscle of obese dams at late gestation. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1254–E1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, L.A.; Bal, N.C.; Periasamy, M. The role of skeletal-muscle-based thermogenic mechanisms in vertebrate endothermy. Biol. Rev. Camb. Philos. Soc. 2015, 90, 1279–1297. [Google Scholar] [CrossRef] [Green Version]
- Timmons, J.A.; Wennmalm, K.; Larsson, O.; Walden, T.B.; Lassmann, T.; Petrovic, N.; Hamilton, D.L.; Gimeno, R.E.; Wahlestedt, C.; Baar, K.; et al. Myogenic gene expression signature establishes that brown and white adipocytes originate from distinct cell lineages. Proc. Natl. Acad. Sci. USA 2007, 104, 4401–4406. [Google Scholar] [CrossRef] [Green Version]
- Grimpo, K.; Volker, M.N.; Heppe, E.N.; Braun, S.; Heverhagen, J.T.; Heldmaier, G. Brown adipose tissue dynamics in wild-type and ucp1-knockout mice: In vivo insights with magnetic resonance. J. Lipid Res. 2014, 55, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Berg, S.A.A.; Lichtenbelt, W.V.; van Dijk, K.W.; Schrauwen, P. Skeletal muscle mitochondrial uncoupling, adaptive thermogenesis and energy expenditure. Curr. Opin. Clin. Nutr. 2011, 14, 243–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, S.H.; Son, M.K.; Koh, H.J.; Lee, S.M.; Song, I.H.; Kim, Y.O.; Lee, Y.S.; Jeong, K.S.; Kim, W.B.; Park, J.W.; et al. Control of mitochondrial redox balance and cellular defense against oxidative damage by mitochondrial nadp+-dependent isocitrate dehydrogenase. J. Biol. Chem. 2001, 276, 16168–16176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Kim, S.H.; Park, K.M.; Lee, J.H.; Park, J.W. Increased obesity resistance and insulin sensitivity in mice lacking the isocitrate dehydrogenase 2 gene. Free Radic. Biol. Med. 2016, 99, 179–188. [Google Scholar] [CrossRef]
- Lee, S.J.; Cha, H.; Kim, H.; Lee, J.H.; Park, J.W. Amelioration of late-onset hepatic steatosis in idh2-deficient mice. Free Radic. Res. 2017, 51, 368–374. [Google Scholar] [CrossRef]
- Suzuki, A.; Pelikan, R.C.; Iwata, J. Wnt/β-catenin signaling regulates multiple steps of myogenesis by regulating step-specific targets. Mol. Cell. Biol. 2015, 35, 1763–1776. [Google Scholar] [CrossRef] [Green Version]
- Downie, D.; Delday, M.I.; Maltin, C.A.; Sneddon, A.A. Clenbuterol increases muscle fiber size and gata-2 protein in rat skeletal muscle in utero. Mol. Reprod. Dev. 2008, 75, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R.F.; Cicogna, A.C.; Rocha Campos, G.E.; da Silva Lopes, F.; Sugizaki, M.M.; Nogueira, C.R.; Dal Pai-Silva, M. Heart failure alters myod and mrf4 expressions in rat skeletal muscle. Int. J. Exp. Pathol. 2006, 87, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Phillips, N.R.; Sprouse, M.L.; Roby, R.K. Simultaneous quantification of mitochondrial DNA copy number and deletion ratio: A multiplex real-time pcr assay. Sci. Rep. 2014, 4, 3887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, M.C.; Arany, Z. The many roles of pgc-1α in muscle—Recent developments. Metab. Clin. Exp. 2014, 63, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Collu-Marchese, M.; Shuen, M.; Pauly, M.; Saleem, A.; Hood David, A. The regulation of mitochondrial transcription factor a (tfam) expression during skeletal muscle cell differentiation. Biosci. Rep. 2015, 35, e00221. [Google Scholar] [CrossRef]
- LaBarge, S.; McDonald, M.; Smith-Powell, L.; Auwerx, J.; Huss, J.M. Estrogen-related receptor-α (errα) deficiency in skeletal muscle impairs regeneration in response to injury. FASEB J. 2014, 28, 1082–1097. [Google Scholar] [CrossRef] [Green Version]
- Bach, D.; Naon, D.; Pich, S.; Soriano, F.X.; Vega, N.; Rieusset, J.; Laville, M.; Guillet, C.; Boirie, Y.; Wallberg-Henriksson, H.; et al. Expression of mfn2, the charcot-marie-tooth neuropathy type 2a gene, in human skeletal muscle: Effects of type 2 diabetes, obesity, weight loss, and the regulatory role of tumor necrosis factor α and interleukin-6. Diabetes 2005, 54, 2685–2693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, A.W.; Chen, L.; Fisher, S.J.; Szanto, I.; Ristow, M.; Jozsi, A.C.; Hirshman, M.F.; Rosen, E.D.; Goodyear, L.J.; Gonzalez, F.J.; et al. Muscle-specific pparγ-deficient mice develop increased adiposity and insulin resistance but respond to thiazolidinediones. J. Clin. Investig. 2003, 112, 608–618. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Das, A.K.; Yang, Q.-Y.; Zhu, M.-J.; Du, M. Zfp423 promotes adipogenic differentiation of bovine stromal vascular cells. PLoS ONE 2012, 7, e47496. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhang, W.; Chen, Y.; Li, Y.; Sun, L.; Liu, Y.; Liu, M.; Yu, M.; Li, X.; Han, J.; et al. Activation of peroxisome proliferator-activated receptor γ (pparγ) and cd36 protein expression: The dual pathophysiological roles of progesterone. J. Biol. Chem. 2016, 291, 15108–15118. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Arany, Z.; Seale, P.; Mepani, R.J.; Ye, L.; Conroe, H.M.; Roby, Y.A.; Kulaga, H.; Reed, R.R.; Spiegelman, B.M. Transcriptional control of preadipocyte determination by zfp423. Nature 2010, 464, 619–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, C.; Schneeberger, A.G.; Hoppeler, H.; Meyer, D.C. Correlation of atrophy and fatty infiltration on strength and integrity of rotator cuff repairs: A study in thirteen patients. J. Shoulder Elb. Surg. 2007, 16, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Adjeitey, C.N.-K.; Mailloux, R.J.; de Kemp, R.A.; Harper, M.-E. Mitochondrial uncoupling in skeletal muscle by ucp1 augments energy expenditure and glutathione content while mitigating ros production. Am. J. Physiol.-Endocrinol. Metab. 2013, 305, E405–E415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kim, S.Y.; Ku, H.J.; Jeon, Y.H.; Lee, H.W.; Lee, J.; Kwon, T.K.; Park, K.M.; Park, J.W. Suppression of tumorigenesis in mitochondrial nadp(+)-dependent isocitrate dehydrogenase knock-out mice. Biochim. Biophys. Acta 2014, 1842, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, J.H.; Tang, J.; Kim, Y.J.; Lee, J.H.; Shin, E.-C.; Zhao, J.; Kim, K.-H.; Hwang, K.A.; Huang, Y.; Kim, J.K. IDH2 Deficiency Is Critical in Myogenesis and Fatty Acid Metabolism in Mice Skeletal Muscle. Int. J. Mol. Sci. 2020, 21, 5596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165596
Pan JH, Tang J, Kim YJ, Lee JH, Shin E-C, Zhao J, Kim K-H, Hwang KA, Huang Y, Kim JK. IDH2 Deficiency Is Critical in Myogenesis and Fatty Acid Metabolism in Mice Skeletal Muscle. International Journal of Molecular Sciences. 2020; 21(16):5596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165596
Chicago/Turabian StylePan, Jeong Hoon, Jingsi Tang, Young Jun Kim, Jin Hyup Lee, Eui-Cheol Shin, Jiangchao Zhao, Kee-Hong Kim, Kyung A. Hwang, Yan Huang, and Jae Kyeom Kim. 2020. "IDH2 Deficiency Is Critical in Myogenesis and Fatty Acid Metabolism in Mice Skeletal Muscle" International Journal of Molecular Sciences 21, no. 16: 5596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165596