Medaka (Oryzias latipes) Embryo as a Model for the Screening of Compounds That Counteract the Damage Induced by Ultraviolet and High-Energy Visible Light

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
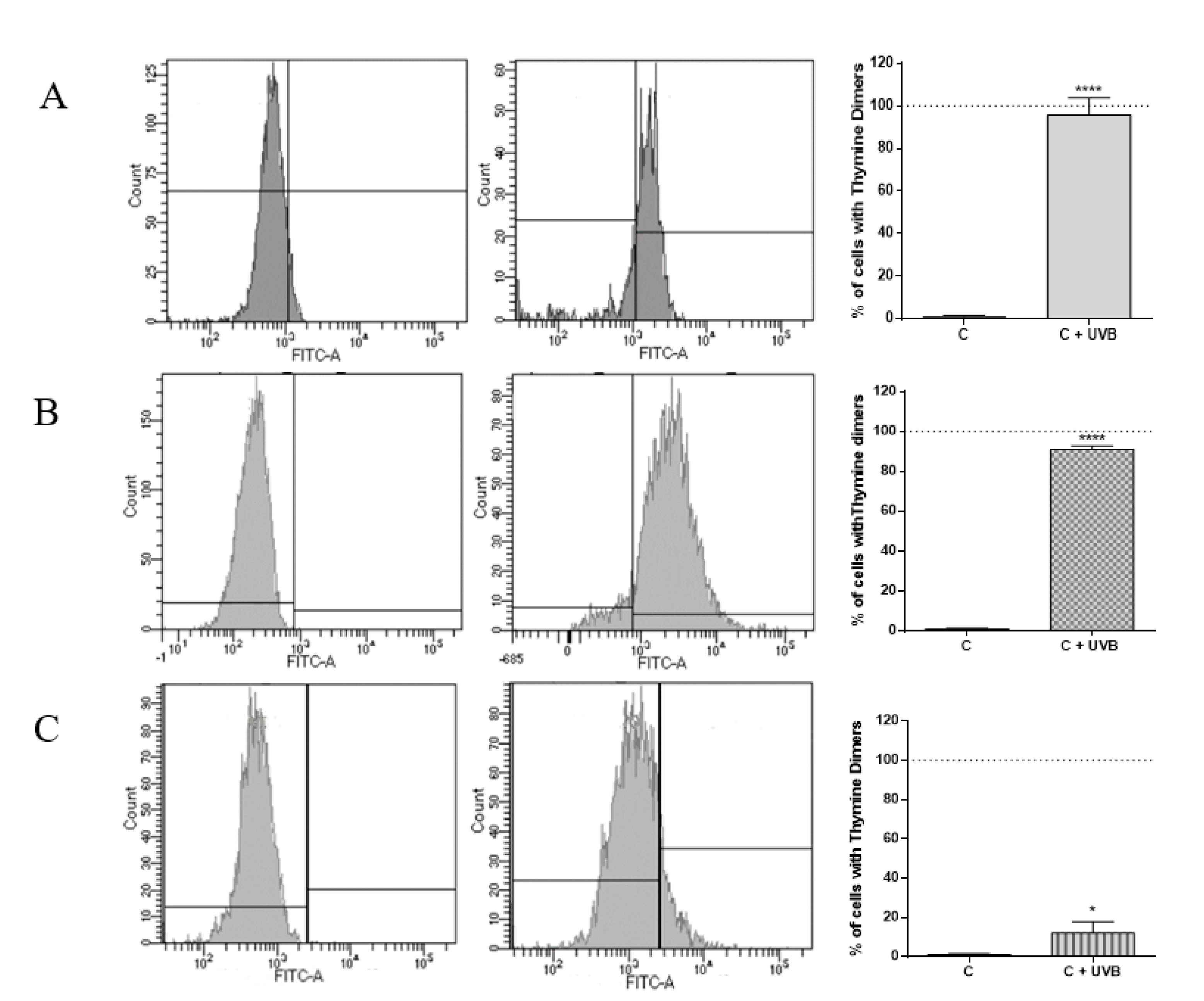
2.1. Evaluation of DNA Damage Generated by UVB in HaCat Culture, RHE and Medaka Embryos
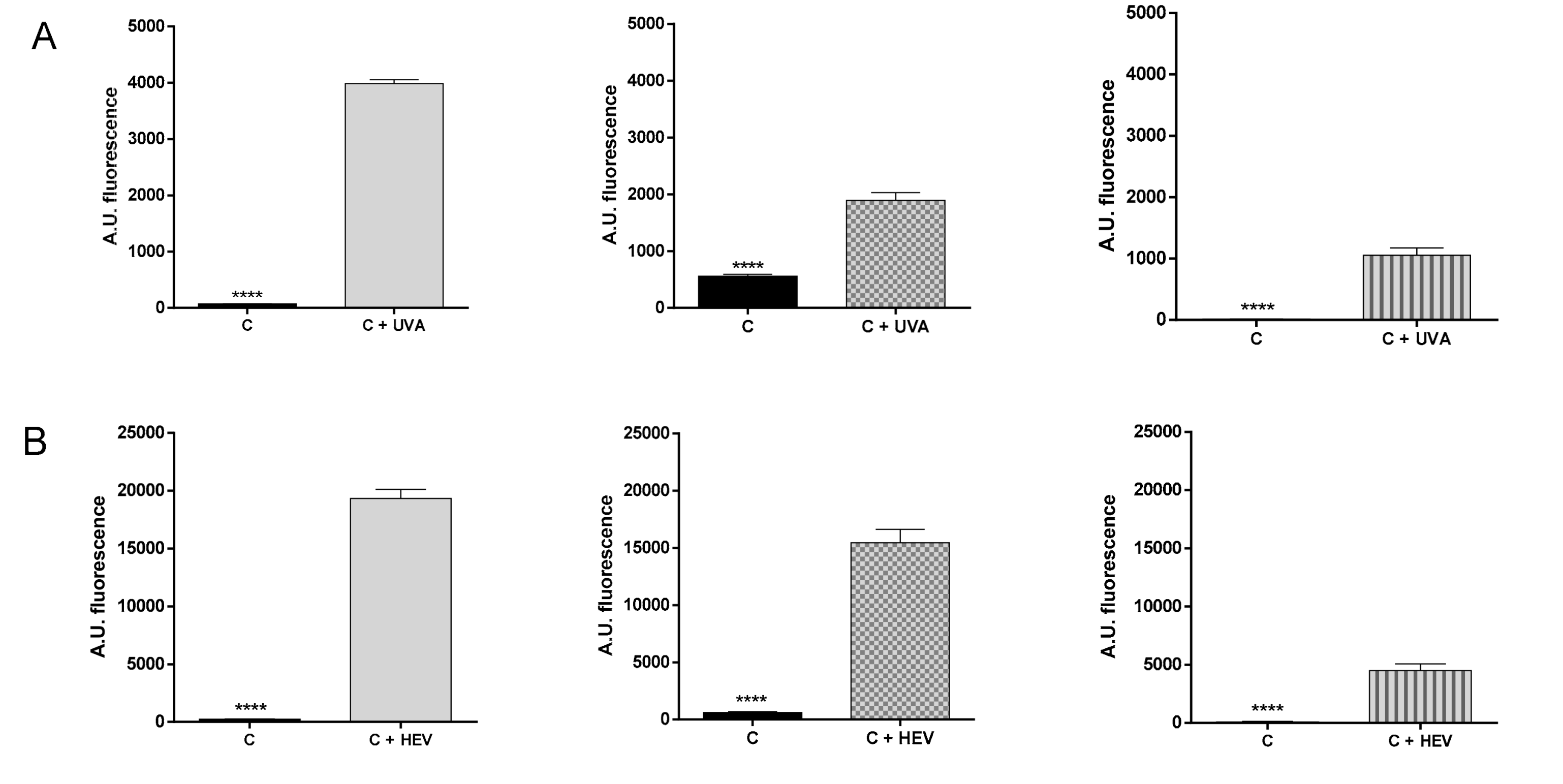
2.2. Evaluation of ROS Generation by UVA and Visible Light in HaCat Culture, RHE and Medaka Embryos
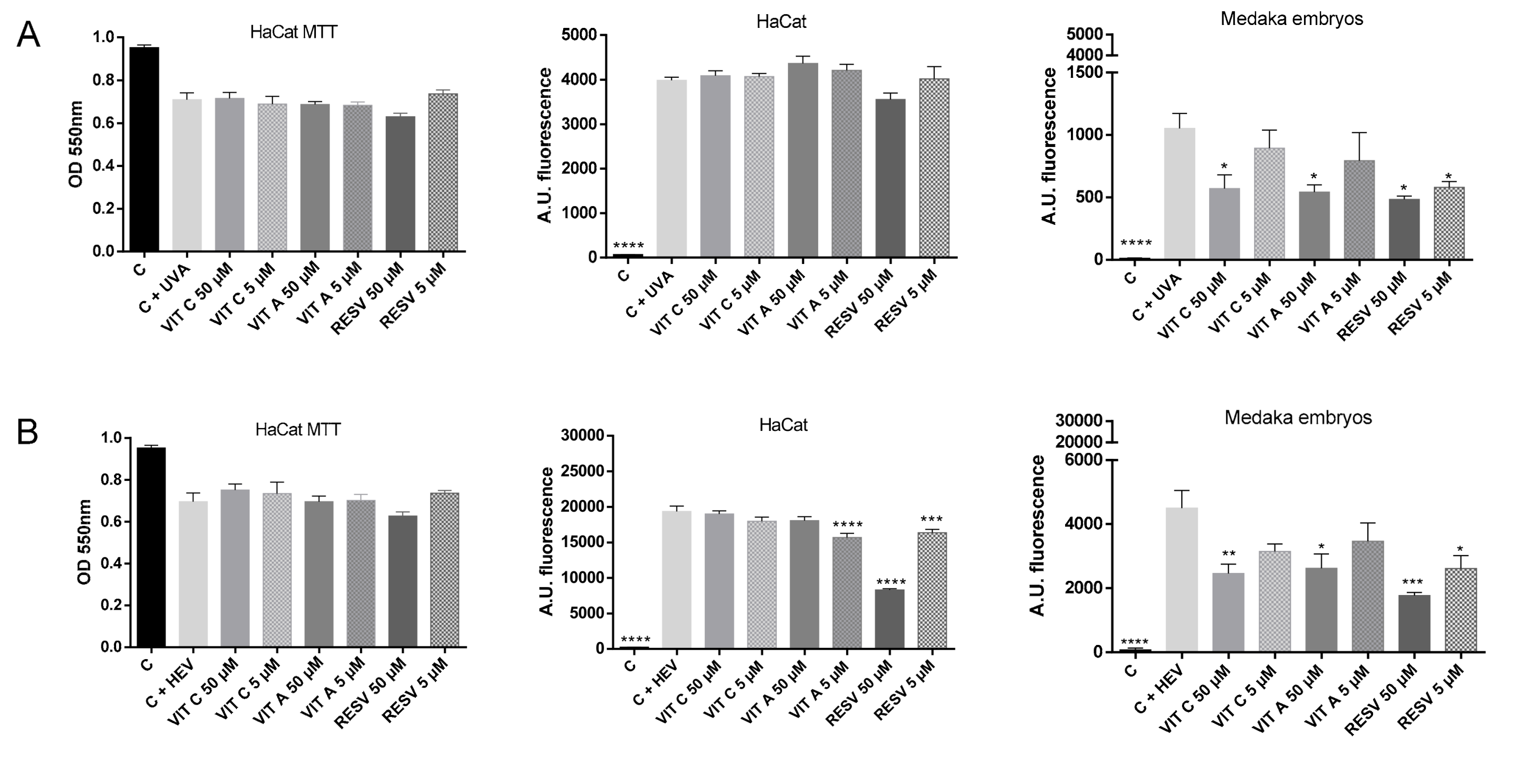
2.3. Testing of Antioxidant Compounds after UVA and HEV Radiation in HaCat Culture and Medaka Embryos
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Reconstructed Human Epidermis EpiDermTM (MatTek)
4.3. Fish Embryos
4.4. Flow Cytometric Analysis of Thymine Dimers (DNA Damage)
4.5. MTT Assay
4.6. UV Irradiation
4.7. ROS Detection
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AU | Arbitrary units |
| HEV | High Energy Visible light |
| RHE | Reconstructed human epidermis |
| ROS | Reactive Oxygen Species |
| UV | Ultraviolet |
References
- Juzeniene, A.; Moan, J. Beneficial effects of UV radiation other than via vitamin D production. Dermatoendocrinology 2012, 4, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaro-Ortiz, A.; Yan, B.; D’Orazio, J.A. Ultraviolet radiation, aging and the skin: Prevention of damage by topical cAMP manipulation. Molecules 2014, 19, 6202–6219. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.; Holman, D.M.; Maguire-Eisen, M. Ultraviolet Radiation Exposure and Its Impact on Skin Cancer Risk. Semin. Oncol. Nurs. 2016, 32, 241–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.K.; Trost, A.; Richter, K. Oxidative stress in aging human skin. Biomolecules 2015, 5, 545–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebel, F.; Kaur, S.; Ruvolo, E.; Kollias, N.; Southall, M.D. Irradiation of skin with visible light induces reactive oxygen species and matrix-degrading enzymes. J. Investig. Dermatol. 2012, 132, 1901–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Choi, M.S.; Bae, I.H.; Jung, J.Y.; Son, E.D.; Lee, T.R.; Shin, D.W. Short Wavelength Visible Light Suppresses Innate Immunity-Related Responses by Modulating Protein S-Nitrosylation in Keratinocytes. J. Invest. Dermatol. 2016, 136, 727–731. [Google Scholar] [CrossRef] [Green Version]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Intrinsic and extrinsic factors in skin aging: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef]
- Panich, U.; Sittithumcharee, G.; Rathviboon, N.; Jirawatnotai, S. Ultraviolet Radiation-Induced Skin Aging: The Role of DNA Damage and Oxidative Stress in Epidermal Stem Cell Damage Mediated Skin Aging. Stem Cells Int. 2016, 2016, 7370642. [Google Scholar] [CrossRef] [Green Version]
- Karran, P.; Brem, R. Protein oxidation, UVA and human DNA repair. DNA Repair (Amst.) 2016, 44, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Ravanat, J.L.; Douki, T.; Cadet, J. Direct and indirect effects of UV radiation on DNA and its components. J. Photochem. Photobiol. B. 2001, 63, 88–102. [Google Scholar] [CrossRef]
- Kamileri, I.; Karakasilioti, I.; Garinis, G.A. Nucleotide excision repair: New tricks with old bricks. Trends Genet. 2012, 28, 566–573. [Google Scholar] [CrossRef]
- Vogel, H.G. In Vitro Test Systems for Evaluation of the Physical Properties of the Skin. In Skin Models: Models to Study Function and Disease of Skin; Marks, R., Plewig, G., Eds.; Springer: Dusseldorf, Germany, 1986; pp. 412–419. [Google Scholar] [CrossRef]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D cell cultures—A comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Semlin, L.; Schäfer-Korting, M.; Borelli, C.; Korting, H.C. In vitro models for human skin disease. Drug Discov. Today 2011, 16, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Marionnet, C.; Pierrard, C.; Golebiewski, C.; Bernerd, F. Diversity of biological effects induced by longwave UVA rays (UVA1) in reconstructed skin. PLoS ONE 2014, 9, e105263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avon, S.L.; Wood, R.E. Porcine skin as an in-vivo model for ageing of human bite marks. J. Forensic Odontostomatol. 2005, 23, 30–39. [Google Scholar] [PubMed]
- Prasad, A.; Balukova, A.; Pospíšil, P. Triplet Excited Carbonyls and Singlet Oxygen Formation During Oxidative Radical Reaction in Skin. Front. Physiol. 2018, 9, 1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittbrodt, J.; Shima, A.; Schartl, M. Medaka--a model organism from the far East. Nat. Rev. Genet. 2002, 3, 53–64. [Google Scholar] [CrossRef]
- Adler, S.; Basketter, D.; Creton, S.; Pelkonen, O.; van Benthem, J.; Zuang, V.; Andersen, K.E.; Angers-Loustau, A.; Aptula, A.; Bal-Price, A.; et al. Alternative (non-animal) methods for cosmetics testing: Current status and future prospects-2010. Arch. Toxicol. 2011, 85, 367–485. [Google Scholar] [CrossRef]
- Lee, W.; Yang, K.L. Using medaka embryos as a model system to study biological effects of the electromagnetic fields on development and behavior. Ecotoxicol. Environ. Saf. 2014, 108, 187–194. [Google Scholar] [CrossRef]
- Aller, E.; Sánchez-Sánchez, A.V.; Chicote, J.U.; García-García, G.; Udaondo, P.; Cavallé, L.; Piquer-Gil, M.; García-España, A.; Díaz-Llopis, M.; Millán, J.M.; et al. Analysis of the Ush2a gene in medaka fish (Oryzias latipes). PLoS ONE 2013, 8, e74995. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; Chiang, C.Y.; Tsai, H.J. Zebrafish and Medaka: New model organisms for modern biomedical research. J. Biomed. Sci. 2016, 23, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Clares, M.P.; Serena, C.; Blasco, S.; Nebot, A.; del Castillo, L.; Soriano, C.; Domènech, A.; Sánchez-Sánchez, A.V.; Soler-Calero, L.; Mullor, J.L.; et al. Mn(II) complexes of scorpiand-like ligands. A model for the MnSOD active centre with high in vitro and in vivo activity. J. Inorg. Biochem. 2015, 143, 1–8. [Google Scholar] [CrossRef]
- Sánchez-Sánchez, A.V.; Leal-Tassias, A.; Rodríguez-Sánchez, N.; Piquer-Gil, M.; Martorell, P.; Genovés, S.; Acosta, C.; Burks, D.; Ramón, D.; Mullor, J.L. Use of Medaka Fish as Vertebrate Model to Study the Effect of Cocoa Polyphenols in the Resistance to Oxidative Stress and Life Span Extension. Rejuvenation Res. 2018, 21, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Shima, A.; Mitani, H. Medaka as a research organism: Past, present and future. Mech. Dev. 2004, 121, 599–604. [Google Scholar] [CrossRef]
- Le Guellec, D.; Morvan-Dubois, G.; Sire, J.Y. Skin development in bony fish with particular emphasis on collagen deposition in the dermis of the zebrafish (Danio rerio). Int. J. Dev. Biol. 2004, 48, 217–231. [Google Scholar] [CrossRef] [Green Version]
- Abdo, J.M.; Sopko, N.A.; Milner, S.M. The applied anatomy of human skin: A model for regeneration. Wound Med. 2020, 28, 100179. [Google Scholar] [CrossRef]
- Iwamatsu, T. Stages of normal development in the medaka Oryzias latipes. Mech. Dev. 2004, 121, 605–618. [Google Scholar] [CrossRef]
- Pei, D.S.; Strauss, P.R. Zebrafish as a model system to study DNA damage and repair. Mutat. Res. 2013, 743, 151–159. [Google Scholar] [CrossRef]
- Jackson, G.R.; Morgan, B.C.; Werrbach-Perez, K.; Perez-Polo, J.R. Antioxidant effect of retinoic acid on PC12 rat pheochromocytoma. Int J. Dev. Neurosci. 1991, 9, 161–163. [Google Scholar] [CrossRef]
- Gęgotek, A.; Ambrożewicz, E.; Jastrząb, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Rutin and ascorbic acid cooperation in antioxidant and antiapoptotic effect on human skin keratinocytes and fibroblasts exposed to UVA and UVB radiation. Arch. Dermatol. Res. 2019, 311, 203–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.; Lee, J.H. Protective effects of resveratrol on UVB-irradiated HaCaT cells through attenuation of the caspase pathway. Oncol. Rep. 2008, 19, 413–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bykov, V.J.; Jansen, C.T.; Hemminki, K. High levels of dipyrimidine dimers are induced in human skin by solar-simulating UV radiation. Cancer Epidemiol. Biomarkers Prev. 1998, 7, 199–202. [Google Scholar] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Vandersee, S.; Beyer, M.; Lademann, J.; Darvin, M.E. Blue-violet light irradiation dose dependently decreases carotenoids in human skin, which indicates the generation of free radicals. Oxid. Med. Cell Longev. 2015, 2015, 579675. [Google Scholar] [CrossRef] [Green Version]
- Marionnet, C.; Tricaud, C.; Bernerd, F. Exposure to non-extreme solar UV daylight: Spectral characterization, effects on skin and photoprotection. Int. J. Mol. Sci. 2014, 16, 68–90. [Google Scholar] [CrossRef]
- Varma, S.R.; Sivaprakasam, T.O.; Mishra, A.; Prabhu, S.; Rafiq, M.; Rangesh, P. Imiquimod-induced psoriasis-like inflammation in differentiated Human keratinocytes: Its evaluation using curcumin. Eur. J. Pharmacol. 2017, 813, 33–41. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediators Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Hofmann, U.; Priem, M.; Bartzsch, C.; Winckler, T.; Feller, K.H. A sensitive sensor cell line for the detection of oxidative stress responses in cultured human keratinocytes. Sensors (Basel) 2014, 14, 11293–11307. [Google Scholar] [CrossRef] [Green Version]
- Siddikuzzaman; Grace, V.M. Antioxidant potential of all-trans retinoic acid (ATRA) and enhanced activity of liposome encapsulated ATRA against inflammation and tumor-directed angiogenesis. Immunopharmacol. Immunotoxicol. 2013, 35, 164–173. [Google Scholar] [CrossRef]
- Farris, P.; Zeichner, J.; Berson, D. Efficacy and Tolerability of a Skin Brightening/Anti-Aging Cosmeceutical Containing Retinol 0.5%, Niacinamide, Hexylresorcinol, and Resveratrol. J. Drugs Dermatol. 2016, 15, 863–868. [Google Scholar] [PubMed]
- Saitoh, Y.; Umezaki, T.; Yonekura, N.; Nakawa, A. Resveratrol potentiates intracellular ascorbic acid enrichment through dehydroascorbic acid transport and/or its intracellular reduction in HaCaT cells. Mol. Cell Biochem. 2020, 467, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Valacchi, G.; Sticozzi, C.; Belmonte, G.; Cervellati, F.; Demaude, J.; Chen, N.; Krol, Y.; Oresajo, C. Vitamin C Compound Mixtures Prevent Ozone-Induced Oxidative Damage in Human Keratinocytes as Initial Assessment of Pollution Protection. PLoS ONE 2015, 10, e0131097. [Google Scholar] [CrossRef] [PubMed]
- Savini, I.; D’Angelo, I.; Ranalli, M.; Melino, G.; Avigliano, L. Ascorbic acid maintenance in HaCaT cells prevents radical formation and apoptosis by UV-B. Free Radic. Biol. Med. 1999, 26, 1172–1180. [Google Scholar] [CrossRef]
- Pérez-Sánchez, A.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Nutraceuticals for Skin Care: A Comprehensive Review of Human Clinical Studies. Nutrients 2018, 10, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, M.A.; Quan, T.; Voorhees, J.J.; Fisher, G.J. Extracellular matrix regulation of fibroblast function: Redefining our perspective on skin aging. J. Cell Commun. Signal. 2018, 12, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horiguchi, M.; Fujimori, C.; Ogiwara, K.; Moriyama, A.; Takahashi, T. Collagen type-I alpha1 chain mRNA is expressed in the follicle cells of the medaka ovary. Zoolog Sci. 2008, 25, 937–945. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Kelsh, R.N.; Inoue, C.; Momoi, A.; Kondoh, H.; Furutani-Seiki, M.; Ozato, K.; Wakamatsu, Y. The Tomita collection of medaka pigmentation mutants as a resource for understanding neural crest cell development. Mech Dev. 2004, 121, 841–859. [Google Scholar] [CrossRef]
- Maeda, T.; Chua, P.P.; Chong, M.T.; Sim, A.B.; Nikaido, O.; Tron, V.A. Nucleotide excision repair genes are upregulated by low-dose artificial ultraviolet B: Evidence of a photoprotective SOS response? J. Invest. Dermatol. 2001, 117, 1490–1497. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Richardson, J.; Verduzco, D.; Mitchell, D.L.; Patton, E.E. Zebrafish have a competent p53-dependent nucleotide excision repair pathway to resolve ultraviolet B-induced DNA damage in the skin. Version 2. Zebrafish 2009, 6, 405–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basketter, D.A.; White, I.R.; Kullavanijaya, P.; Tresukosol, P.; Wichaidit, M.; McFadden, J.P. Influence of vitamin C on the elicitation of allergic contact dermatitis to p-phenylenediamine. Contact Dermatitis 2016, 74, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Azevedo Martins, T.E.; Sales de Oliveira Pinto, C.A.; Costa de Oliveira, A.; Robles Velasco, M.V.; Gorriti Gutiérrez, A.R.; Cosquillo Rafael, M.F.; Tarazona, J.P.H.; Retuerto-Figueroa, M.G. Contribution of topical antioxidants to maintain healthy skin—A review. Sci. Pharm. 2020, 88, 27. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merino, M.; Mullor, J.L.; Sánchez-Sánchez, A.V. Medaka (Oryzias latipes) Embryo as a Model for the Screening of Compounds That Counteract the Damage Induced by Ultraviolet and High-Energy Visible Light. Int. J. Mol. Sci. 2020, 21, 5769. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165769
Merino M, Mullor JL, Sánchez-Sánchez AV. Medaka (Oryzias latipes) Embryo as a Model for the Screening of Compounds That Counteract the Damage Induced by Ultraviolet and High-Energy Visible Light. International Journal of Molecular Sciences. 2020; 21(16):5769. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165769
Chicago/Turabian StyleMerino, Marián, José Luis Mullor, and Ana Virginia Sánchez-Sánchez. 2020. "Medaka (Oryzias latipes) Embryo as a Model for the Screening of Compounds That Counteract the Damage Induced by Ultraviolet and High-Energy Visible Light" International Journal of Molecular Sciences 21, no. 16: 5769. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165769