Expression and Localization of BDNF/TrkB System in the Zebrafish Inner Ear

 , ,
, ,  , and
, and 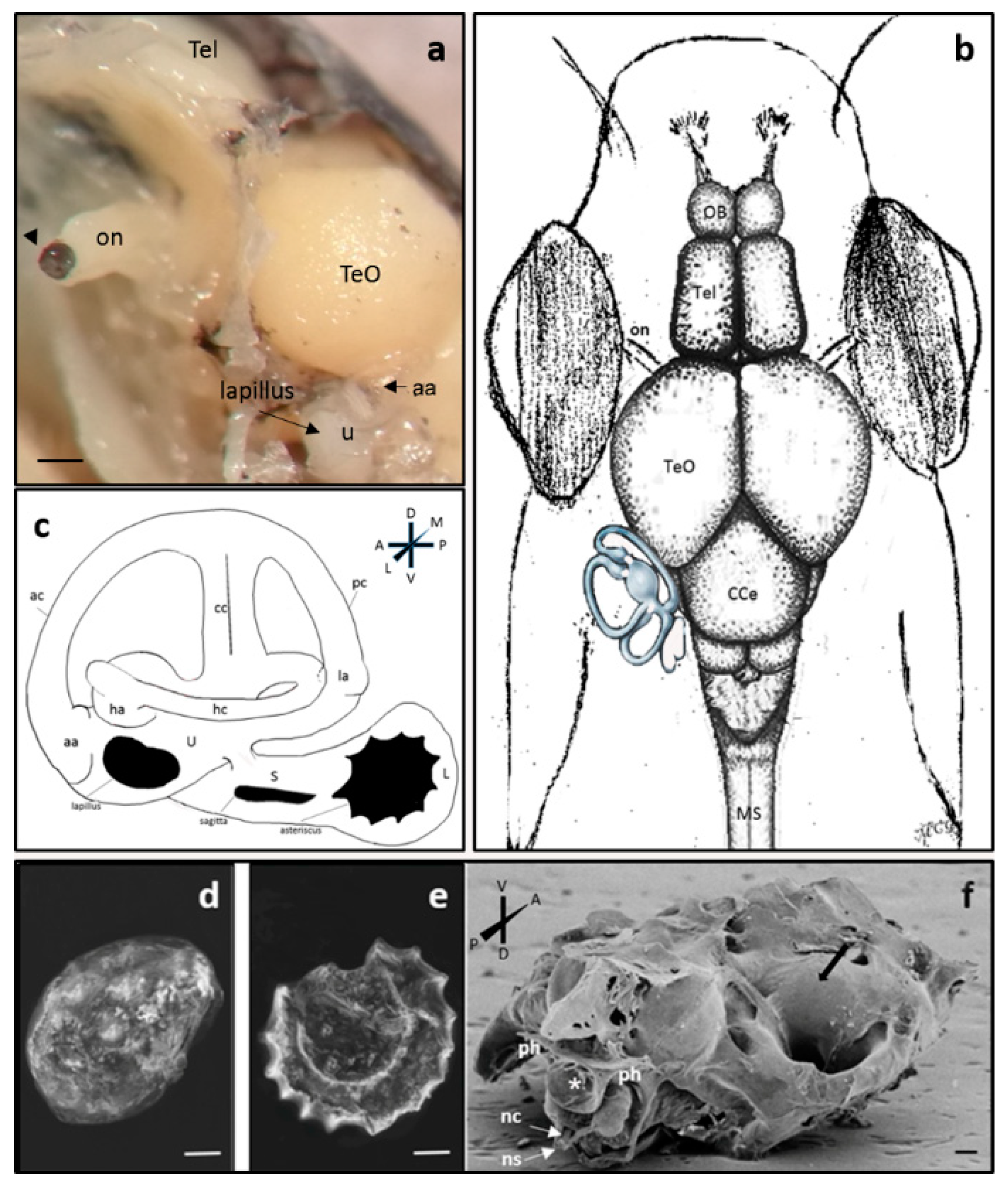{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
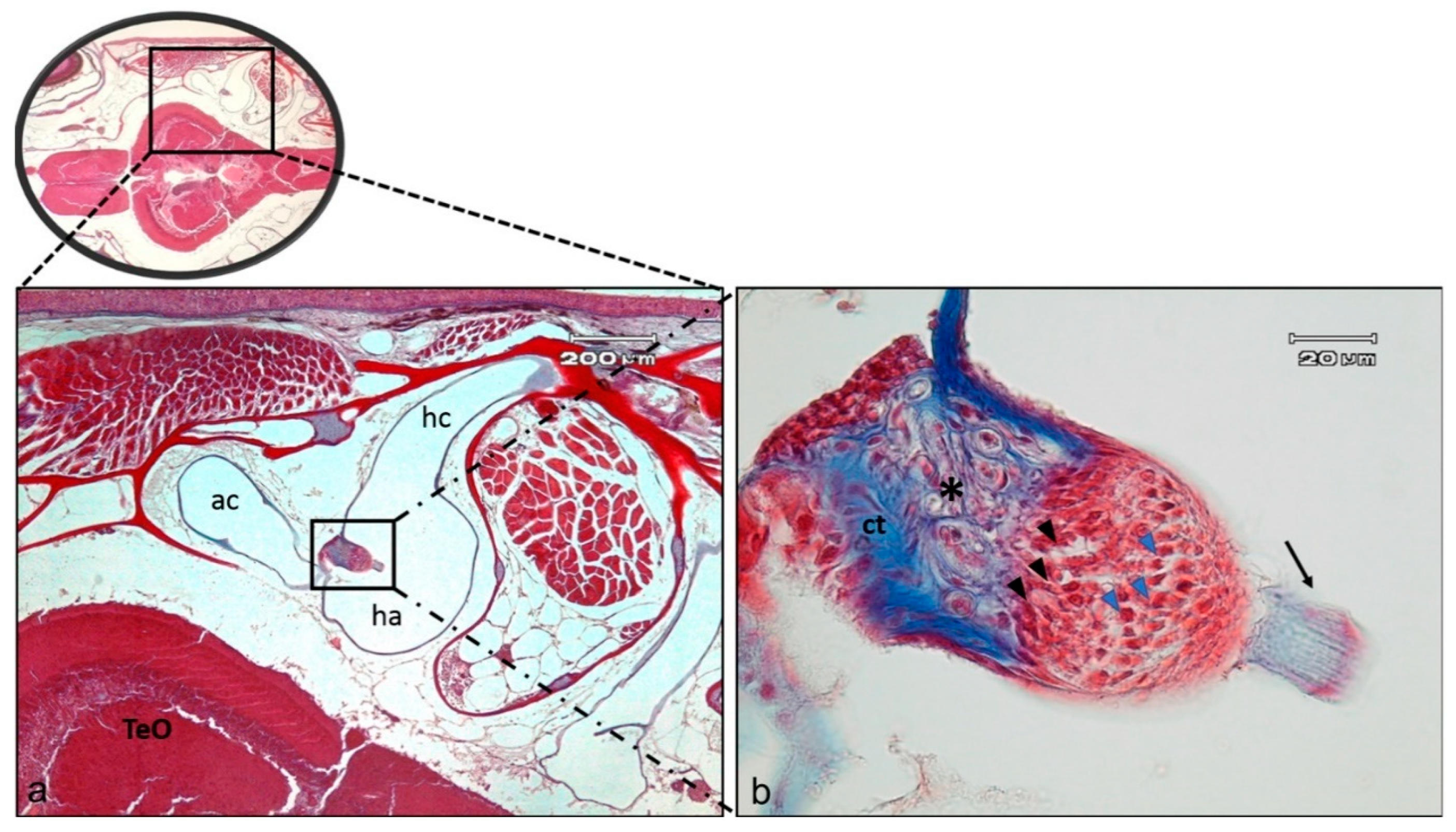
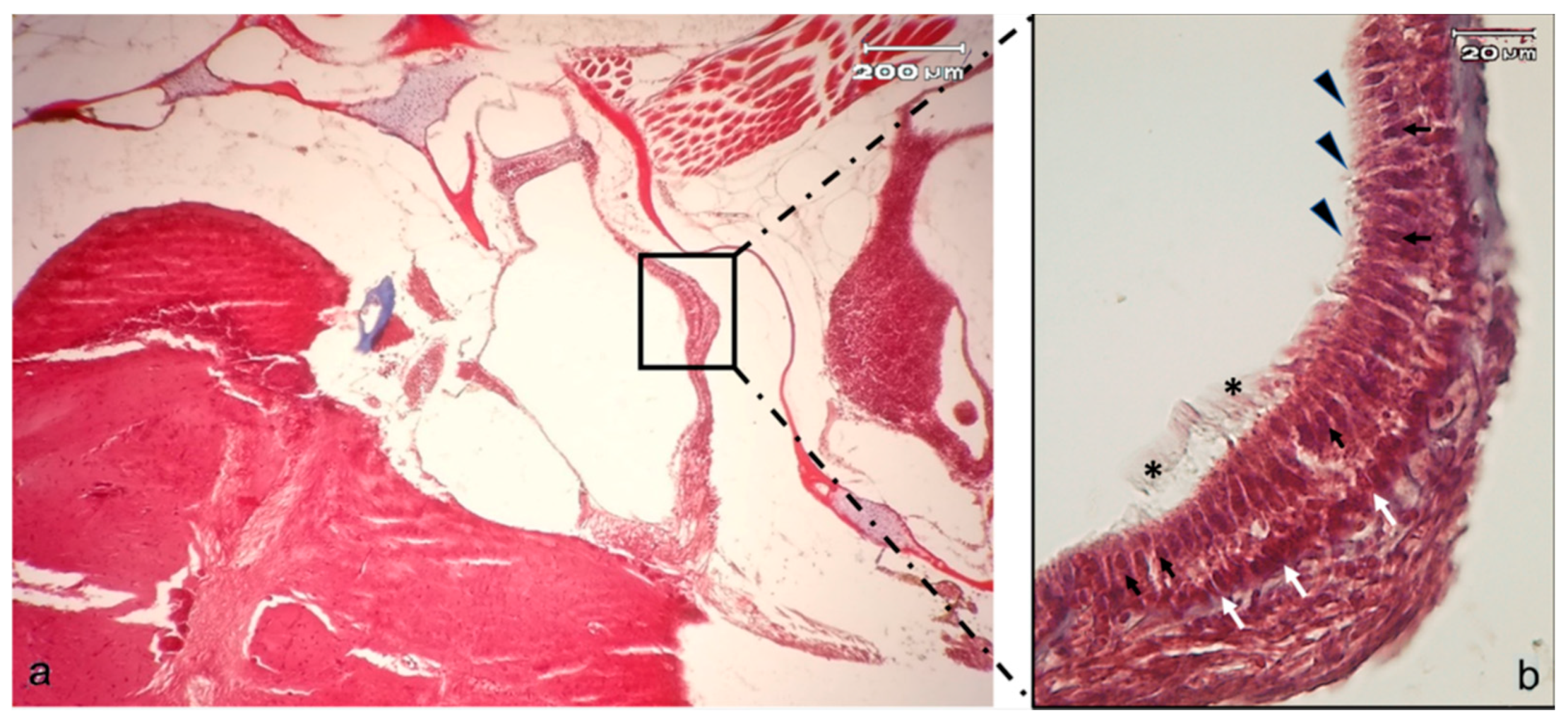
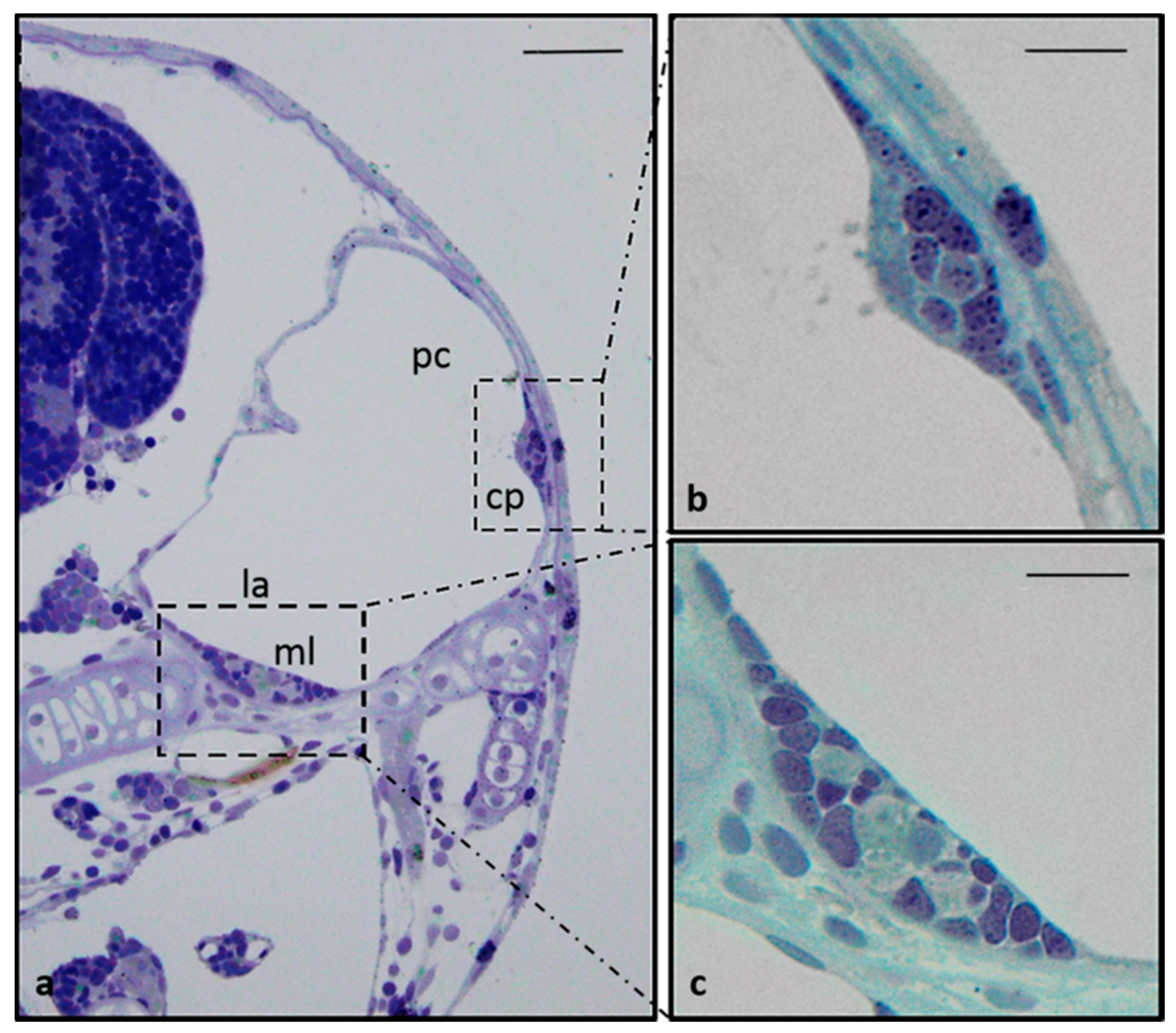
2.1. Anatomical Study of Zebrafish Inner Ear
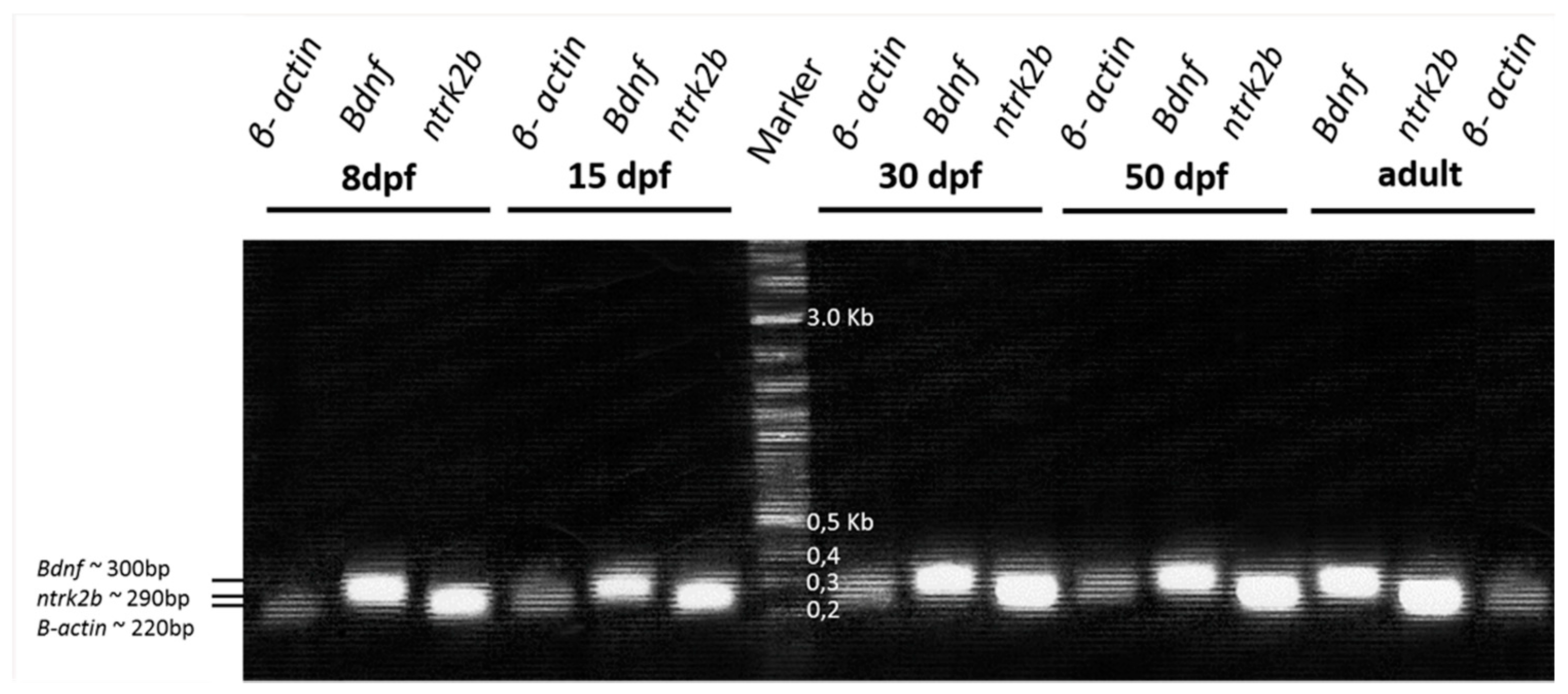
2.2. Expression and Occurrence of Bdnf and TrkB in Zebrafish Inner Ear
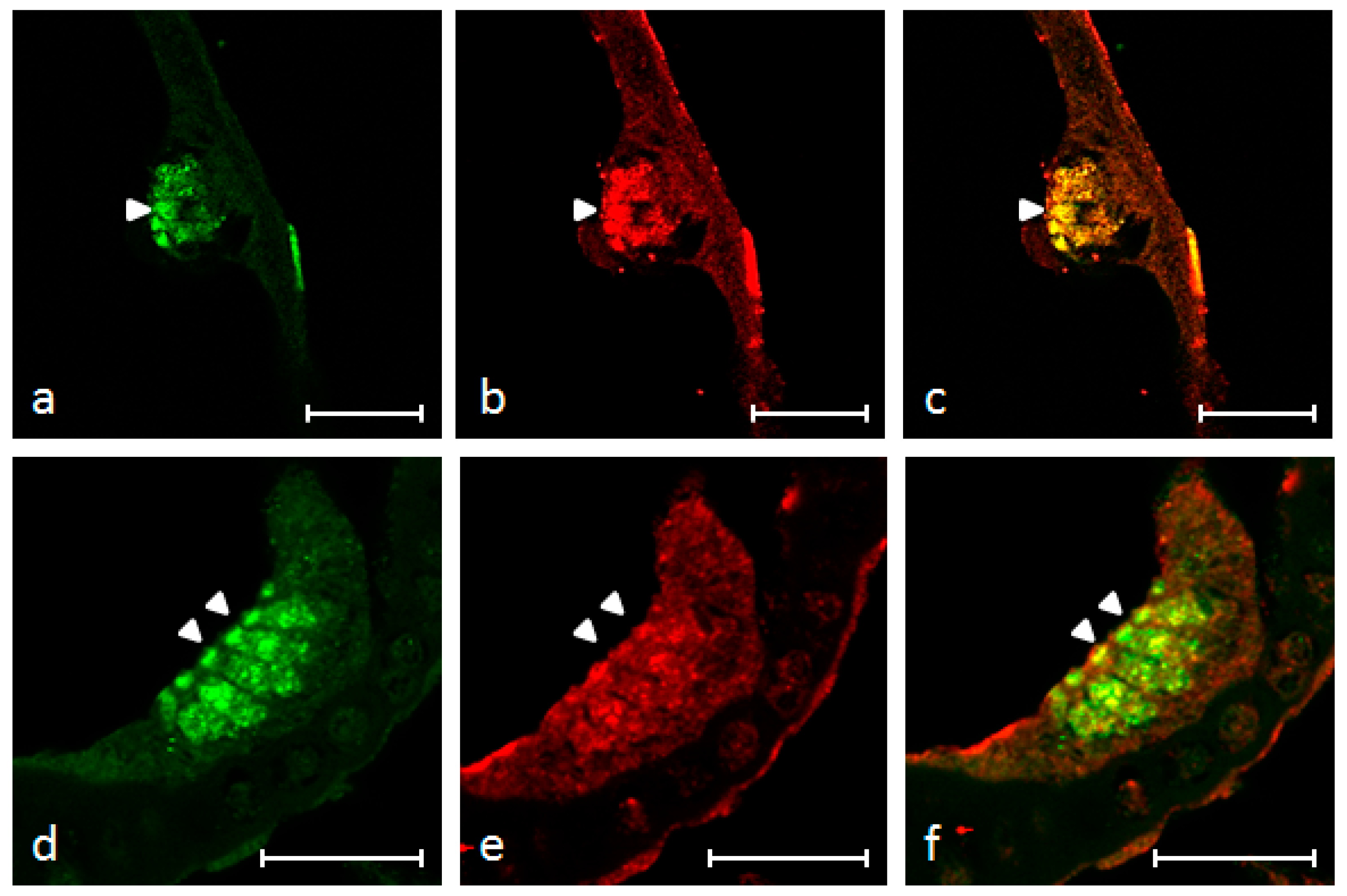
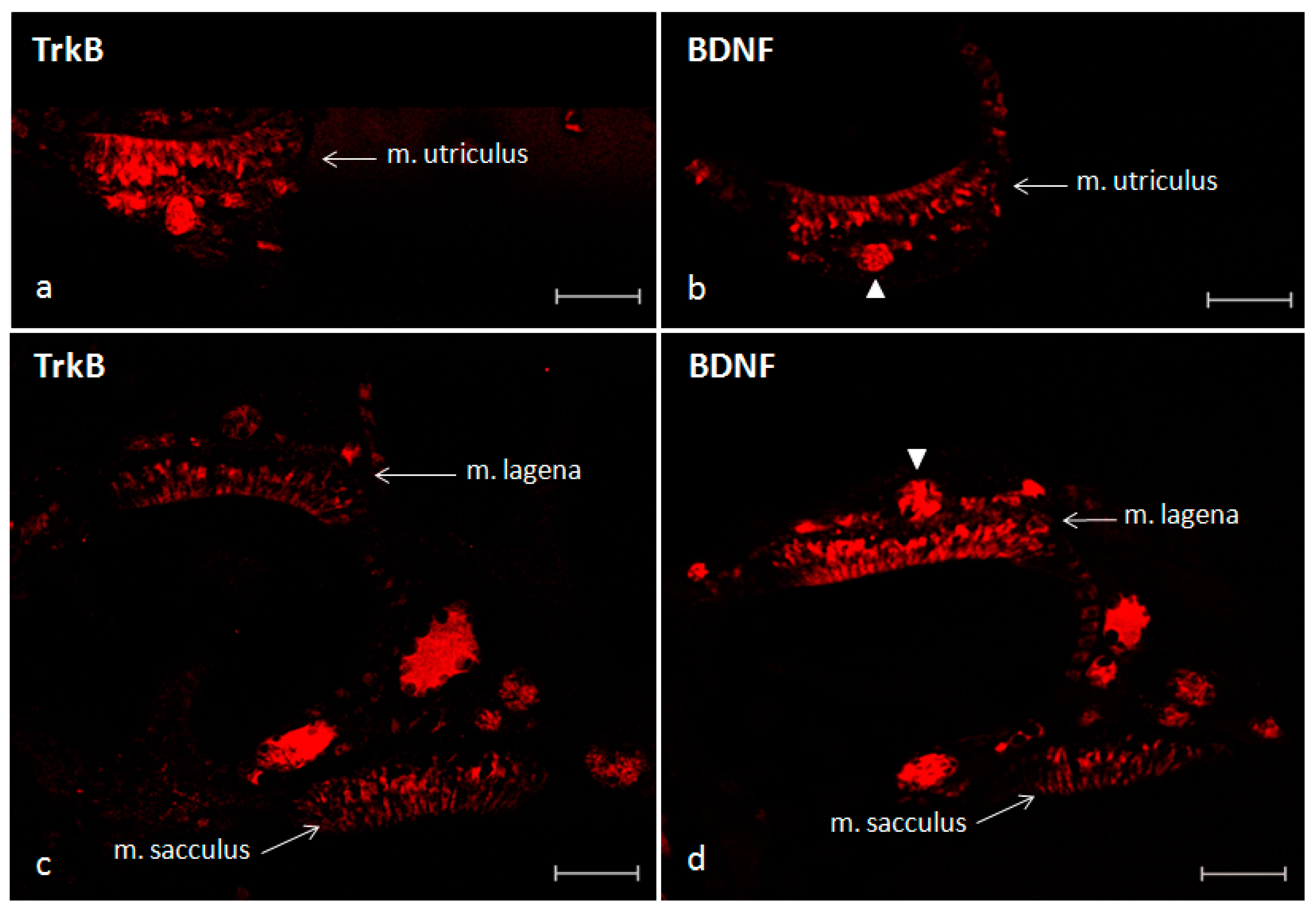
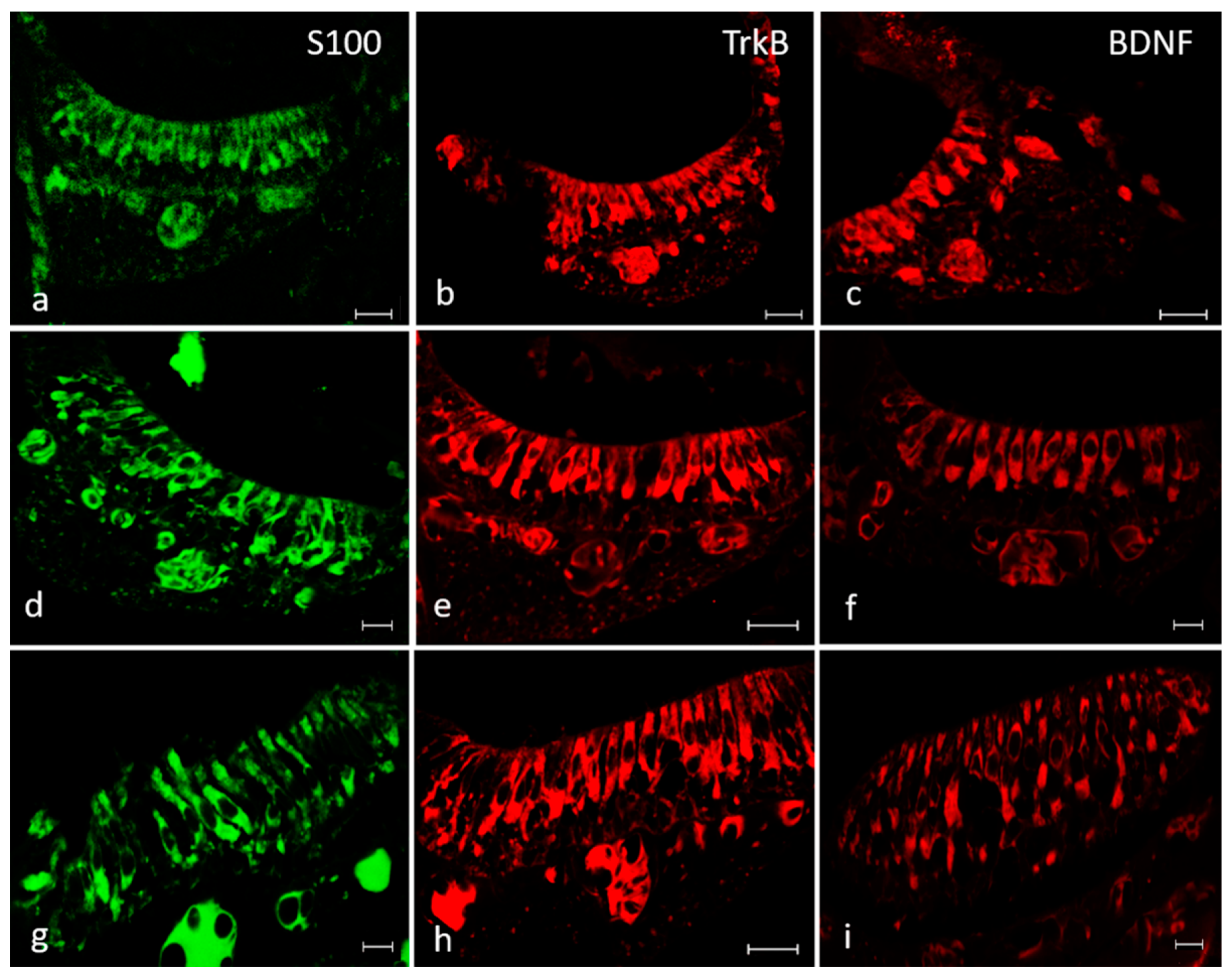
2.3. Immunofluorescence
3. Discussion
4. Materials and Methods
4.1. Zebrafish Breeding and Tissue Treatments
4.2. RT-PCR
4.3. Western Blot
4.4. Localization of BDNF, TrkB and S100 Protein Using Single and Double Immunofluorescence Staining
4.5. Scanning Electron Microscopy
Defleshing (Stripping)
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritzsch, B.; Tessarollo, L.; Coppola, E.; Reichardt, L.F. Neurotrophins in the ear: Their roles in sensory neuron survival and fiber guidance. Prog. Brain Res. 2004, 146, 265–278. [Google Scholar] [PubMed]
- Hannestad, J.; Marino, F.; Germanà, A.; Catania, S.; Abbate, F.; Ciriaco, E.; Vega, J. Trk neurotrophin receptor-like proteins in the teleost Dicentrarchus labrax. Cell Tissue Res. 2000, 300, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Germanà, A.; Catania, S.; Cavallaro, M.; González-Martínez, T.; Ciriaco, E.; Hannestad, J.; Vega, J. Immunohistochemical localization of BDNF-, TrkB- and TrkA-like proteins in the teleost lateral line system. J. Anat. 2002, 200, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Catania, S.; Germana, A.; Cabo, R.; Ochoa-Erena, F.; Guerrera, M.; Hannestad, J.; Represa, J.; Vega, J. Neurotrophin and Trk neurotrophin receptors in the inner ear of Salmo salar and Salmo trutta. J. Anat. 2007, 210, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Hallböök, F. Evolution of the vertebrate neurotrophin and Trk receptor gene families. Curr. Opin. Neurobiol. 1999, 9, 616–621. [Google Scholar] [CrossRef]
- Lucini, C.; D’Angelo, L.; Cacialli, P.; Palladino, A.; de Girolamo, P. BDNF, Brain, and Regeneration: Insights from Zebrafish. Int. J. Mol. Sci. 2018, 19, 3155. [Google Scholar] [CrossRef] [Green Version]
- Montalbano, G.; Mania, M.; Guerrera, M.C.; Abbate, F.; Laurà, R.; Navarra, M.; Vega, J.A.; Ciriaco, E.; Germanà, A. Morphological differences in adipose tissue and changes in BDNF/Trkb expression in brain and gut of a diet induced obese zebrafish model. Ann. Anat. 2016, 204, 36–44. [Google Scholar] [CrossRef]
- Cacialli, P.; Gueguen, M.-M.; Coumailleau, P.; D’Angelo, L.; Kah, O.; Lucini, C.; Pellegrini, E. BDNF expression in larval and adult zebrafish brain: Distribution and cell identification. PLoS ONE 2016, 11, e0158057. [Google Scholar] [CrossRef] [Green Version]
- De Felice, E.; Porreca, I.; Alleva, E.; De Girolamo, P.; Ambrosino, C.; Ciriaco, E.; Germanà, A.; Sordino, P. Localization of BDNF expression in the developing brain of zebrafish. J. Anat. 2014, 224, 564–574. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Levanti, M.B.; Germanà, G.P.; Navarra, M.; Laurà, R.; Vega, J.; Ciriaco, E.; Germanà, A. Expression and anatomical distribution of TrkB in the encephalon of the adult zebrafish (Danio rerio). Neurosci. Lett. 2014, 563, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ramos, C.; Bonnin-Arias, C.; Guerrera, M.C.; Calavia, M.; Chamorro, E.; Montalbano, G.; López-Velasco, S.; López-Muñiz, A.; Germanà, A.; Vega, J.A. Light regulates the expression of the BDNF/TrkB system in the adult zebrafish retina. Microsc. Res. Tech. 2013, 76, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Germanà, A.; Laurà, R.; Montalbano, G.; Guerrera, M.C.; Amato, V.; Zichichi, R.; Campo, S.; Ciriaco, E.; Vega, J. Expression of brain-derived neurotrophic factor and TrkB in the lateral line system of zebrafish during development. Cell. Mol. Neurobiol. 2010, 30, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Germana, A.; Abbate, F.; González-Martínez, T.; del Valle, M.; de Carlos, F.; Germanà, G.; Vega, J. S100 protein is a useful and specific marker for hair cells of the lateral line system in postembryonic zebrafish. Neurosci. Lett. 2004, 365, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Bang, P.I.; Sewell, W.F.; Malicki, J.J. Morphology and cell type heterogeneities of the inner ear epithelia in adult and juvenile zebrafish (Danio rerio). J. Comp. Neurol. 2001, 438, 173–190. [Google Scholar] [CrossRef]
- Monroe, J.D.; Rajadinakaran, G.; Smith, M.E. Sensory hair cell death and regeneration in fishes. Front. Cell. Neurosci. 2015, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.A.; Cheng, A.G.; Cunningham, L.L.; MacDonald, G.; Raible, D.W.; Rubel, E.W. Neomycin-induced hair cell death and rapid regeneration in the lateral line of zebrafish (Danio rerio). J. Assoc. Res. Otolaryngol. 2003, 4, 219–234. [Google Scholar] [CrossRef]
- Owens, K.N.; Santos, F.; Roberts, B.; Linbo, T.; Coffin, A.B.; Knisely, A.J.; Simon, J.A.; Rubel, E.W.; Raible, D.W. Identification of genetic and chemical modulators of zebrafish mechanosensory hair cell death. PLoS Genet. 2008, 4, e1000020. [Google Scholar] [CrossRef] [Green Version]
- Nicolson, T. The genetics of hair-cell function in zebrafish. J. Neurogenet. 2017, 31, 102–112. [Google Scholar] [CrossRef]
- Blanco-Sánchez, B.; Clément, A.; Phillips, J.; Westerfield, M. Zebrafish models of human eye and inner ear diseases. In Method Cell Biology; Academic Press: Cambridge, MA, USA, 2017; Volume 138, pp. 415–467. [Google Scholar]
- Rubel, E.W.; Furrer, S.A.; Stone, J.S. A brief history of hair cell regeneration research and speculations on the future. Hear. Res. 2013, 297, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Montalbano, G.; Abbate, F.; Levanti, M.B.; Germanà, G.P.; Laurà, R.; Ciriaco, E.; Vega, J.A.; Germanà, A. Topographical and drug specific sensitivity of hair cells of the zebrafish larvae to aminoglycoside-induced toxicity. Ann. Anat. 2014, 196, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Montalbano, G.; Capillo, G.; Laurà, R.; Abbate, F.; Levanti, M.; Guerrera, M.C.; Ciriaco, E.; Germanà, A. Neuromast hair cells retain the capacity of regeneration during heavy metal exposure. Ann. Anat. 2018, 218, 183–189. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Bao, B.; Li, H. Using zebrafish as a model to study the role of epigenetics in hearing loss. Expert Opin. Drug Discov. 2017, 12, 967–975. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Chai, R.; Li, H. Hair Cell Regeneration. Adv. Exp. Med. Biol. 2019, 1130, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Montalbano, G.; Mania, M.; Guerrera, M.C.; Laurà, R.; Abbate, F.; Levanti, M.; Maugeri, A.; Germanà, A.; Navarra, M. Effects of a Flavonoid-Rich Extract from Citrus sinensis Juice on a Diet-Induced Obese Zebrafish. Int. J. Mol. Sci. 2019, 20, 5116. [Google Scholar] [CrossRef] [Green Version]
- Germanà, A.; Marino, F.; Guerrera, M.C.; Campo, S.; De Girolamo, P.; Montalbano, G.; Germanà, G.P.; Ochoa-Erena, F.J.; Ciriaco, E.; Vega, J. Expression and distribution of S100 protein in the nervous system of the adult zebrafish (Danio rerio). Microsc. Res. Tech. 2008, 71, 248–255. [Google Scholar] [CrossRef]
- Germanà, A.; Paruta, S.; Germanà, G.P.; Ochoa-Erena, F.J.; Montalbano, G.; Cobo, J.; Vega, J.A. Differential distribution of S100 protein and calretinin in mechanosensory and chemosensory cells of adult zebrafish (Danio rerio). Brain Res. 2007, 1162, 48–55. [Google Scholar] [CrossRef]
- Germana’, A.; Montalbano, G.; Laura, R.; Ciriaco, E.; Del Valle, M.; Vega, J.A. S100 protein-like immunoreactivity in the crypt olfactory neurons of the adult zebrafish. Neurosci. Lett. 2004, 371, 196–198. [Google Scholar] [CrossRef]
- Abbate, F.; Catania, S.; Germana, A.; González, T.; Diaz-Esnal, B.; Germana, G.; Vega, J. S-100 protein is a selective marker for sensory hair cells of the lateral line system in teleosts. Neurosci. Lett. 2002, 329, 133–136. [Google Scholar] [CrossRef]
- Guerrera, M.C.; Montalbano, G.; Germana, A.; Maricchiolo, G.; Ciriaco, E.; Abbate, F. Morphology of the tongue dorsal surface in white sea bream (Diplodus sargus sargus). Acta Zool. 2015, 96, 236–241. [Google Scholar] [CrossRef]
- Germanà, A.; Sánchez-Ramos, C.; Guerrera, M.C.; Calavia, M.; Navarro, M.; Zichichi, R.; García-Suárez, O.; Pérez-Piñera, P.; Vega, J.A. Expression and cell localization of brain-derived neurotrophic factor and TrkB during zebrafish retinal development. J. Anat. 2010, 217, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Montcouquiol, M.; Valat, J.; Travo, C.; Sans, A. A role for BDNF in early postnatal rat vestibular epithelia maturation: Implication of supporting cells. Eur. J. Neurosci. 1998, 10, 598–606. [Google Scholar] [CrossRef]
- Pirvola, U.; Hallböök, F.; Xing-Qun, L.; Virkkala, J.; Saarma, M.; Ylikoski, J. Expression of neurotrophins and Trk receptors in the developing, adult, and regenerating avian cochlea. J. Neurobiol. 1997, 33, 1019–1033. [Google Scholar] [CrossRef]
- Don, D.M.; Newman, A.N.; Micevych, P.E.; Popper, P. Expression of brain-derived neurotrophic factor and its receptor mRNA in the vestibuloauditory system of the bullfrog. Hear. Res. 1997, 114, 10–20. [Google Scholar] [CrossRef]
- Heinrich, G.; Lum, T. Fish neurotrophins and Trk receptors. Int. J. Dev. Neurosci. 2000, 18, 1–27. [Google Scholar] [CrossRef]
- Nittoli, V.; Sepe, R.M.; Coppola, U.; D’Agostino, Y.; De Felice, E.; Palladino, A.; Vassalli, Q.A.; Locascio, A.; Ristoratore, F.; Spagnuolo, A.; et al. A comprehensive analysis of neurotrophins and neurotrophin tyrosine kinase receptors expression during development of zebrafish. In J. Comp. Neurol.; 2018; Volume 526, pp. 1057–1072. [Google Scholar] [CrossRef]
- Coffin, A.B.; Ramcharitar, J. Chemical ototoxicity of the fish inner ear and lateral line. In Fish Hearing and Bioacoustics; Springer: Cham, Switzerland, 2016; pp. 419–437. [Google Scholar]
- Stawicki, T.M.; Esterberg, R.; Hailey, D.W.; Raible, D.W.; Rubel, E.W. Using the zebrafish lateral line to uncover novel mechanisms of action and prevention in drug-induced hair cell death. Front. Cell. Neurosci. 2015, 9, 46. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.D.; Raible, D.W. Distinct progenitor populations mediate regeneration in the zebrafish lateral line. eLife 2019, 8, e43736. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, S.; Wong, H.-T.C.; He, X.J.; Beirl, A.; Petralia, R.S.; Wang, Y.-X.; Kindt, K.S. Synaptically silent sensory hair cells in zebrafish are recruited after damage. Nat. Commun. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Thomas, J.; Gangappa, S.; Kanangat, S.; Rouse, B. On the essential involvement of neutrophils in the immunopathologic disease: Herpetic stromal keratitis. J. Immunol. 1997, 158, 1383–1391. [Google Scholar]
- Nicolson, T. The genetics of hearing and balance in zebrafish. Annu. Rev. Genet. 2005, 39, 9–22. [Google Scholar] [CrossRef]
- Shepherd, R.K.; Coco, A.; Epp, S.B.; Crook, J.M. Chronic depolarization enhances the trophic effects of brain-derived neurotrophic factor in rescuing auditory neurons following a sensorineural hearing loss. J. Comp. Neurol. 2005, 486, 145–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budenz, C.L.; Pfingst, B.E.; Raphael, Y. The use of neurotrophin therapy in the inner ear to augment cochlear implantation outcomes. Anat. Rec. 2012, 295, 1896–1908. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, N.; Schulze, J.; Warwas, D.P.; Ehlert, N.; Lenarz, T.; Warnecke, A.; Behrens, P. Long-term delivery of brain-derived neurotrophic factor (BDNF) from nanoporous silica nanoparticles improves the survival of spiral ganglion neurons in vitro. PLoS ONE 2018, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Laurà, R.; Abbate, F.; Germanà, G.; Montalbano, G.; Germanà, A.; Levanti, M. Fine structure of the canal neuromasts of the lateral line system in the adult zebrafish. Anat. Histol. Embryol. 2018, 47, 322–329. [Google Scholar] [CrossRef]
- Amato, V.; Vina, E.; Calavia, M.; Guerrera, M.C.; Laurà, R.; Navarro, M.; De Carlos, F.; Cobo, J.; Germanà, A.; Vega, J. TRPV4 in the sensory organs of adult zebrafish. Microsc. Res. Tech. 2012, 75, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Gier, R. Preparation of great horned owl skeleton. Trans. Kans. Acad. Sci. (1903-) 1951, 54, 344–345. [Google Scholar] [CrossRef]
- Mairs, S.; Swift, B.; Rutty, G.N. Detergent: An alternative approach to traditional bone cleaning methods for forensic practice. Am. J. Forensic Med. Pathol. 2004, 25, 276–284. [Google Scholar] [CrossRef]
- Modina, S.; Abbate, F.; Germana, G.P.; Lauria, A.; Luciano, A.M. β-Catenin localization and timing of early development of bovine embryos obtained from oocytes matured in the presence of follicle stimulating hormone. Anim. Reprod. Sci. 2007, 100, 264–279. [Google Scholar] [CrossRef]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germanà, A.; Guerrera, M.C.; Laurà, R.; Levanti, M.; Aragona, M.; Mhalhel, K.; Germanà, G.; Montalbano, G.; Abbate, F. Expression and Localization of BDNF/TrkB System in the Zebrafish Inner Ear. Int. J. Mol. Sci. 2020, 21, 5787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165787
Germanà A, Guerrera MC, Laurà R, Levanti M, Aragona M, Mhalhel K, Germanà G, Montalbano G, Abbate F. Expression and Localization of BDNF/TrkB System in the Zebrafish Inner Ear. International Journal of Molecular Sciences. 2020; 21(16):5787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165787
Chicago/Turabian StyleGermanà, Antonino, Maria Cristina Guerrera, Rosaria Laurà, Maria Levanti, Marialuisa Aragona, Kamel Mhalhel, Germana Germanà, Giuseppe Montalbano, and Francesco Abbate. 2020. "Expression and Localization of BDNF/TrkB System in the Zebrafish Inner Ear" International Journal of Molecular Sciences 21, no. 16: 5787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165787