Dermal Fibroblasts Internalize Phosphatidylserine-Exposed Secretory Melanosome Clusters and Apoptotic Melanocytes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
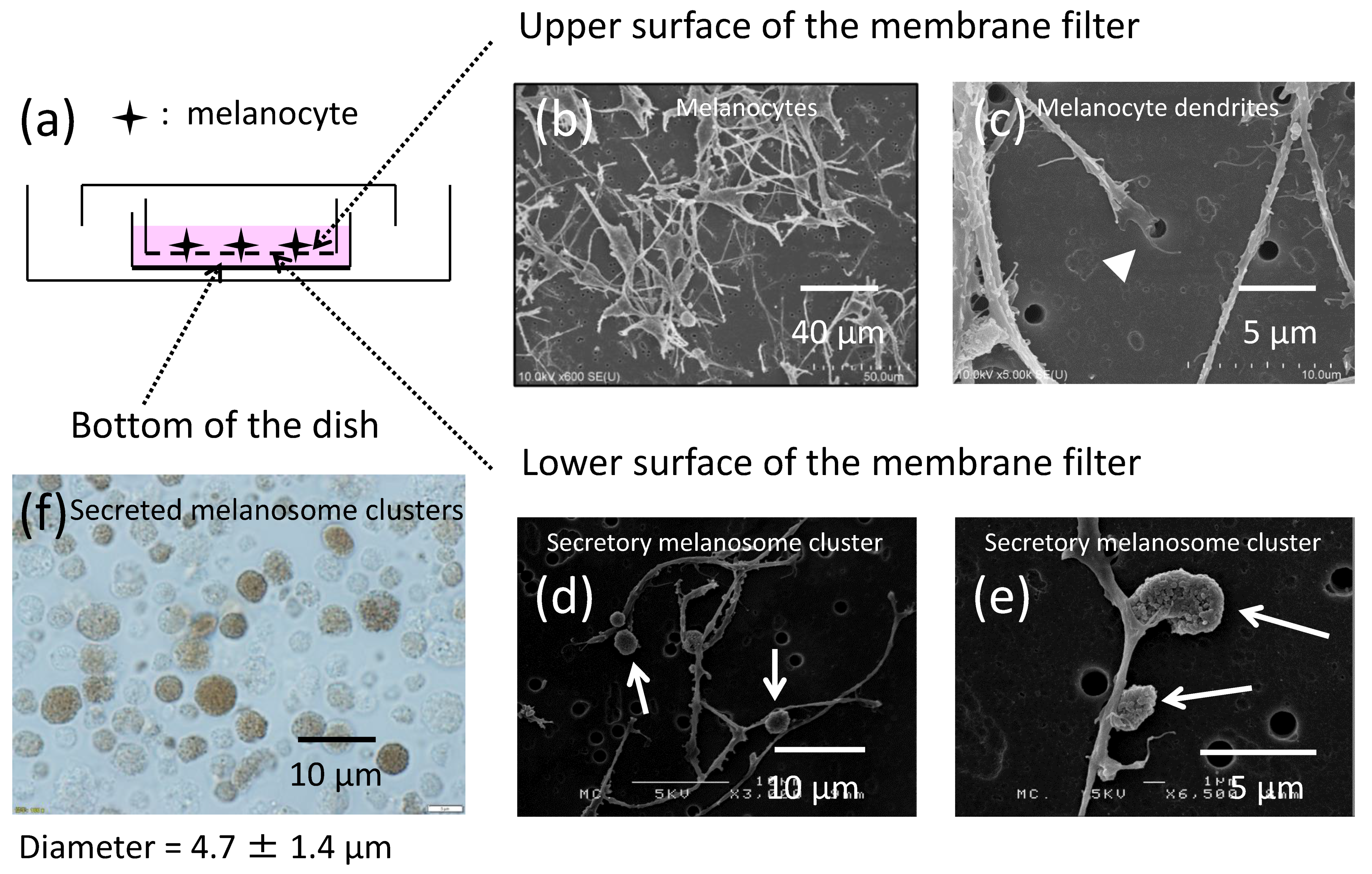
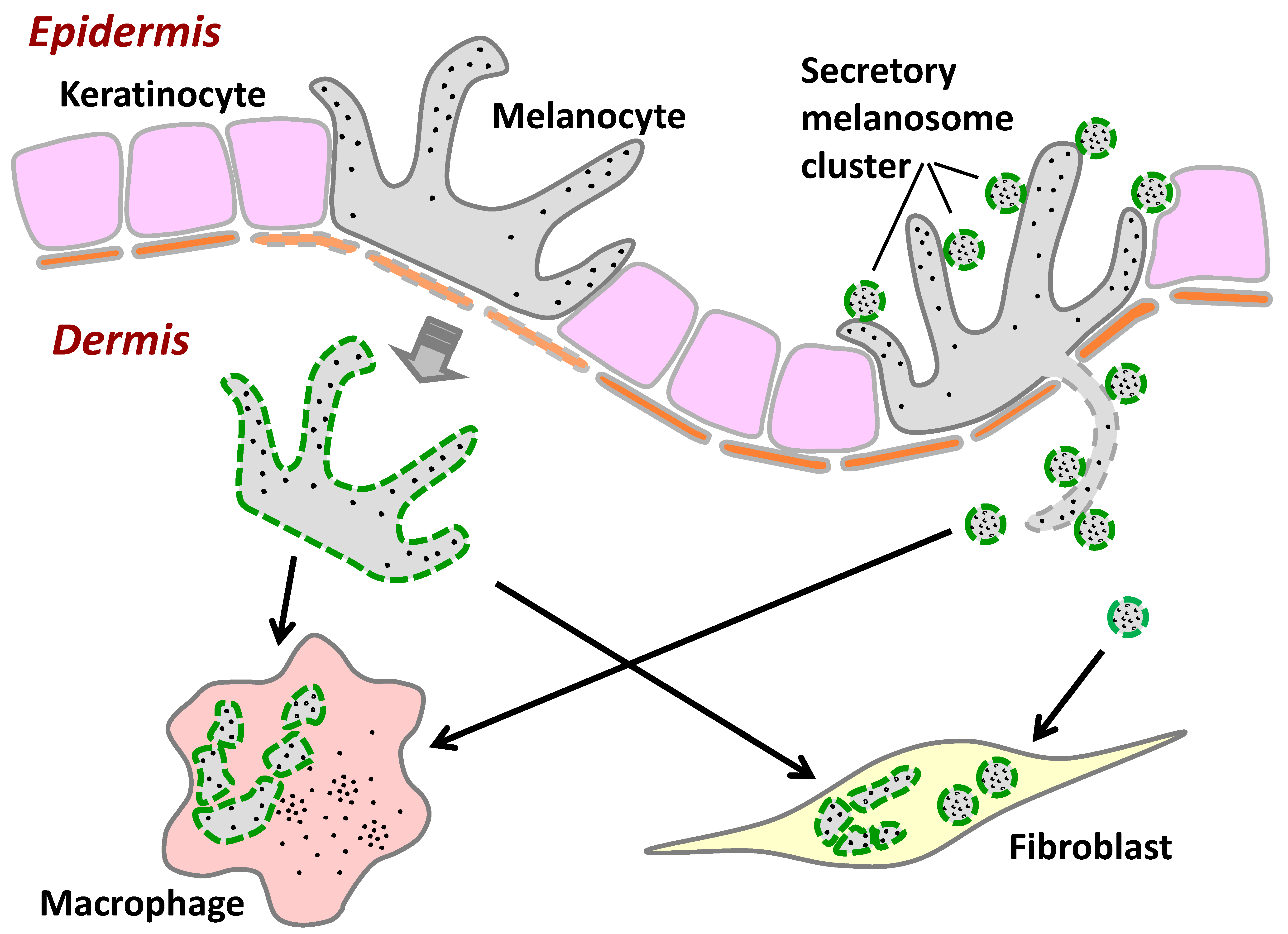
2.1. Melanocytes Produce Secretory Melanosome Clusters along Their Dendrites with Exposed Phosphatidylserine (PtdSer)
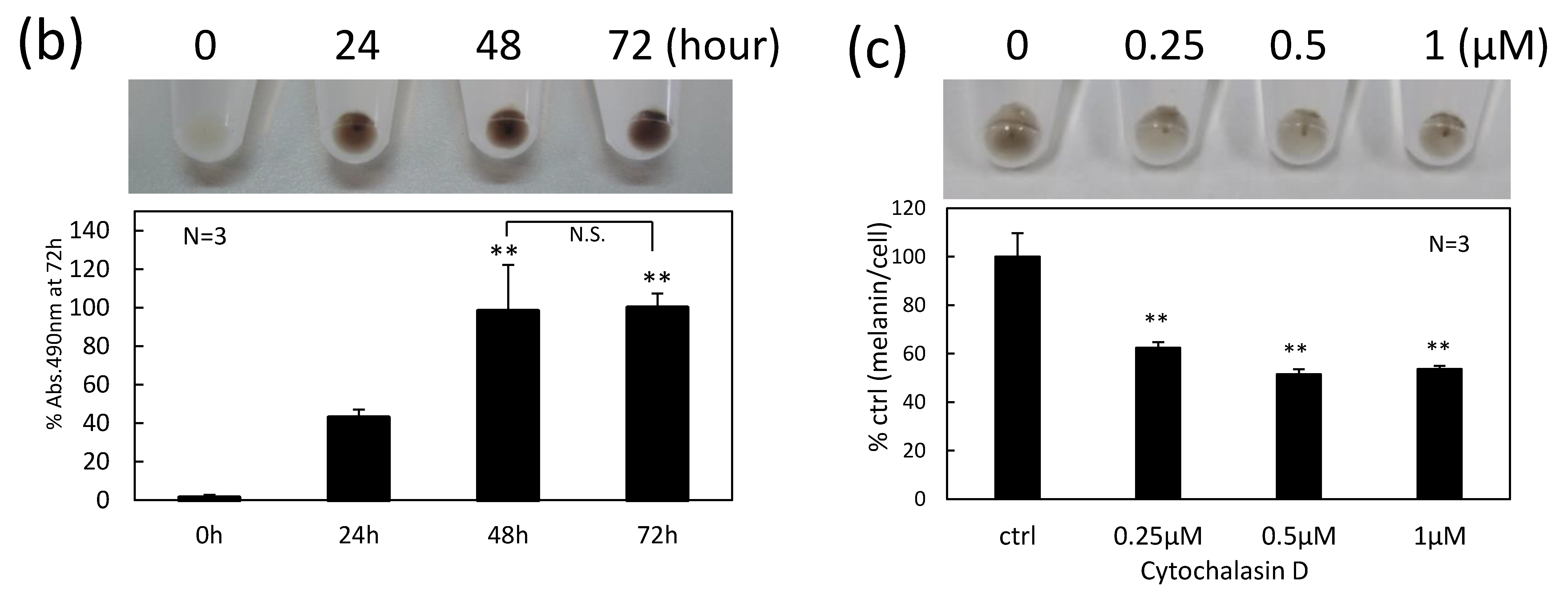
2.2. Fibroblasts Internalize Secreted Melanosome Clusters by Actin-Dependent Endocytosis
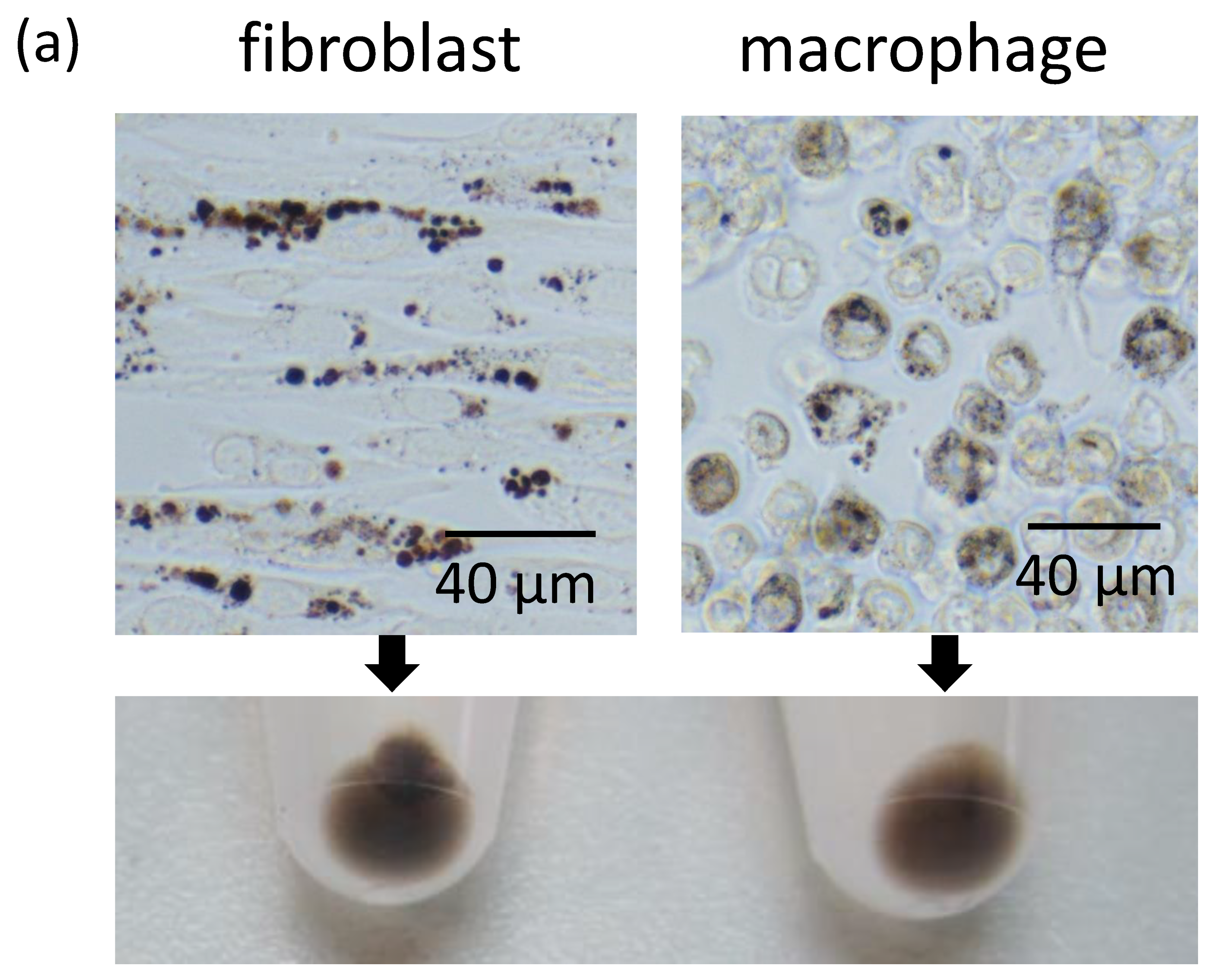
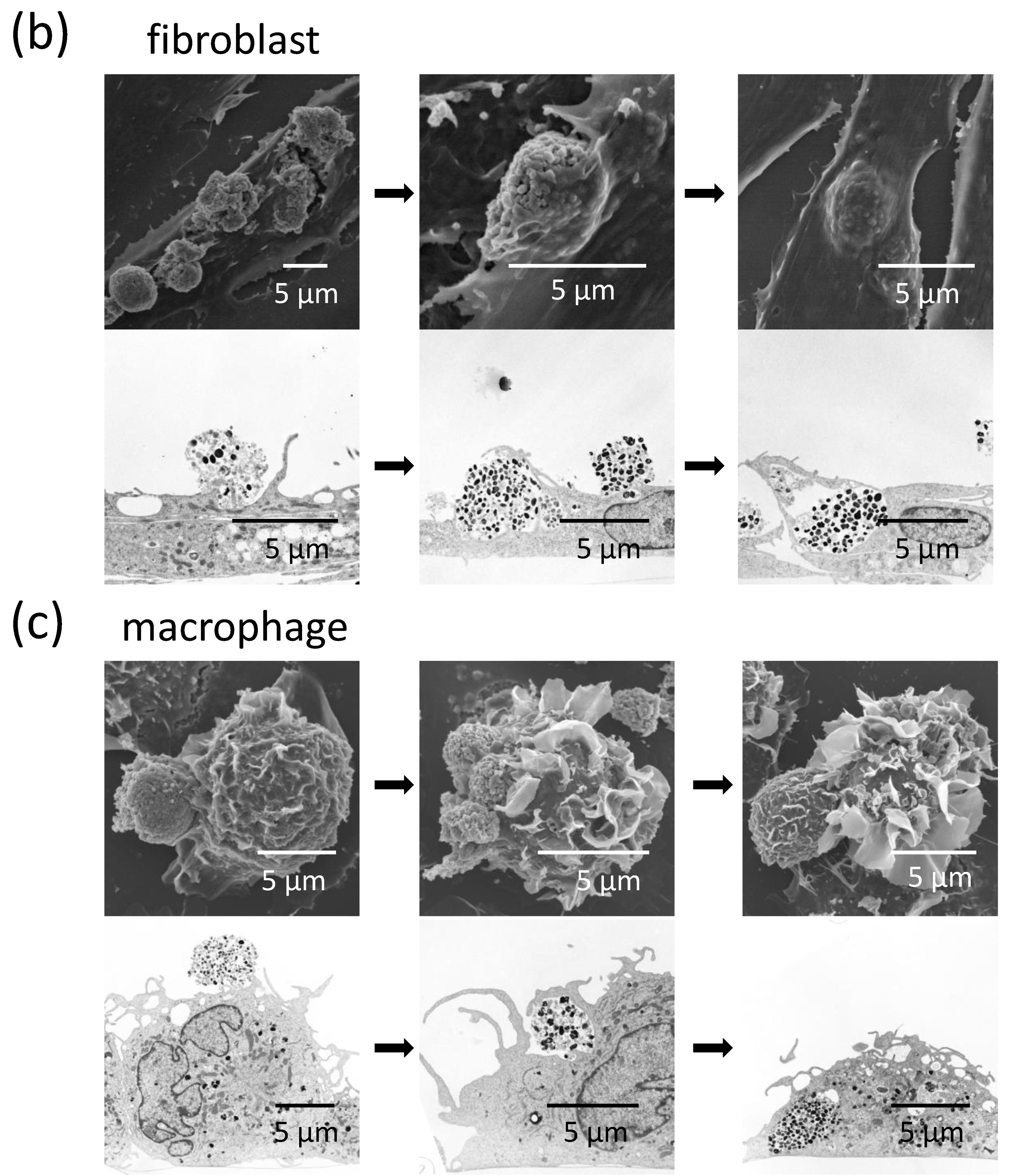
2.3. Fibroblasts Uptake Secreted Melanosome Clusters as do Macrophages
2.4. Fibroblasts Internalize Apoptotic Melanocytes
3. Discussion
4. Materials and Methods
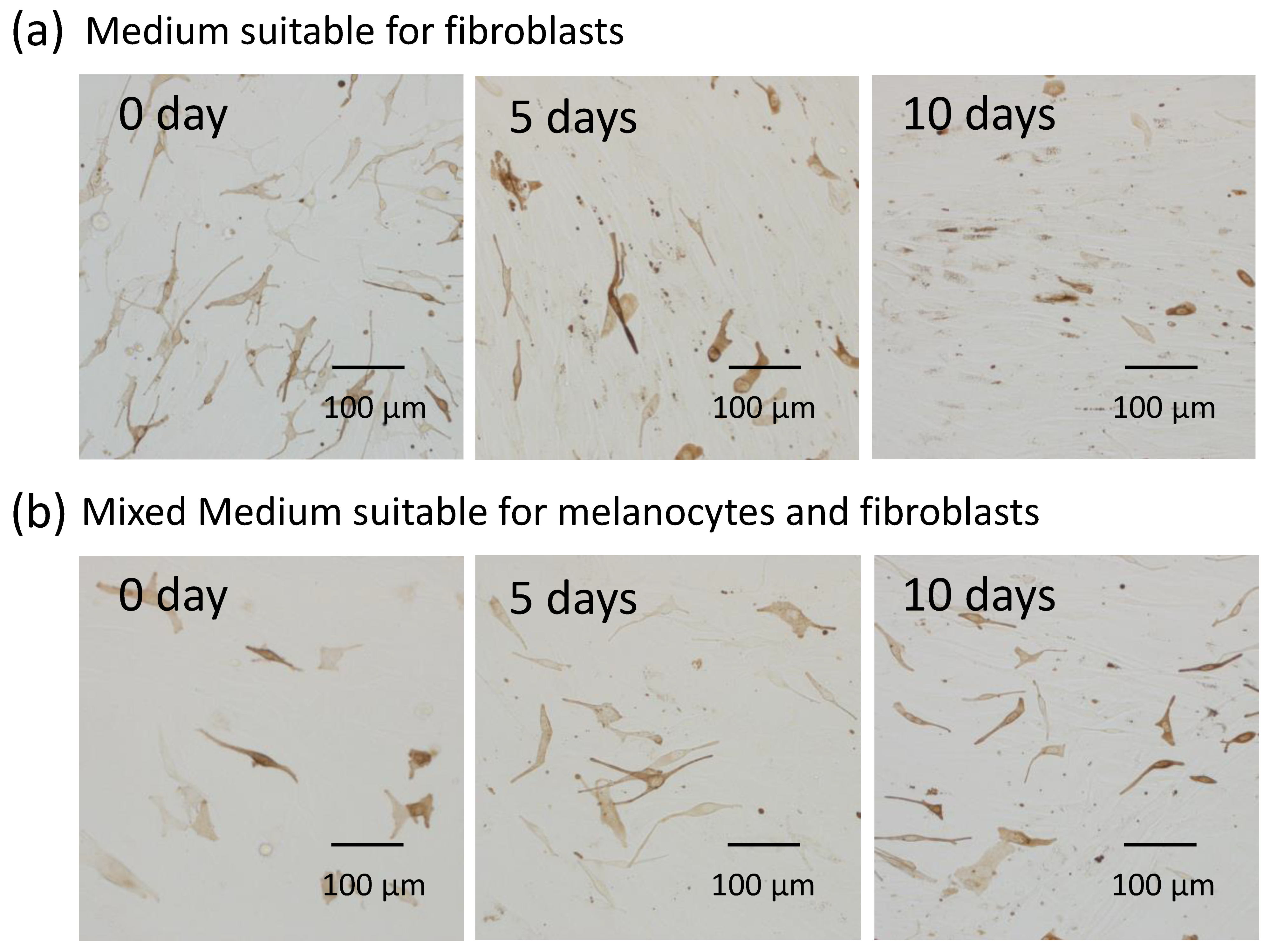
4.1. Cell Culture
4.2. PtdSer Staining of Secretory Melanosome Clusters and Apoptotic Melanocytes
4.3. Isolation and Treatment of Secreted Melanosome Clusters
4.4. Treatment with an Endocytosis Inhibitor
4.5. Detection of TIM4 in Fibroblasts
4.6. Electron Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| D-PBS | Ca2+ and Mg2+ free Dulbecco’s phosphate buffered saline |
| PtdSer | Phosphatidylserine |
| PMA | Phorbol 12-myristate 13-acetate |
| SEM | Scanning electron microscopy |
| TEM | Transmission electron microscopy |
References
- Wu, X.; Hammer, J.A. Melanosome transfer: It is best to give and receive. Curr. Opin. Cell Biol. 2014, 29, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun-Falco, O.; Schoefinius, H.H. Lentigo senilis. Hautarzt 1971, 22, 277–283. [Google Scholar] [PubMed]
- Hashmi, S.; Marinkovich, M.P. Molecular organization of the basement membrane zone. Clin. Dermatol. 2011, 29, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Nagao, S.; Iijima, S. Light and electron microscopic study of Riehl’s melanosis. Possible mode of its pigmentary incontinence. J. Cutan. Pathol. 1974, 1, 165–175. [Google Scholar] [CrossRef]
- Masu, S.; Seiji, M. Pigmentary incontinence in fixed drug eruptions. Histologic and electron microscopic findings. J. Am. Acad. Dermatol. 1983, 8, 525–532. [Google Scholar] [CrossRef]
- Barros, H.R.; Almeida, J.R.; Mattos e Dinato, S.L.; Sementilli, A.; Romiti, N. Lichen planus pigmentosus inversus. An. Bras. Dermatol. 2013, 88, 146–149. [Google Scholar] [CrossRef] [Green Version]
- Ishii, M.; Terao, Y.; Asai, Y.; Hamada, T. High melanosome engulfing activity of cutaneous fibroblasts in macular amyloidosis: An electron microscopic study. J. Cutan. Pathol. 1984, 11, 476–484. [Google Scholar] [CrossRef]
- Ohkuma, M. Presence of melanophages in the normal Japanese skin. Am. J. Dermatopathol. 1991, 13, 32–37. [Google Scholar] [CrossRef]
- Ünver, N.; Freyschmidt-Paul, P.; Hörster, S.; Wenck, H.; Stäb, F.; Blatt, T.; Elsässer, H.P. Alterations in the epidermal-dermal melanin axis and factor XIIIa melanophages in senile lentigo and ageing skin. Brit. J. Dermatol. 2006, 155, 119–128. [Google Scholar] [CrossRef]
- Tanemura, A.; Yang, L.; Yang, F.; Nagata, Y.; Wataya-Kaneda, M.; Fukai, K.; Tsuruta, D.; Ohe, R.; Yamakawa, M.; Suzuki, T.; et al. An immune pathological and ultrastructural skin analysis for rhododenol-induced leukoderma patients. J. Dermatol. Sci. 2015, 77, 185–188. [Google Scholar] [CrossRef]
- Ando, H.; Niki, Y.; Yoshida, M.; Ito, M.; Akiyama, K.; Kim, J.H.; Yoon, T.J.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Involvement of pigment globules containing multiple melanosomes in the transfer of melanosomes from melanocytes to keratinocytes. Cell. Logist. 2011, 1, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasraee, B.; Pataky, M.; Nikolic, D.S.; Carraux, P.; Piguet, V.; Salomon, D.; Sorg, O.; Saurat, J.H. A new spectrophotometric method for simple quantification of melanosomal transfer from melanocytes to keratinocytes. Exp. Dermatol. 2011, 20, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes are transferred from melanocytes to keratinocytes through the processes of packaging, release, uptake, and dispersion. J. Investig. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef] [Green Version]
- Scott, G. Demonstration of melanosome transfer by a shedding microvesicle mechanism. J. Investig. Dermatol. 2012, 132, 1073–1074. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.S.; Masedunskas, A.; Weigert, R.; Copeland, N.G.; Jenkins, N.A.; Hammer, J.A. Melanoregulin regulates a shedding mechanism that drives melanosome transfer from melanocytes to keratinocytes. Proc. Natl. Acad. Sci. USA 2012, 109, E2101–E2109. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.J.; Ma, H.Y.; Yang, Y.; Li, P.C.; Zi, S.X.; Jia, C.Y.; Chen, R. α-Melanocyte stimulating hormone (MSH) and prostaglandin E2 (PGE2) drive melanosome transfer by promoting filopodia delivery and shedding spheroid granules: Evidence from atomic force microscopy observation. J. Dermatol. Sci. 2014, 76, 222–230. [Google Scholar] [CrossRef]
- Andree, H.A.; Reutelingsperger, C.P.; Hauptmann, R.; Hemker, H.C.; Hermens, W.T.; Willems, G.M. Binding of vascular anticoagulant alpha (VAC alpha) to planar phospholipid bilayers. J. Biol. Chem. 1990, 265, 4923–4928. [Google Scholar] [PubMed]
- van Engeland, M.; Nieland, L.J.; Ramaekers, F.C.; Schutte, B.; Reutelingsperger, C.P. Annexin V-affinity assay: A review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry 1998, 31, 1–9. [Google Scholar] [CrossRef]
- Nagata, S.; Suzuki, J.; Segawa, K.; Fujii, T. Exposure of phosphatidylserine on the cell surface. Cell Death Differ. 2016, 23, 952–961. [Google Scholar] [CrossRef] [Green Version]
- Miyanishi, M.; Tada, K.; Koike, M.; Uchiyama, Y.; Kitamura, T.; Nagata, S. Identification of Tim4 as a phosphatidylserine receptor. Nature 2007, 450, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Nishi, C.; Toda, S.; Segawa, K.; Nagata, S. Tim4- and MerTK-mediated engulfment of apoptotic cells by mouse resident peritoneal macrophages. Mol. Cell. Biol. 2014, 34, 1512–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, H.; Nishii, Y.; Yamashita, K.; Kawamata, S.; Yoshikawa, K. The uptake and long-term storage of india ink particles and latex beads by fibroblasts in the dermis and subcutis of mice, with special regard to the non-inflammatory defense reaction by fibroblasts. Arch. Histol. Cytol. 1988, 51, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aspengren, S.; Hedberg, D.; Wallin, M. Studies of pigment transfer between Xenopus laevis melanophores and fibroblasts in vitro and in vivo. Pigment Cell Res. 2006, 19, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dror, S.; Sander, L.; Schwartz, H.; Sheinboim, D.; Barzilai, A.; Dishon, Y.; Apcher, S.; Golan, T.; Greenberger, S.; Barshack, I.; et al. Melanoma miRNA trafficking controls tumour primary niche formation. Nat. Cell Biol. 2016, 18, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Oakford, M.E.; Dixon, S.V.; August, S.; Pickard, C.; Ardern-Jones, M.; Lackie, P.; Friedmann, P.S.; Healy, E. Migration of immunocytes across the basement membrane in skin: The role of basement membrane pores. J. Investig. Dermatol. 2011, 131, 1950–1953. [Google Scholar] [CrossRef] [Green Version]
- Moretti, S.; Nassini, R.; Prignano, F.; Pacini, A.; Materazzi, S.; Naldini, A.; Simoni, A.; Baroni, G.; Pellerito, S.; Filippi, I.; et al. Protease-activated receptorF downregulation is associated to vitiligo lesions. Pigment Cell Melanoma Res. 2009, 22, 335–338. [Google Scholar] [CrossRef]
- Sharlow, E.R.; Paine, C.S.; Babiarz, L.; Eisinger, M.; Shapiro, S.; Seiberg, M. The protease-activated receptor-2 upregulates keratinocyte phagocytosis. J. Cell Sci. 2000, 113, 3093–3101. [Google Scholar]
- Fadok, V.A.; Voelker, D.R.; Campbell, P.A.; Cohen, J.J.; Bratton, D.L.; Henson, P.M. Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. J. Immunol. 1992, 148, 2207–2216. [Google Scholar]
- Tadokoro, R.; Murai, H.; Sakai, K.I.; Okui, T.; Yokota, Y.; Takahashi, Y. Melanosome transfer to keratinocyte in the chicken embryonic skin is mediated by vesicle release associated with Rho-regulated membrane blebbing. Sci. Rep. 2016, 6, 38277. [Google Scholar] [CrossRef]
- Conner, S.D.; Schmid, S.L. Regulated portals of entry into the cell. Nature 2003, 422, 37–44. [Google Scholar] [CrossRef]
- Kaksonen, M.; Toret, C.P.; Drubin, D.G. Harnessing actin dynamics for clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2006, 7, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Donaldson, J.G. Search for inhibitors of endocytosis: Intended specificity and unintended consequences. Cell. Logist. 2012, 2, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641–24654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.K.; Kurfurst, R.; Nizard, C.; Schnebert, S.; Perrier, E.; Tobin, D.J. Melanin transfer in human skin cells is mediated by filopodia—a model for homotypic and heterotypic lysosome-related organelle transfer. FASEB J. 2010, 24, 3756–3769. [Google Scholar] [CrossRef] [PubMed]
- Tarafder, A.K.; Bolasco, G.; Correia, M.S.; Pereira, F.J.C.; Iannone, L.; Hume, A.N.; Kirkpatrick, N.; Picardo, M.; Torrisi, M.R.; Rodrigues, I.P.; et al. Rab11b mediates melanin transfer between donor melanocytes and acceptor keratinocytes via coupled exo/endocytosis. J. Investig. Dermatol. 2014, 134, 1056–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.; Paus, R.; Schadendorf, D. Melanocytes as “sensory” and regulatory cells in the epidermis. J. Theor. Biol. 1993, 164, 103–120. [Google Scholar] [CrossRef]
- Slominski, A.; Kim, T.-K.; Brozyna, A.A.; Janjetovic, Z.; Brooks, D.L.P.; Schwab, L.P.; Skobowiat, C.; Jozwicki, W.; Seagroves, T.N. The role of melanogenesis in regulation of melanoma behavior: Melanogenesis leads to stimulation of HIF-1α expression and HIF-dependent attendant pathways. Arch. Biochem. Biophys. 2014, 563, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Okuma, M.; Seiji, M. Lymphatic transport of melanosomes to the lymph node. Tohoku J. Exp. Med. 1973, 111, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Grossi, A.B.; Hyttel, P.; Jensen, H.E.; Leifsson, P.S. Porcine melanotic cutaneous lesions and lymph nodes: Immunohistochemical differentiation of melanocytes and melanophages. Vet. Pathol. 2015, 52, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Hass, R.; Bartels, H.; Topley, N.; Hadam, M.; Köhler, L.; Goppelt-Strübe, M.; Resch, K. TPA-induced differentiation and adhesion of U937 cells: Change in ultrastructure, cytoskeletal organization and expression of cell surface antigens. Eur. J. Cell Biol. 1989, 48, 282–293. [Google Scholar]
- García, A.; Serrano, A.; Abril, E.; Jimenez, P.; Real, L.M.; Cantón, J.; Garrido, F.; Ruiz-Cabello, F. Differential effect on U937 cell differentiation by targeting transcriptional factors implicated in tissue- or stage-specific induced integrin expression. Exp. Hematol. 1999, 27, 353–364. [Google Scholar] [CrossRef]
- Ando, H.; Ohagi, Y.; Yoshida, M.; Yoshimoto, S.; Higashi, Y.; Yagi, M.; Monji, K.; Yagi, M.; Uchiumi, T.; Kang, D.; et al. Melanin pigment interrupts the fluorescence staining of mitochondria in melanocytes. J. Dermatol. Sci. 2016, 84, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Hiroyasu, S.; Ozawa, T.; Kobayashi, H.; Ishii, M.; Aoyama, Y.; Kitajima, Y.; Hashimoto, T.; Jones, J.C.; Tsuruta, D. Bullous pemphigoid IgG induces BP180 internalization via a macropinocytic pathway. Am. J. Pathol. 2013, 182, 828–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ando, H.; Yoshimoto, S.; Yoshida, M.; Shimoda, N.; Tadokoro, R.; Kohda, H.; Ishikawa, M.; Nishikata, T.; Katayama, B.; Ozawa, T.; et al. Dermal Fibroblasts Internalize Phosphatidylserine-Exposed Secretory Melanosome Clusters and Apoptotic Melanocytes. Int. J. Mol. Sci. 2020, 21, 5789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165789
Ando H, Yoshimoto S, Yoshida M, Shimoda N, Tadokoro R, Kohda H, Ishikawa M, Nishikata T, Katayama B, Ozawa T, et al. Dermal Fibroblasts Internalize Phosphatidylserine-Exposed Secretory Melanosome Clusters and Apoptotic Melanocytes. International Journal of Molecular Sciences. 2020; 21(16):5789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165789
Chicago/Turabian StyleAndo, Hideya, Satoshi Yoshimoto, Moemi Yoshida, Nene Shimoda, Ryosuke Tadokoro, Haruka Kohda, Mami Ishikawa, Takahito Nishikata, Bunpei Katayama, Toshiyuki Ozawa, and et al. 2020. "Dermal Fibroblasts Internalize Phosphatidylserine-Exposed Secretory Melanosome Clusters and Apoptotic Melanocytes" International Journal of Molecular Sciences 21, no. 16: 5789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165789