Transcriptional Induction of Cystathionine γ-Lyase, a Reactive Sulfur-Producing Enzyme, by Copper Diethyldithiocarbamate in Cultured Vascular Endothelial Cells

 , , and
, , and
Abstract
:1. Introduction
2. Results
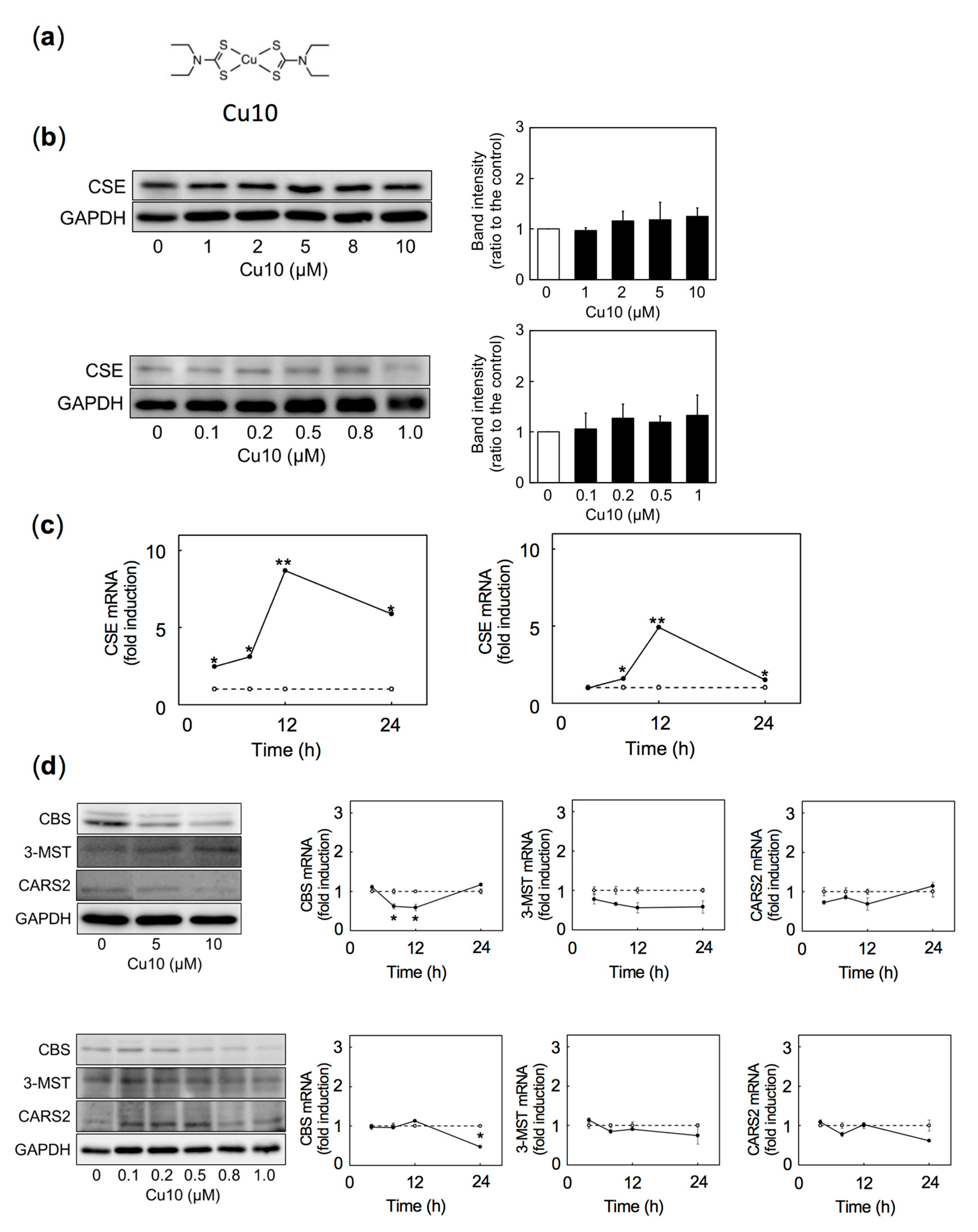
2.1. Transcriptional Induction of CSE, a Reactive Sulfur Species-Producing Enzyme
2.2. Role of Copper and Ligand Structure of the Cu10 Molecule in Induction of Endothelial CSE Transcription
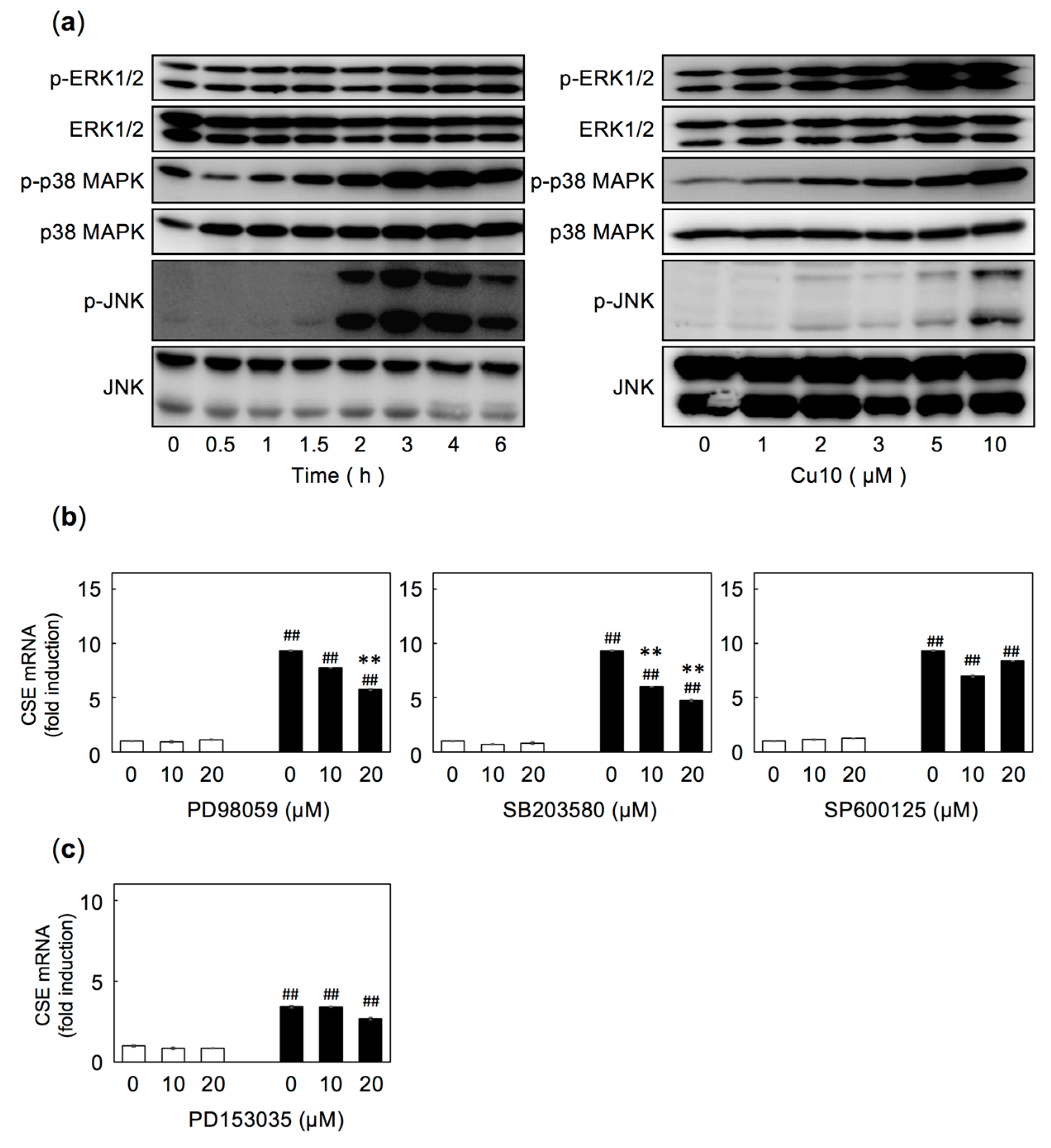
2.3. The extracellular signal-regulated kinase (ERK) and p38 MAPK Pathways are Involved in Cu10-Mediated Induction of CSE Expression in Endothelial Cells
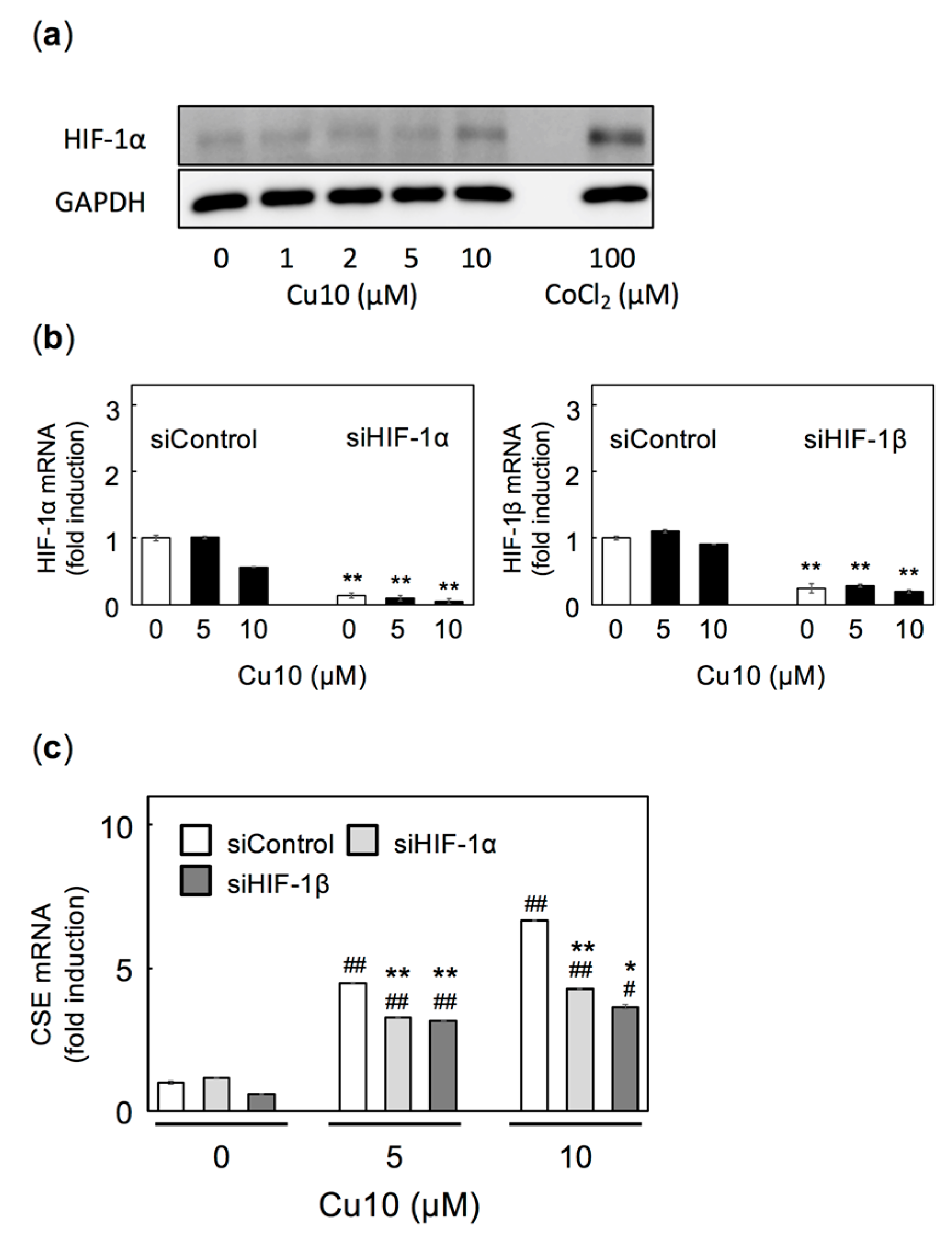
2.4. HIF-1α/HIF-1β is Also Involved in Cu10-Mediated Induction of Endothelial Cell CSE Transcription
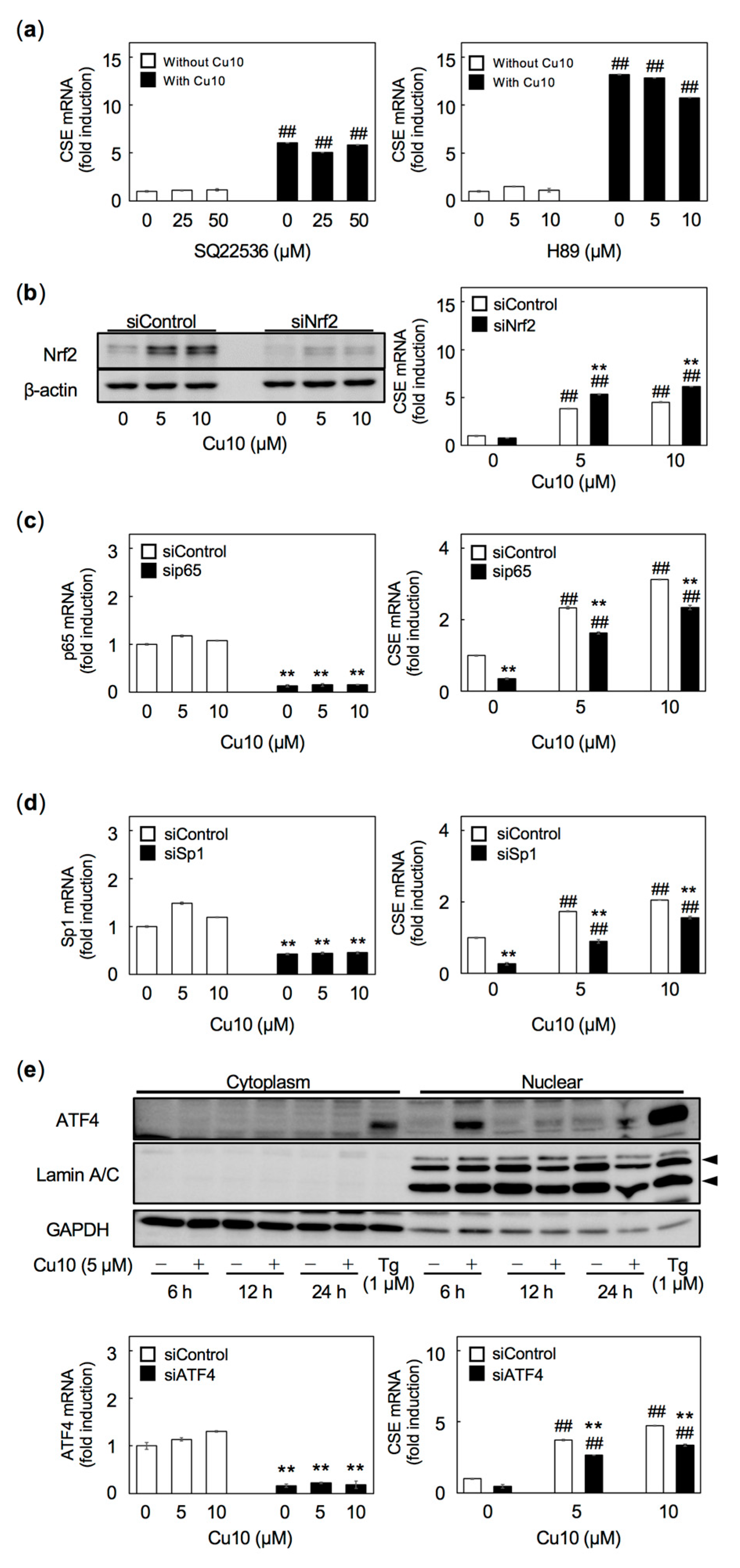
3. Discussions
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Treatments
4.3. siRNA Transfection
4.4. Western Blot Analysis
4.5. Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ATF4 | Activating transcription factor 4 |
| B2M | β2-microgloblin |
| CARS2 | Cysteinyl-tRNA synthetase 2 |
| CBS | Cystathionine beta-synthase |
| CSE | Cystathionine gamma-lyase |
| Cu10 | Copper (II) bis(diethyldithiocarbamate) |
| Cu36 | Bis(1-pyrrolidinecarbodithioato)copper (II) |
| Cu38 | Bis(1-piperidinecarbodithioato)copper (II) |
| Cu40 | Bis(4-morpholinecarbodithioato)copper (II) |
| Cu42 | Bis(4-methylpiperazine-1-carbodithioato)copper (II) |
| Cu50 | Bis(diisobutyldithiocarbamato)copper (II) |
| Cu52 | Bis(butylethyldithiocarbamato)copper (II) |
| EGFR | Epidermal growth factor receptor |
| ERK | Extracellular signal-regulated kinase |
| Fe05 | Iron(III) tris(diethyldithiocarbamate) |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| HIF-1 | Hypoxia-inducible factor-1 |
| JNK | c-Jun N-terminal kinase |
| MAPK | Mitrogen-activated phosphate kinase |
| 3-MST | 3-mercaptopyruvate sulfertransferase |
| Na01 | Sodium diethyldithiocarbamate trihydrate |
| Ni06 | Nickel(II) bis(diethyldithiocarbamate) |
| PKA | Protein kinase A |
| NF-κB | Nuclear factor-kappa B |
| Nrf2 | NF-E2-related factor 2 |
| RSS | Reactive sulfur species |
| Sp1 | Specificity protein-1 |
| TTBS | Tween tris buffered saline |
| Zn01 | Zinc(II) bis(diethyldithiocarbamate) |
References
- Kaji, T.; Mishima, A.; Yamamoto, C.; Sakamoto, M.; Koizumi, F. Effect of cadmium on the monolayer maintenance of vascular endothelial cells in culture. Toxicology 1992, 71, 267–276. [Google Scholar] [CrossRef]
- Yamamoto, C.; Kaji, T.; Sakamoto, M.; Kozuka, H. Cadmium stimulation of plasminogen activator inhibitor-1 release from human vascular endothelial cells in culture. Toxicology 1993, 83, 215–223. [Google Scholar] [CrossRef]
- Kaji, T.; Yamamoto, C.; Sakamoto, M.; Kozuka, H. Inhibitory effect of lead on the release of tissue plasminogen activator from human vascular endothelial cells in culture. Toxicology 1992, 73, 219–227. [Google Scholar] [CrossRef]
- Kaji, T.; Mishima, A.; Yamamoto, C.; Sakamoto, M.; Kozuka, H. Zinc protection against cadmium-induced destruction of the monolayer of cultured vascular endothelial cells. Toxicol. Lett. 1993, 66, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Probst, G.S.; Bousquet, W.F.; Miya, T.S. Correlation of hepatic metallothionein concentrations with acute cadmium toxicity in the mouse. Toxicol. Appl. Pharmacol. 1977, 39, 61–69. [Google Scholar] [CrossRef]
- Kaji, T.; Mishima, A.; Koyanagi, E.; Yamamoto, C.; Sakamoto, M.; Kozuka, H. Possible mechanism for zinc protection against cadmium cytotoxicity in cultured vascular endothelial cells. Toxicology 1992, 76, 257–270. [Google Scholar] [CrossRef]
- Fujie, T.; Segawa, Y.; Uehara, A.; Nakamura, T.; Kimura, T.; Yoshida, E.; Yamamoto, C.; Uchiyama, M.; Naka, H.; Kaji, T. Zinc diethyldithiocarbamate as an inducer of metallothionein in cultured vascular endothelial cells. J. Toxicol. Sci. 2016, 41, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, E.; Toyama, T.; Shinkai, Y.; Sawa, T.; Akaike, T.; Kumagai, Y. Detoxification of methylmercury by hydrogen sulfide-producing enzyme in mammalian cells. Chem. Res. Toxicol. 2011, 24, 1633–1635. [Google Scholar] [CrossRef]
- Shinkai, Y.; Masuda, A.; Akiyama, M.; Xian, M.; Kumagai, Y. Cadmium-mediated activation of the HSP90/HSF1 pathway regulated by reactive persulfides/polysulfides. Toxicol. Sci. 2017, 156, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [Green Version]
- Módis, K.; Asimakopoulou, A.; Coletta, C.; Papapetropoulos, A.; Szabo, C. Oxidative stress suppresses the cellular bioenergetic effect of the 3-mercaptopyruvate sulfurtransferase/hydrogen sulfide pathway. Biochem. Biophys. Res. Commun. 2013, 433, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Akaike, T.; Ida, T.; Wei, F.Y.; Nishida, M.; Kumagai, Y.; Alam, M.M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S.; et al. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujie, T.; Hara, T.; Kaji, T. Toxicology of organic–inorganic hybrid molecules: Bio-organometallics and its toxicology. J. Toxicol. Sci. 2016, 41, SP81–SP88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohri, K.; Yoshida, E.; Yasuike, S.; Fujie, T.; Yamamoto, C.; Kaji, T. The cytotoxicity of organobismuth compounds with certain molecular structures can be diminished by replacing the bismuth atom with an antimony atom in the molecules. J. Toxicol. Sci. 2015, 40, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Yoshida, E.; Fujie, T.; Ogata, F.; Yamamoto, C.; Kawasaki, N.; Kaji, T. Synergistic cytotoxicity caused by forming a complex of copper and 2,9-dimethyl-1,10-phenanthroline in cultured vascular endothelial cells. J. Toxicol. Sci. 2017, 42, 683–687. [Google Scholar] [CrossRef] [Green Version]
- Hara, T.; Nonaka, Y.; Yasuike, S.; Kaji, T.; Yamamoto, C. Structure-activity relationship of [1,5]azastibocines in cytotoxicity to vascular endothelial cells. J. Toxicol. Sci. 2018, 43, 735–740. [Google Scholar] [CrossRef]
- Fujie, T.; Yamamoto, T.; Yamamoto, C.; Kaji, T. Bis (1,4-dihydro-2-methyl-1-phenyl-4-thioxo-3-pyridiolato)zinc(II) exhibits strong cytotoxicity and a high intracellular accumulation in cultured vascular endothelial cells. J. Toxicol. Sci. 2019, 44, 113–120. [Google Scholar] [CrossRef]
- Hara, T.; Nakano, S.; Kitamura, Y.; Yamamoto, C.; Yasuike, S.; Kaji, T. Intracellular accumulation-independent cytotoxicity of pentavalent organoantimony compounds in cultured vascular endothelial cells. J. Toxicol. Sci. 2019, 44, 845–848. [Google Scholar] [CrossRef] [Green Version]
- Fujie, T.; Murakami, M.; Yoshida, E.; Tachinami, T.; Shinkai, Y.; Fujiwara, Y.; Yamamoto, C.; Kumagai, Y.; Naka, H.; Kaji, T. Copper diethyldithiocarbamate as an activator of Nrf2 in cultured vascular endothelial cells. J. Biol. Inorg. Chem. 2016, 21, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Fujie, T.; Segawa, Y.; Yoshida, E.; Kimura, T.; Fujiwara, Y.; Yamamoto, C.; Satoh, M.; Naka, H.; Kaji, T. Induction of metallothionein isoforms by copper diethyldithiocarbamate in cultured vascular endothelial cells. J. Toxicol. Sci. 2016, 41, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Fujie, T.; Murakami, M.; Yoshida, E.; Yasuike, S.; Kimura, T.; Fujiwara, Y.; Yamamoto, C.; Kaji, Y. Transcriptional induction of metallothionein by tris(pentafluorophenyl)stibane in cultured bovine aortic endothelial cells. Int. J. Mol. Sci. 2016, 17, 1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujie, T.; Takenaka, F.; Yoshida, E.; Yasuike, S.; Fujiwara, Y.; Shinkai, Y.; Kumagai, Y.; Yamamoto, C.; Kaji, T. Possible mechanisms underlying transcriptional induction of metallothionein isoforms by tris (pentafluorophenyl) stibane, tris (pentafluorophenyl) arsane, and tris (pentafluorophenyl) phosphane in cultured bovine aortic endothelial cells. J. Toxicol. Sci. 2019, 44, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef] [PubMed]
- De Agostini, A.I.; Watkins, S.C.; Slayter, H.S.; Youssoufian, H.; Rosenberg, R.D. Localization of anticoagulantly active heparan sulfate proteoglycans in vascular endothelium: Antithrombin binding on cultured endothelial cells and perfused rat aorta. J. Cell Biol. 1990, 111, 1293–1304. [Google Scholar] [CrossRef]
- Hara, T.; Kojima, T.; Matsuzaki, H.; Nakamura, T.; Yoshida, E.; Fujiwara, Y.; Yamamoto, C.; Saito, S.; Kaji, T. Induction of syndecan-4 by organic-inorganic hybrid molecules with a 1,10-phenanthroline structure in cultured vascular endothelial cells. Int. J. Mol. Sci. 2017, 18, 352. [Google Scholar] [CrossRef] [Green Version]
- Hara, T.; Tatsuishi, H.; Banno, T.; Fujie, T.; Yamamoto, C.; Naka, H.; Kaji, T. Copper(II) bis(diethyldithiocarbamate) induces the expression of syndecan-4, a transmembrane heparan sulfate proteoglycan, via p38 MAPK activation in vascular endothelial cells. Int. J. Mol. Sci. 2018, 19, 3302. [Google Scholar] [CrossRef] [Green Version]
- Bakthavatsalam, S.; Sleeper, M.L.; Dharani, A.; George, D.J.; Zhang, T.; Franz, K.J. Leveraging γ-glutamyl transferase to direct cytotoxicity of copper dithiocarbamates against prostate cancer cells. Angew. Chem. Int. Ed. 2018, 57, 12780–12784. [Google Scholar] [CrossRef]
- Bakthavatsalam, S.; Wiangnak, P.; George, D.J.; Zhang, T.; Franz, K.J. Dithiocarbamate prodrugs activated by prostate specific antigen to target prostate cancer. Bioorg. Med. Chem. Lett. 2020, 30, 127148. [Google Scholar] [CrossRef]
- Mistry, R.K.; Murray, T.V.; Prysyazhna, O.; Martin, D.; Burgoyne, J.R.; Santos, C.; Eaton, P.; Shah, A.M.; Brewer, A.C. Transcriptional regulation of cystathionine-γ-lyase in endothelial cells by NADPH oxidase 4-dependent signaling. J. Biol. Chem. 2016, 291, 1774–1788. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, Z.; Shi, S.; Gao, F.; Wu, J.; Dong, S.; Zhang, W.; Liu, Y.; Zhong, X. Calcium sensing receptor initiating cystathionine-gamma-lyase/hydrogen sulfide pathway to inhibit platelet activation in hyperhomocysteinemia rat. Exp. Cell Res. 2017, 358, 171–181. [Google Scholar] [CrossRef]
- Trocha, K.M.; Kip, P.; Tao, M.; MacArthur, M.R.; Treviño-Villarreal, H.; Longchamp, A.; Toussaint, W.; Lambrecht, B.N.; De Vries, M.R.; Quax, P.H.A.; et al. Short-term preoperative protein restriction attenuates vein graft disease via induction of cystathionine γ-lyase. Cardiovasc. Res. 2020, 116, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Kaji, T.; Yamada, A.; Miyajima, S.; Yamamoto, C.; Fujiwara, Y.; Wight, T.N.; Kinsella, M.G. Cell density-dependent regulation of proteoglycan synthesis by transforming growth factor-β1 in cultured bovine aortic endothelial cells. J. Biol. Chem. 2000, 275, 1463–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, C.; Morita, Y.; Yamaguchi, S.; Hayashi, T.; Kaji, T. Colominic acid inhibits the proliferation of cultured bovine aortic endothelial cells and injures their monolayers: Cell density-dependent effects prevented by sulfation. Life Sci. 2006, 78, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Hirooka, T.; Fujiwara, Y.; Shinkai, Y.; Yamamoto, C.; Yasutake, A.; Satoh, M.; Eto, K.; Kaji, T. Resistance of human brain microvascular endothelial cells in culture to methylmercury: Cell-density-dependent defense mechanisms. J. Toxicol. Sci. 2010, 35, 278–294. [Google Scholar] [CrossRef] [Green Version]
- Hara, T.; Sakamaki, S.; Ikeda, A.; Nakamura, T.; Yamamoto, C.; Kaji, T. Cell density-dependent modulation of perlecan synthesis by dichloro (2,9-dimethyl-1,10-phenanthroline)zinc(II) in vascular endothelial cells. J. Toxicol. Sci. 2020, 45, 109–115. [Google Scholar] [CrossRef]
- Hara, T.; Yabushita, S.; Yamamoto, C.; Kaji, T. Cell density-dependent fibroblast growth factor-2 signaling regulates syndecan-4 expression in cultured vascular endothelial cells. Int. J. Mol. Sci. 2020, 21, 3698. [Google Scholar] [CrossRef]
- Yoshida, E.; Kurita, M.; Eto, K.; Kumagai, Y.; Kaji, T. Methylmercury promotes prostacyclin release from cultured human brain microvascular endothelial cells via induction of cyclooxygenase-2 through activation of the EGFR-p38 MAPK pathway by inhibiting protein tyrosine phosphatase 1B activity. Toxicology 2017, 392, 40–46. [Google Scholar] [CrossRef]
- Wang, M.; Guo, Z.; Wang, S. Regulation of cystathionine γ-lyase in mammalian cells by hypoxia. Biochem. Genet. 2014, 52, 29–37. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, Q.; Nishitani, J.; Brown, J.; Shi, S.; Le, A.D. Overexpression of human papillomavirus type 16 oncoproteins enhances hypoxia-Inducible 1actor 1α protein accumulation and vascular endothelial growth factor expression in human cervical carcinoma cells. Clin. Cancer Res. 2007, 13, 2568–2576. [Google Scholar] [CrossRef] [Green Version]
- Renga, B.; Bucci, M.; Cipriani, S.; Carino, A.; Monti, M.C.; Zampella, A.; Gargiulo, A.; D’Emmanuele di Villa Bianca, R.; Distrutti, E.; Fiorucci, S. Cystathionine γ-lyase, a H2S-generating enzyme, is a GPBAR1-regulated gene and contributes to vasodilation caused by secondary bile acids. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H114–H126. [Google Scholar] [CrossRef] [Green Version]
- Hosoki, R.; Matsuki, N.; Kimura, H. The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badiei, A.; Gieseg, S.; Davies, S.; Izani Othman, M.; Bhatia, M. LPS up-regulates cystathionine γ -lyase gene expression in primary human macrophages via NF-κB/ERK pathway. Inflamm. Allergy Drug Targets 2015, 14, 99–104. [Google Scholar] [CrossRef]
- Gong, H.; Chen, Z.; Zhang, X.; Li, Y.; Zhang, J.; Chen, Y.; Ding, Y.; Zhang, G.; Yang, C.; Zhu, Y.; et al. Urotensin II protects cardiomyocytes from apoptosis induced by oxidative stress through the CSE/H2S pathway. Int. J. Mol. Sci. 2015, 16, 12482–12498. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, S.; Kimura, T.; Umeki, T.; Kimura, Y.; Kimura, H.; Ishii, I.; Itoh, N.; Naito, Y.; Yamamoto, H.; Niki, I. Protein phosphorylation involved in the gene expression of the hydrogen sulphide producing enzyme cystathionine γ-lyase in the pancreatic β-cell. Mol. Cell. Endocrinol. 2012, 350, 31–38. [Google Scholar] [CrossRef]
- Zheng, Y.; Luo, N.; Mu, D.; Jiang, P.; Liu, R.; Sun, H.; Xiong, S.; Liu, X.; Wang, L.; Chu, Y. Lipopolysaccharide regulates biosynthesis of cystathionine γ-lyase and hydrogen sulfide through Toll-like receptor-4/p38 and Toll-like receptor-4/NF-κB pathways in macrophages. In Vitro Cell. Dev. Biol. Anim. 2013, 49, 679–688. [Google Scholar] [CrossRef]
- Kandil, S.; Brennan, L.; McBean, G.J. Glutathione depletion causes a JNK and p38MAPK-mediated increase in expression of cystathionine-gamma-lyase and upregulation of the transsulfuration pathway in C6 glioma cells. Neurochem. Int. 2010, 4, 611–619. [Google Scholar] [CrossRef]
- Calvert, J.W.; Jha, S.; Gundewar, S.; Elrod, J.W.; Ramachandran, A.; Pattillo, C.B.; Kevil, C.G.; Lefer, D.J. Hydrogen sulfide mediates cardioprotection through Nrf2 signaling. Circ. Res. 2009, 105, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Fiorucci, S.; Antonelli, E.; Distrutti, E.; Rizzo, G.; Mencarelli, A.; Orlandi, S.; Zanardo, R.; Renga, B.; Di Sante, M.; Morelli, A.; et al. Inhibition of hydrogen sulfide generation contributes to gastric injury caused by anti-inflammatory nonsteroidal drugs. Gastroenterology 2005, 4, 1210–1224. [Google Scholar] [CrossRef] [Green Version]
- Yin, P.; Zhao, C.; Li, Z.; Mei, C.; Yao, W.; Liu, Y.; Li, N.; Qi, J.; Wang, L.; Shi, Y.; et al. Sp1 is involved in regulation of cystathionine gamma-lyase gene expression and biological function by PI3K/Akt pathway in human hepatocellular carcinoma cell lines. Cell Signal. 2012, 24, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, G.; Tang, G.; Wu, L.; Wang, R. Rat pancreatic level of cystathionine γ-lyase is regulated by glucose level via specificity protein 1 (SP1) phosphorylation. Diabetologia 2011, 10, 2615–2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racine, M.; Fu, M.; Shuang, T.; Zhang, Y.; Pei, Y.; Wu, L.; Wang, R.; Yang, G. Reversal of Sp1 transactivation and TGFβ1/SMAD1 signaling by H2S prevent nickel-induced fibroblast activation. Toxicol. Appl. Pharmacol. 2018, 356, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ali, A.; Jin, Z.; Pei, Y.; Yang, G. Induction of cystathionine gamma-lyase expression and metallothionein-1 S-sulfhydration alleviate cadmium-induced cell death in myoblast cells. Ecotoxicol. Environ. Saf. 2019, 179, 222–231. [Google Scholar] [CrossRef]
- Cheung, S.H.; Kwok, W.K.; To, K.F.; Lau, J.Y. Anti-atherogenic effect of hydrogen sulfide by over-expression of cystathionine gamma-lyase (CSE) gene. PLoS ONE 2014, 11, e113038. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Guo, F.; Li, L.; Guo, L.; Hou, Y.; Hao, E.; Yan, S.; Allen, T.D.; Liu, J. Cadmium induces vascular permeability via activation of the p38 MAPK pathway. Biochem. Biophys. Res. Commun. 2014, 450, 447–452. [Google Scholar] [CrossRef]
- Van Hove, J.L.K.; Freehauf, C.L.; Ficicioglu, C.; Pena, L.D.M.; Moreau, K.L.; Henthorn, T.K.; Christians, U.; Jiang, H.; Cowan, T.M.; Young, S.P.; et al. Biomarkers of oxidative stress, inflammation, and vascular dysfunction in inherited cystathionine β-synthase deficient homocystinuria and the impact of taurine treatment in a phase 1/2 human clinical trial. J. Inherit. Metab. Dis. 2019, 42, 424–437. [Google Scholar] [CrossRef]
- Barroso, M.; Kao, D.; Blom, H.J.; Tavares de Almeida, I.; Castro, R.; Loscalzo, J.; Handy, D.E. S-adenosylhomocysteine induces inflammation through NFkB: A possible role for EZH2 in endothelial cell activation. Biochim. Biophys. Acta 2016, 1862, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Mosharov, E.; Cranford, M.R.; Banerjee, R. The quantitatively important relationship between homocysteine metabolism and glutathione synthesis by the transsulfuration pathway and its regulation by redox changes. Biochemistry 2000, 39, 13005–13011. [Google Scholar] [CrossRef]
- Carta, F.; Aggarwal, M.; Maresca, A.; Scozzafava, A.; McKenna, R.; Masini, E.; Supuran, C.T. Dithiocarbamates strongly inhibit carbonic anhydrase and show antiglaucoma action in vivo. J. Med. Chem. 2012, 55, 1721–1730. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense (5′→3′) | Antisense (5′→3′) |
|---|---|---|
| HIF-1α | GGGAUUAACUCAGUUUGAACUdTdT | UUCAAACUGAGUUAAUCCCAUdTdT |
| HIF-1β | GAACUCUUAGGAAAGAAUAUUdTdT | UAUUCUUUCCUAAGAGUUCCUdTdT |
| Nrf2 | CCAUUGAUCUCUCUGAUCUdTdT | AGAUCAGAGAGAUCAAUCGdTdT |
| p65 | AUUGAAAGGGCUCUUUUUCAUdTdT | GAAAAAGAGCCCUUUCAAUGGdTdT |
| Sp1 | GUUUAUAUAUACAUACAUAAU | UAUGUAUGUAUAUAUAAACUA |
| ATF4 | AAUCAAACUCCUUCAAAUCdTdT | GAUUUGAAGGAGUUUGAUUdTdT |
| Negative control | UUCUCCGAACGUGUCACGUdTdT | ACGUGACACGUUCGGAGAAdTdT |
| Gene | Sense (5′→3′) | Antisense (5′→3′) |
|---|---|---|
| CSE | TCTCTTGGAGCAGTTCCATCTCCTA | GCAGCCCAGGATAAATAACCTTTTC |
| CBS | GGACTCGGTGCGGAACTACA | GGCAACACGGTCAGCGG |
| 3-MST | GCAGTGGGTGGCTGAGGC | CGATGTCAAAGAAGGCGGC |
| CARS2 | GAGGCGACAGGTACGGCAAG | CAGACTGGCGATGGTGGAAC |
| HIF-1α | GCTTGCTCATCAGTTGCCAC | GCATCCAGAAGTTTCCTCACAC |
| HIF-1β | TAAGGAGCGGTTTGCCAGGTC | TTCTGTTATGTAGGCTGTCATCTTGTTC |
| p65 | GATGGCTTCTATGAGGCTGAG | TTGTTGTTGGTCTGGATGC |
| Sp1 | CTCTAAGCATCAGGAATCAGAAGTC | TCAGAAGCCCACACATCAAAG |
| ATF4 | TGGTCTCAGACAACAGCAAG | AGCTCATCTGGCATGGTTTC |
| B2M | CCATCCAGCGTCCTCCAAAGA | TTCAATCTGGGGTGGATGGAA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujie, T.; Takahashi, A.; Takahashi, M.; Hara, T.; Soyama, A.; Makino, K.; Takahashi, H.; Yamamoto, C.; Kumagai, Y.; Naka, H.; et al. Transcriptional Induction of Cystathionine γ-Lyase, a Reactive Sulfur-Producing Enzyme, by Copper Diethyldithiocarbamate in Cultured Vascular Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 6053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176053
Fujie T, Takahashi A, Takahashi M, Hara T, Soyama A, Makino K, Takahashi H, Yamamoto C, Kumagai Y, Naka H, et al. Transcriptional Induction of Cystathionine γ-Lyase, a Reactive Sulfur-Producing Enzyme, by Copper Diethyldithiocarbamate in Cultured Vascular Endothelial Cells. International Journal of Molecular Sciences. 2020; 21(17):6053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176053
Chicago/Turabian StyleFujie, Tomoya, Akane Takahashi, Musubu Takahashi, Takato Hara, Asuka Soyama, Kosho Makino, Hideyo Takahashi, Chika Yamamoto, Yoshito Kumagai, Hiroshi Naka, and et al. 2020. "Transcriptional Induction of Cystathionine γ-Lyase, a Reactive Sulfur-Producing Enzyme, by Copper Diethyldithiocarbamate in Cultured Vascular Endothelial Cells" International Journal of Molecular Sciences 21, no. 17: 6053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176053