Discriminating miRNA Profiles between Endometrioid Well- and Poorly-Differentiated Tumours and Endometrioid and Serous Subtypes of Endometrial Cancers


Abstract
:1. Introduction
2. Results
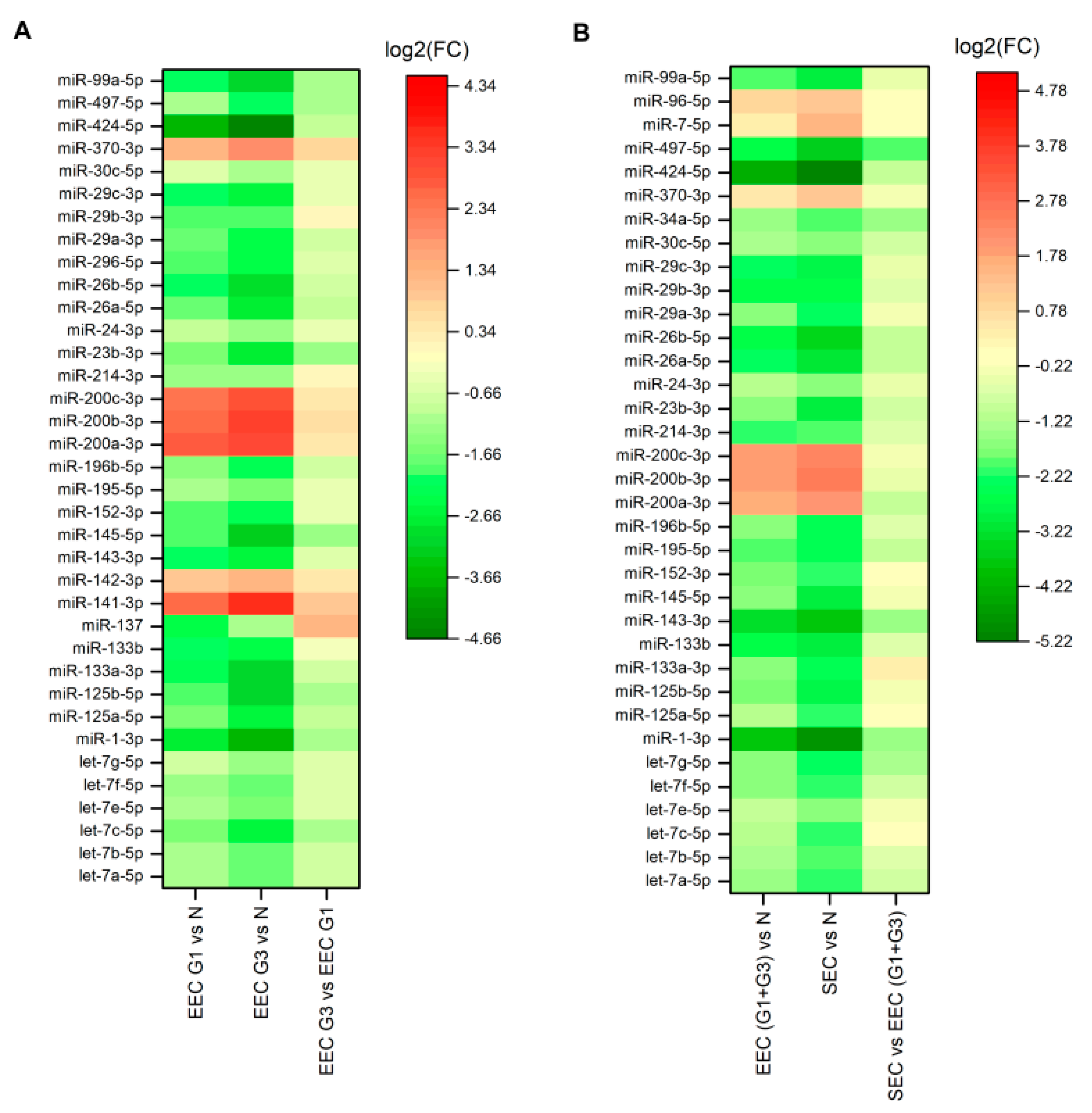
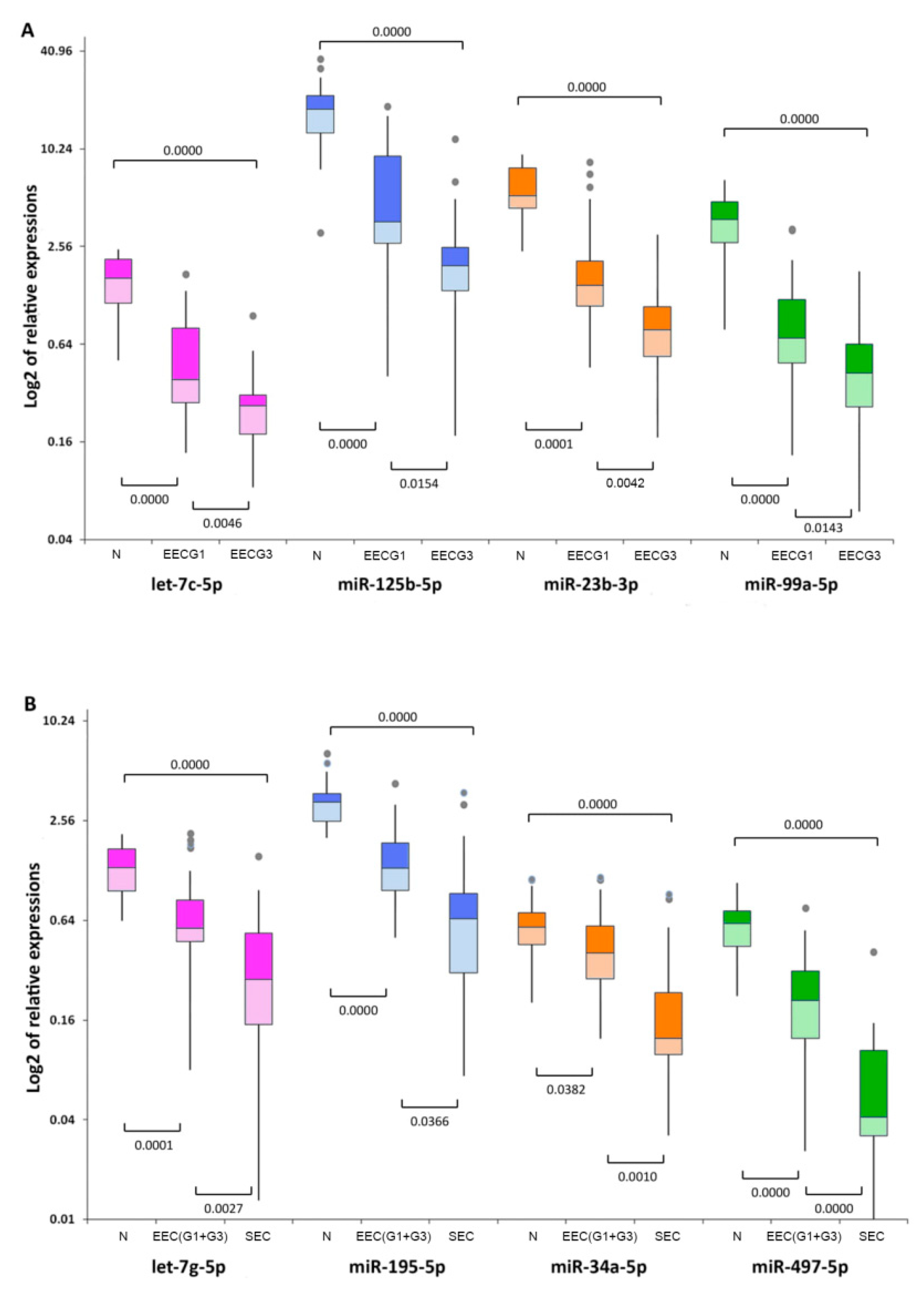
2.1. Differences in miRNA Expression between Well- and Poorly-Differentiated EEC
2.2. Differences in miRNA Expression between Endometrioid and Serous EC
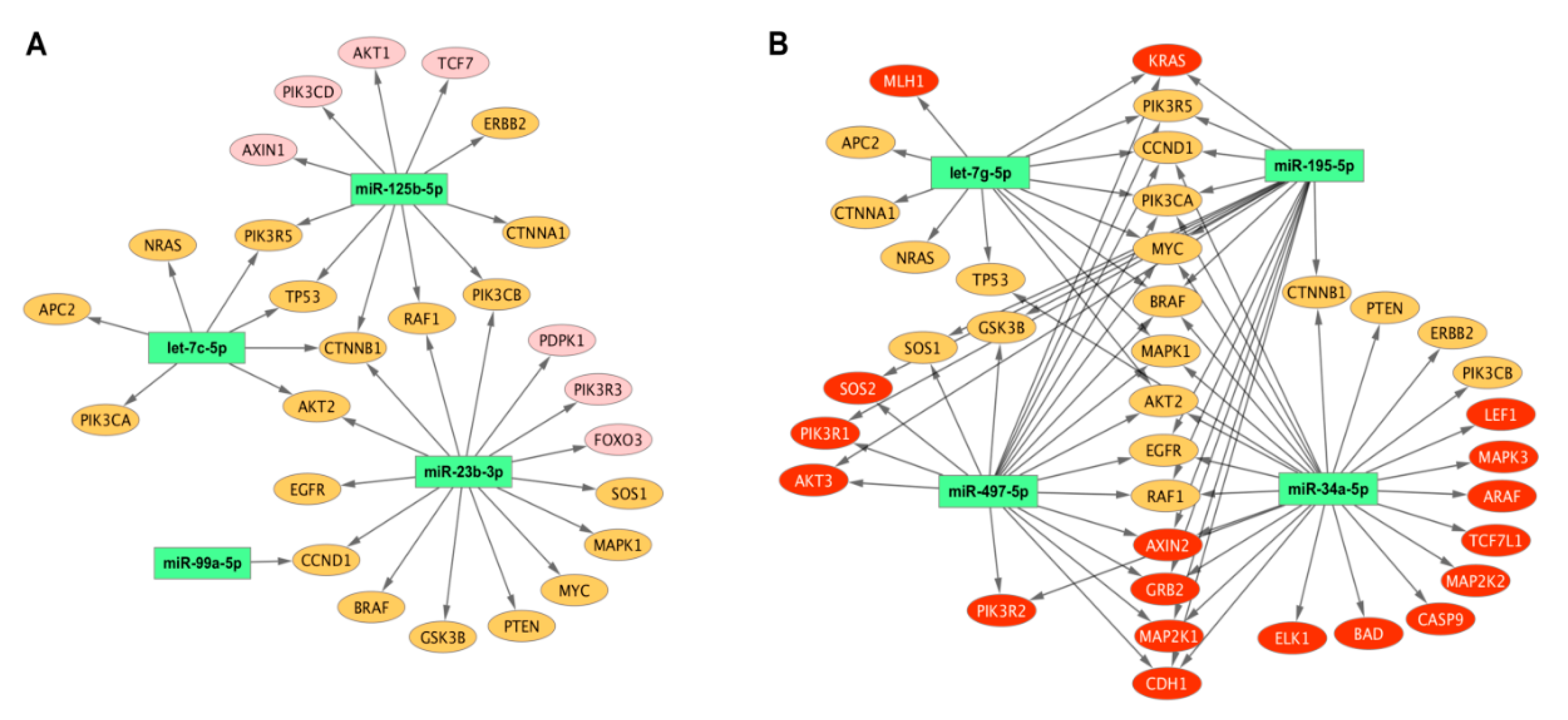
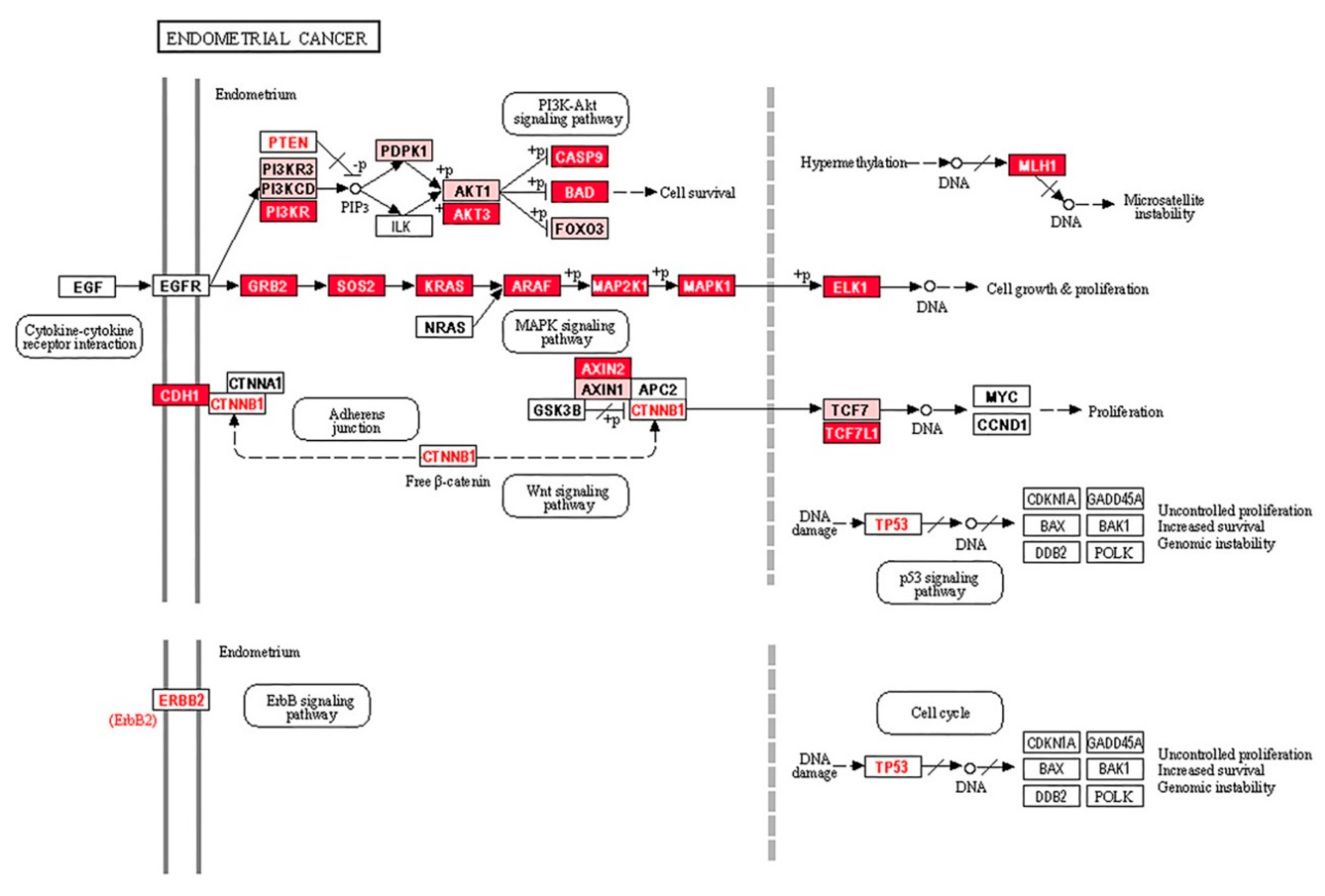
2.3. Aberrant miRNA Expression in EC Tumourigenesis
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. miRNA Extraction and Real-Time PCR
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EC | Endometrial Carcinoma |
| EEC | Endometrioid Endometrial Carcinoma |
| SEC | Serous Endometrial Carcinoma |
| TCGA | The Cancer Genome Atlas |
| MSI | Microsatellite Instability |
| miRNA | MicroRNA |
| G3 | Grade 3, Poorly-differentiated tumour |
| G1 | Grade 1, Well-differentiated tumour |
| FFPE | Formalin-Fixed Paraffin Embedded |
| EMT | Epithelial-Mesenchymal Transition |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Murali, R.; Soslow, R.A.; Weigelt, B. Classification of endometrial carcinoma: More than two types. Lancet Oncol. 2014, 7, e268–e278. [Google Scholar] [CrossRef]
- Bokhman, J.V. Two pathogenetic types of endometrial carcinoma. Gynecol. Oncol. 1983, 15, 10–17. [Google Scholar] [CrossRef]
- Matias-Guiu, X.; Prat, J. Molecular pathology of endometrial carcinoma. Histopathology 2013, 62, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Zaino, R.; Carinelli, S.G.; Ellenson, L.H.; End, C.; Katabuchi, H.; Konishi, I.; Lax, S.; Matias-Guiu, X.; Mutter, G.L.; Peters, W.A., III; et al. Epithelial tumours and precursors. In WHO Classification of Tumours of Female Reproductive Organs, 4th ed.; Kurman, R., Carcangiu, M.L., Herrington, S., Young, R.H., Eds.; IARC: Lyon, France, 2014; Volume 6, pp. 125–134. [Google Scholar]
- Colombo, N.; Preti, E.; Landoni, F.; Carinelli, S.; Colombo, A.; Marini, C.; Sessa, C. ESMO Guidelines Working Group Endometrial Cancer. ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-up. Ann. Anncol. 2013, 24, vi33–vi38. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network; Kandoth, C.; Schultz, N.; Cherniack, A.D.; Akbani, R.; Liu, Y.; Shen, H.; Robertson, A.G.; Pashtan, I.; Shen, R.; et al. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar]
- Morice, P.; Leary, A.; Creutzberg, C.; Abu-Rustum, N.; Darai, E. Endometrial cancer. Lancet 2016, 387, 1094–1108. [Google Scholar] [CrossRef]
- Bell, D.W. Novel genetic targets in endometrial cancer. Expert Opin. Ther. Targets 2014, 18, 725–730. [Google Scholar] [CrossRef]
- Esteller, M.; Catasus, L.; Matias-Guiu, X.; Mutter, G.L.; Prat, J.; Baylin, S.B.; Herman, J.G. hMLH1 promoter hypermethylation is an early event in human endometrial tumorigenesis. Am. J. Pathol. 1999, 155, 1767–1772. [Google Scholar] [CrossRef] [Green Version]
- Salvesen, H.B.; MacDonald, N.; Ryan, A.; Jacobs, I.J.; Lynch, E.D.; Akslen, L.A.; Das, S. PTEN methylation is associated with advanced stage and microsatellite instability in endometrial carcinoma. Int. J. Cancer 2001, 91, 22–26. [Google Scholar] [CrossRef]
- Liao, X.; Siu, M.K.; Chan, K.Y.; Wong, E.S.; Ngan, H.Y.; Chan, Q.K.; Li, A.S.; Khoo, U.S.; Cheung, A.N. Hypermethylation of RAS effector related genes and DNA methyltransferase 1 expression in endometrial carcinogenesis. Int. J. Cancer 2008, 123, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Bueno, G.; Hardisson, D.; Sanchez, C.; Sarrio, D.; Cassia, R.; Garcia-Rostan, G.; Prat, J.; Guo, M.; Herman, J.G.; Matias-Guiu, X.; et al. Abnormalities of the APC/beta-catenin pathway in endometrial cancer. Oncogene 2002, 21, 7981–7990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Dowdy, S.C.; Xue, A.; Shujuan, J.; Eberhardt, N.L.; Podratz, K.C.; Jiang, S.W. Opposite alterations of DNA methyltransferase gene expression in endometrioid and serous endometrial cancers. Gynecol. Oncol. 2005, 96, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Fambrini, M.; Bussani, C.; Sorbi, F.; Pieralli, A.; Cioni, R. Methylation of the HOXA10 homeobox gene promoter is associated with endometrial cancer: A pilot study. J. Obstet. Gynaecol. 2013, 33, 519–520. [Google Scholar] [CrossRef] [PubMed]
- Multinu, F.; Chen, J.; Madison, J.D.; Torres, M.; Casarin, J.; Visscher, D.; Shridhar, V.; Bakkum-Gamez, J.; Sherman, M.; Wentzensen, N.; et al. Analysis of DNA methylation in endometrial biopsies to predict risk of endometrial cancer. Gynecol. Oncol. 2020, 156, 682–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Xing, X.; Li, J.; Lowdon, R.F.; Zhou, Y.; Lin, N.; Zhang, B.; Sundaram, V.; Chiappinelli, K.B.; Hagemann, I.S.; et al. Comparative DNA methylome analysis of endometrial carcinoma reveals complex and distinct deregulation of cancer promoters and enhancers. BMC Genom. 2014, 15, 868. [Google Scholar] [CrossRef] [Green Version]
- van Schooneveld, E.; Wildiers, H.; Vergote, I.; Vermeulen, P.B.; Dirix, L.Y.; Van Laere, S.J. Dysregulation of microRNAs in breast cancer and their potential role as prognostic and predictive biomarkers in patient management. Breast Cancer Res. 2015, 18, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Xie, B.; Ding, Q.; Han, H.; Wu, D. miRCancer: A microRNA-cancer association database constructed by text mining on literature. Bioinformatics 2013, 29, 638–644. [Google Scholar] [CrossRef]
- Delangle, R.; De Foucher, T.; Larsen, A.K.; Sabbah, M.; Azaïs, H.; Bendifallah, S.; Daraï, E.; Ballester, M.; Mehats, C.; Uzan, C.; et al. The Use of microRNAs in the Management of Endometrial Cancer: A Meta-Analysis. Cancers 2019, 11, 832. [Google Scholar] [CrossRef] [Green Version]
- Cohn, D.E.; Fabbri, M.; Valeri, N.; Alder, H.; Ivanov, I.; Liu, C.G.; Croce, C.M.; Resnick, K.E. Comprehensive miRNA profiling of surgically staged endometrial cancer. Am. J. Obstet. Gynecol. 2010, 202, e1–e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroki, E.; Akahira, J.; Suzuki, F.; Nagase, S.; Ito, K.; Suzuki, T.; Sasano, H.; Yaegashi, N. Changes in microRNA expression levels correlate with clinicopathological features and prognoses in endometrial serous adenocarcinomas. Cancer Sci. 2010, 101, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Widodo; Djati, M.S.; Rifa’I, M. Role of MicroRNAs in carcinogenesis that potential for biomarker of endometrial cancer. Ann. Med. Surg. 2016, 7, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Gilks, C.B.; Oliva, E.; Soslow, R.A. Poor interobserver reproducibility in diagnosis of high-grade endometrial carcinoma. Am. J. Surg. Pathol. 2013, 37, 874–881. [Google Scholar] [CrossRef]
- Talhouk, A.; McAlpine, J.N. New classification of endometrial cancers: The development and potential applications of genomic-based classification in research and clinical care. Gynecol. Oncol. Res. Pract. 2016, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Molchadsky, A.; Rivlin, N.; Brosh, R.; Rotter, V.; Sarig, R. p53 is balancing development, differentiation and de-differentiation to assure cancer prevention. Carcinogenesis 2010, 31, 1501–1508. [Google Scholar] [CrossRef] [Green Version]
- Rivlin, N.; Koifman, G.; Rotter, V. p53 orchestrates between normal differentiation and cancer. Semin. Cancer Biol. 2015, 32, 10–17. [Google Scholar] [CrossRef]
- Kohler, M.F.; Berchuck, A.; Davidoff, A.M.; Humphrey, P.A.; Dodge, R.K.; Iglehart, J.D.; Soper, J.T.; Clarke-Pearson, D.L.; Bast, R.C., Jr.; Marks, J.R. Overexpression and mutation of p53 in endometrial carcinoma. Cancer Res. 1992, 52, 1622–1627. [Google Scholar]
- Köbel, M.; Ronnett, B.M.; Singh, N.; Soslow, R.A.; Gilks, C.B.; McCluggage, W.G. Interpretation of P53 Immunohistochemistry in Endometrial Carcinomas: Toward Increased Reproducibility. Int. J. Gynecol. Pathol. 2019, 38, S123–S131. [Google Scholar] [CrossRef]
- Le, M.T.; Teh, C.; Shyh-Chang, N.; Xie, H.; Zhou, B.; Korzh, V.; Lodish, H.F.; Lim, B. MicroRNA-125b is a novel negative regulator of p53. Genes Dev. 2009, 23, 862–876. [Google Scholar] [CrossRef] [Green Version]
- Navarro, F.; Lieberman, J. miR-34 and p53: New Insights into a Complex Functional Relationship. PLoS ONE 2015, 10, e0132767. [Google Scholar] [CrossRef] [PubMed]
- Boyerinas, B.; Park, S.M.; Hau, A.; Murmann, A.E.; Peter, M.E. The role of let-7 in cell differentiation and cancer. Endocr.-Relat. Cancer 2010, 17, F19–F36. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Li, Q.; Liu, S.; Wang, F.; Xiong, Z.; Chen, J.; Chen, H.; Yang, Y.; Tan, X.; Luo, Q.; et al. Integrated microRNA and mRNA transcriptome sequencing reveals the potential roles of miRNAs in stage I endometrioid endometrial carcinoma. PLoS ONE 2014, 9, e110163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, F.; Tang, Q.L.; Ma, F.; Cai, L.; Chen, M.; Ran, X.X.; Wang, X.Y.; Jiang, X.F. Analysis of the circular RNA transcriptome in the grade 3 endometrial cancer. Cancer Manag. Res. 2019, 11, 6215–6227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Chang, L.; Li, Z.; Gao, Q.; Cai, D.; Tian, Y.; Zeng, L.; Li, M. miR-99a and -99b inhibit cervical cancer cell proliferation and invasion by targeting mTOR signaling pathway. Med. Oncol. 2014, 31, 934. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhang, C.; Deng, F.; Zhang, Y. miR-99a suppresses the metastasis of human nonsmall cell lung cancer cells by targeting AKT1 signaling pathway. J. Cell. Biochem. 2015, 116, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.; Torres, K.; Pesci, A.; Ceccaroni, M.; Paszkowski, T.; Cassandrini, P.; Zamboni, G.; Maciejewski, R. Deregulation of miR-100, miR-99a and miR-199b in tissues and plasma coexists with increased expression of mTOR kinase in endometrioid endometrial carcinoma. BMC Cancer 2012, 12, 369. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, Z.; Zhang, X.; Lin, Y.; Luo, T.; Xiao, Z.; Zhou, Q. A dual PI3K/AKT/mTOR signaling inhibitor miR-99a suppresses endometrial carcinoma. Am. J. Transl. Res. 2016, 8, 719–731. [Google Scholar]
- Shang, C.; Lu, Y.M.; Meng, L.R. MicroRNA-125b down-regulation mediates endometrial cancer invasion by targeting ERBB2. Med. Sci. Monit. 2012, 18, BR149–BR155. [Google Scholar] [CrossRef]
- Halle, M.K.; Tangen, I.L.; Berg, H.F.; Hoivik, E.A.; Mauland, K.K.; Kusonmano, K.; Berg, A.; Hurtado, A.; Kalland, K.H.; Øyan, A.M.; et al. HER2 expression patterns in paired primary and metastatic endometrial cancer lesions. Br. J. Cancer 2018, 118, 378–387. [Google Scholar] [CrossRef] [Green Version]
- Murali, R.; Davidson, B.; Fadare, O.; Carlson, J.A.; Crum, C.P.; Gilks, C.B.; Irving, J.A.; Malpica, A.; Matias-Guiu, X.; McCluggage, W.G.; et al. High-grade Endometrial Carcinomas: Morphologic and Immunohistochemical Features, Diagnostic Challenges and Recommendations. Int. J. Gynecol. Pathol. 2019, 38, S40–S63. [Google Scholar] [CrossRef]
- Choi, Y.S.; Lee, K.E. The Significance of miR-34a Expression in Endometrial Carcinogenesis: Correlation with Expression of p16 and Ki-67 Proteins in Endometrial Cancers. J. Cancer Prev. 2015, 20, 268–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wang, W.; Huang, K.; Wang, Y.; Li, J.; Yang, X. MicroRNA-34a inhibits cells proliferation and invasion by downregulating Notch1 in endometrial cancer. Oncotarget 2017, 8, 111258–111270. [Google Scholar] [CrossRef] [PubMed]
- Siemens, H.; Jackstadt, R.; Hünten, S.; Kaller, M.; Menssen, A.; Götz, U.; Hermeking, H. miR-34 and SNAIL form a double-negative feedback loop to regulate epithelial-mesenchymal transitions. Cell Cycle 2011, 10, 4256–4271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirmer, U.; Doberstein, K.; Rupp, A.K.; Bretz, N.P.; Wuttig, D.; Kiefel, H.; Breunig, C.; Fiegl, H.; Müller-Holzner, E.; Zeillinger, R.; et al. Role of miR-34a as a suppressor of L1CAM in endometrial carcinoma. Oncotarget 2014, 5, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosse, T.; Nout, R.A.; Stelloo, E.; Dreef, E.; Nijman, H.W.; Jürgenliemk-Schulz, I.M.; Jobsen, J.J.; Creutzberg, C.L.; Smit, V.T. L1 cell adhesion molecule is a strong predictor for distant recurrence and overall survival in early stage endometrial cancer: Pooled PORTEC trial results. Eur. J. Cancer 2014, 50, 2602–2610. [Google Scholar] [CrossRef] [PubMed]
- Jurcevic, S.; Klinga-Levan, K.; Olsson, B.; Ejeskär, K. Verification of microRNA expression in human endometrial adenocarcinoma. BMC Cancer 2016, 16, 261. [Google Scholar] [CrossRef] [Green Version]
- Devor, E.J.; Hovey, A.M.; Goodheart, M.J.; Ramachandran, S.; Leslie, K.K. microRNA expression profiling of endometrial endometrioid adenocarcinomas and serous adenocarcinomas reveals profiles containing shared, unique and differentiating groups of microRNAs. Oncol. Rep. 2011, 26, 995–1002. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Ma, C.X.; Wang, J. Endometrial Carcinogenesis and Molecular Signaling Pathways. Am. J. Mol. Biol. 2014, 4, 134–149. [Google Scholar] [CrossRef] [Green Version]
- Dong, P.; Konno, Y.; Watari, H.; Hosaka, M.; Noguchi, M.; Sakuragi, N. The impact of microRNA-mediated PI3K/AKT signaling on epithelial-mesenchymal transition and cancer stemness in endometrial cancer. J. Transl. Med. 2014, 12, 231. [Google Scholar] [CrossRef]
- Hoshino, R.; Chatani, Y.; Yamori, T.; Tsuruo, T.; Oka, H.; Yoshida, O.; Shimada, Y.; Ari-i, S.; Wada, H.; Fujimoto, J.; et al. Constitutive Activation of the 41-/43-kDa Mitogen-Activated Protein Kinase Signaling Pathway in Human Tumors. Oncogene 1999, 18, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2[-Delta Delta C[T]] Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Sherman, B.T.; Huang, D.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. DAVID-WS: A stateful web service to facilitate gene/protein list analysis. Bioinformatics 2012, 28, 1805–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.; Li, J.; Huang, K.Y.; Shrestha, S.; Hong, H.C.; Tang, Y.; Chen, Y.G.; Jin, C.N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Pant, G.; Bhavnasi, Y.K.; Blanchard, S.G., Jr.; Brouwer, C. Pathview Web: User friendly pathway visualization and data integration. Nucleic Acids Res. 2017, 45, W501–W508. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC Subtypes | Differently Expressed miRNAs in Each of Two Evaluated Groups Compared to Controls |
|---|---|
| Endometrioid grade 1 vs. grade 3 | let-7a-5p, let-7b-5p, let-7c-5p, let-7e-5p, let-7f-5p, let-7g-5p, miR-1-3p, miR-125a-5p, miR-125b-5p, miR-133a-3p, miR-133b, miR-137, miR-141-3p, miR-142-3p, miR-143-3p, miR-145-5p, miR-152-3p, miR-195-5p, miR-196b-5p, miR-200a-3p, miR-200b-3p, miR-200c-3p, miR-214-3p, miR-23b-3p, miR-24-3p, miR-26a-5p, miR-26b-5p, miR-296-5p, miR-29a-3p, miR-29b-3p, miR-29c-3p, miR-30c-5p, miR-370-3p, miR-424-5p, miR-497-5p, miR-99a-5p |
| Endometrioid vs. Serous | let-7a-5p, let-7b-5p, let-7c-5p, let-7e-5p, let-7f-5p, let-7g-5p, miR-1-3p, miR-125a-5p, miR-125b-5p, miR-133a-3p, miR-133b, miR-143-3p, miR-145-5p, miR-152-3p, miR-195-5p, miR-196b-5p, miR-200a-3p, miR-200b-3p, miR-200c-3p, miR-214-3p, miR-23b-3p, miR-24-3p, miR-26a-5p, miR-26b-5p, miR-29a-3p, miR-29b-3p, miR-29c-3p, miR-30c-5p, miR-34a-5p, miR-370-3p, miR-424-5p, miR-497-5p, miR-7-5p, miR-96-5p, miR-99a-5p |
| EC Subtypes | Downregulated miRNAs * | Database | Enrichment p-value | Target Genes |
|---|---|---|---|---|
| Endometrioid grade 1 vs. grade 3 | let-7c-5p | TargetScan | 1.80 × 1020 | NRAS, PIK3R5, TP53, AKT2, CCND1, APC2, PIK3CA |
| microT-CDS | 5.74 × 103 | NRAS | ||
| miR-125b-5p | Tarbase | 1.80 × 1020 | ERBB2,RAF1, AXIN1,TP53, CTNNB1, CTNNA1,AKT1 | |
| TargetScan | 3.60 × 1011 | PIK3CB, PIK3R5,PIK3CD,TCF7 | ||
| microT-CDS | 5.74 × 1003 | PIK3CB | ||
| miR-23b-3p | Tarbase | 6.13 × 1034 | RAF1, EGFR, AKT2, CCND1, CTNNB1, MYC, SOS1, FOXO3, PDPK1,PTEN, MAPK1 | |
| TargetScan | 2.79 × 1017 | GSK3B, PIK3R3, CCND1, SOS1, PDPK1, PTEN | ||
| microT-CDS | 3.54 × 1014 | BRAF, PIK3CB, PIK3R3, PDPK1, PTEN | ||
| miR-99a-5p | Tarbase | 5.74 × 103 | CCND1 | |
| Endometrioid vs. Serous | let-7g-5p | Tarbase | 4.49 × 1027 | BRAF, NRAS,KRAS, MLH1, TP53, CCND1, CTNNA1,MYC, MAPK1 |
| TargetScan | 1.80 × 1020 | NRAS, PIK3R5, TP53,AKT2, CCND1, APC2, PIK3CA | ||
| microT-CDS | 5.74 × 103 | NRAS | ||
| miR-195-5p | Tarbase | 1.80 × 1020 | CDH1,CCND1, CTNNB1, MYC, AKT3, PIK3CA, GRB2 | |
| TargetScan | 4.89 × 1041 | GSK3B, SOS2, PIK3R5,RAF1, EGFR, KRAS, CDH1,CCND1, AXIN2, PIK3R1, SOS1, AKT3, MAP2K1 | ||
| microT-CDS | 9.75 × 1024 | BRAF, SOS2,RAF1,CCND1, AXIN2, PIK3R1, AKT3, MAP2K1 | ||
| miR-34a-5p | Tarbase | 1.11 × 1083 | BRAF, MAP2K2, PIK3R2, TCF7L1, RAF1, EGFR, ARAF,TP53, AKT2, CDH1,CCND1, CTNNB1,AXIN2,MYC, MAPK3, CASP9, PIK3CA,LEF1,MAP2K1, PTEN, MAPK1, GRB2, BAD, ELK1 | |
| TargetScan | 2.82 × 108 | CCND1,LEF1,MAP2K1 | ||
| microT-CDS | 3.54 × 1014 | ERBB2, PIK3CB,AXIN2,LEF1,MAP2K1 | ||
| miR-497-5p | Tarbase | 1.78 × 1030 | SOS2, PIK3R2, AKT2, CCND1, MYC, AKT3, PIK3CA,MAP2K1, MAPK1, GRB2 | |
| TargetScan | 4.89 × 1041 | GSK3B, SOS2, PIK3R5,RAF1, EGFR, KRAS, CDH1, CCND1, AXIN2, PIK3R1, SOS1, AKT3, MAP2K1 | ||
| microT-CDS | 9.75 × 1024 | BRAF, GSK3B, SOS2,RAF1, AXIN2, PIK3R1, AKT3,MAP2K1 |
| Endometrioid Grade 1 vs. Grade 3 | Endometrioid vs. Serous | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Target Genes | let-7c-5p | miR-125b-5p | miR-23b-3p | miR-99a-5p | let-7g-5p | miR-195-5p | miR-34a-5p | miR-497-5p | Number of Targeting miRNAs |
| FOXO3 | Yes | 1 | |||||||
| PIK3R3 | Yes | 1 | |||||||
| AXIN1 | Yes | 1 | |||||||
| PIK3CD | Yes | 1 | |||||||
| TCF7 | Yes | 1 | |||||||
| AKT1 | Yes | 1 | |||||||
| PDPK1 | Yes | 1 | |||||||
| APC2 | Yes | Yes | 2 | ||||||
| NRAS | Yes | Yes | 2 | ||||||
| CTNNA1 | Yes | Yes | 2 | ||||||
| ERBB2 | Yes | Yes | 2 | ||||||
| PTEN | Yes | Yes | 2 | ||||||
| GSK3B | Yes | Yes | Yes | 3 | |||||
| PIK3CB | Yes | Yes | Yes | 3 | |||||
| SOS1 | Yes | Yes | Yes | 3 | |||||
| EGFR | Yes | Yes | Yes | Yes | 4 | ||||
| MAPK1 | Yes | Yes | Yes | Yes | 4 | ||||
| TP53 | Yes | Yes | Yes | Yes | 4 | ||||
| AKT2 | Yes | Yes | Yes | Yes | Yes | 5 | |||
| CTNNB1 | Yes | Yes | Yes | Yes | Yes | 5 | |||
| MYC | Yes | Yes | Yes | Yes | Yes | 5 | |||
| RAF1 | Yes | Yes | Yes | Yes | Yes | 5 | |||
| BRAF | Yes | Yes | Yes | Yes | Yes | 5 | |||
| PIK3R5 | Yes | Yes | Yes | Yes | Yes | 5 | |||
| PIK3CA | Yes | Yes | Yes | Yes | Yes | 5 | |||
| CCND1 | Yes | Yes | Yes | Yes | Yes | Yes | 6 | ||
| CDH1 | Yes | Yes | Yes | 3 | |||||
| MAP2K1 | Yes | Yes | Yes | 3 | |||||
| GRB2 | Yes | Yes | Yes | 3 | |||||
| KRAS | Yes | Yes | Yes | 3 | |||||
| AXIN2 | Yes | Yes | Yes | 3 | |||||
| SOS2 | Yes | Yes | 2 | ||||||
| PIK3R2 | Yes | Yes | 2 | ||||||
| AKT3 | Yes | Yes | 2 | ||||||
| PIK3R1 | Yes | Yes | 2 | ||||||
| MAP2K2 | Yes | 1 | |||||||
| TCF7L1 | Yes | 1 | |||||||
| ARAF | Yes | 1 | |||||||
| MAPK3 | Yes | 1 | |||||||
| CASP9 | Yes | 1 | |||||||
| LEF1 | Yes | 1 | |||||||
| BAD | Yes | 1 | |||||||
| ELK1 | Yes | 1 | |||||||
| MLH1 | Yes | 1 | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalinkova, L.; Kajo, K.; Karhanek, M.; Wachsmannova, L.; Suran, P.; Zmetakova, I.; Fridrichova, I. Discriminating miRNA Profiles between Endometrioid Well- and Poorly-Differentiated Tumours and Endometrioid and Serous Subtypes of Endometrial Cancers. Int. J. Mol. Sci. 2020, 21, 6071. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176071
Kalinkova L, Kajo K, Karhanek M, Wachsmannova L, Suran P, Zmetakova I, Fridrichova I. Discriminating miRNA Profiles between Endometrioid Well- and Poorly-Differentiated Tumours and Endometrioid and Serous Subtypes of Endometrial Cancers. International Journal of Molecular Sciences. 2020; 21(17):6071. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176071
Chicago/Turabian StyleKalinkova, Lenka, Karol Kajo, Miloslav Karhanek, Lenka Wachsmannova, Peter Suran, Iveta Zmetakova, and Ivana Fridrichova. 2020. "Discriminating miRNA Profiles between Endometrioid Well- and Poorly-Differentiated Tumours and Endometrioid and Serous Subtypes of Endometrial Cancers" International Journal of Molecular Sciences 21, no. 17: 6071. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176071