Quantification of Iron Release from Native Ferritin and Magnetoferritin Induced by Vitamins B2 and C

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
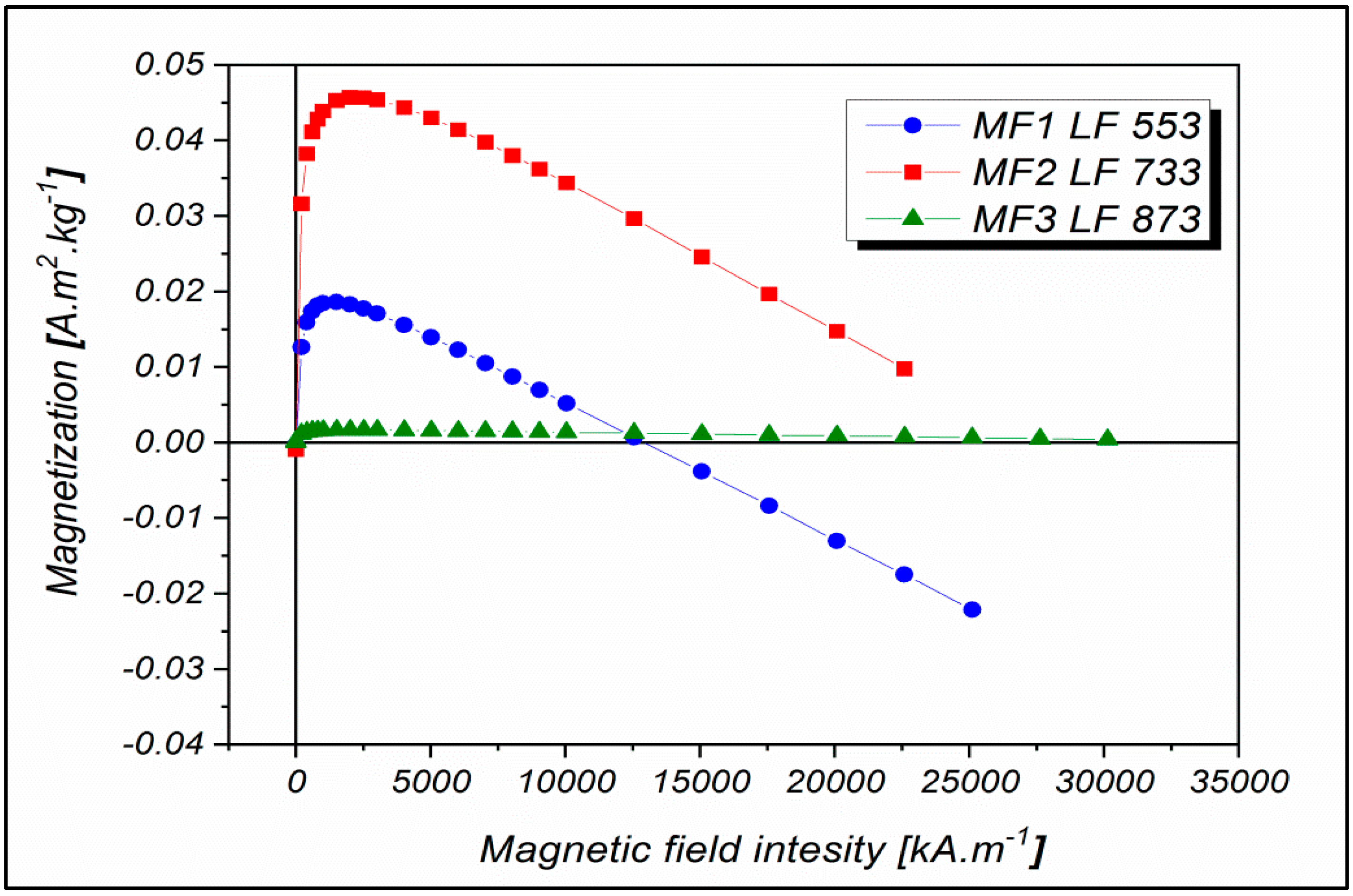
2.1. Magnetoferritin Characterization
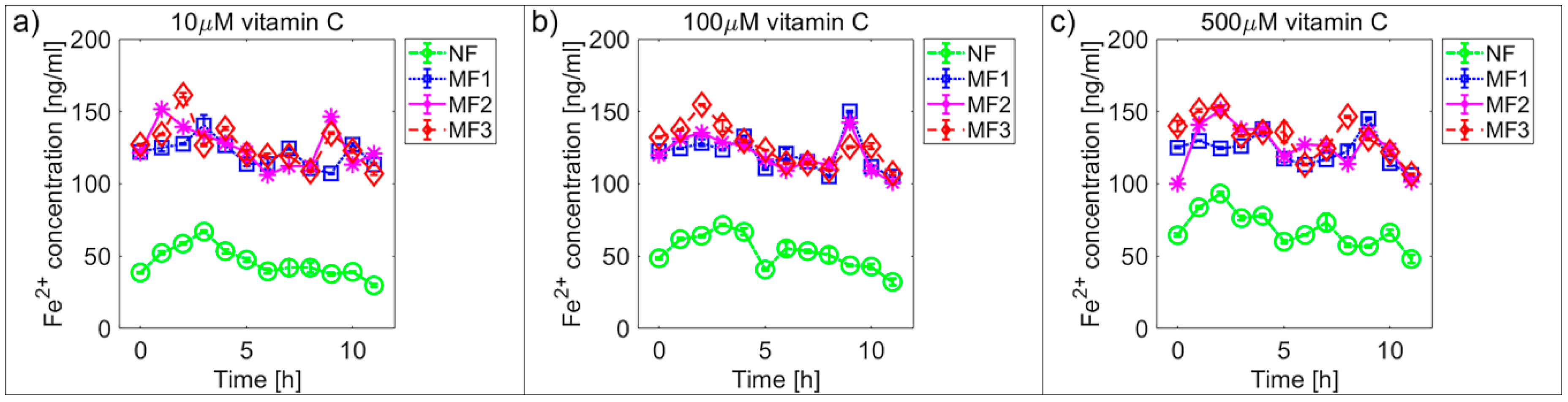
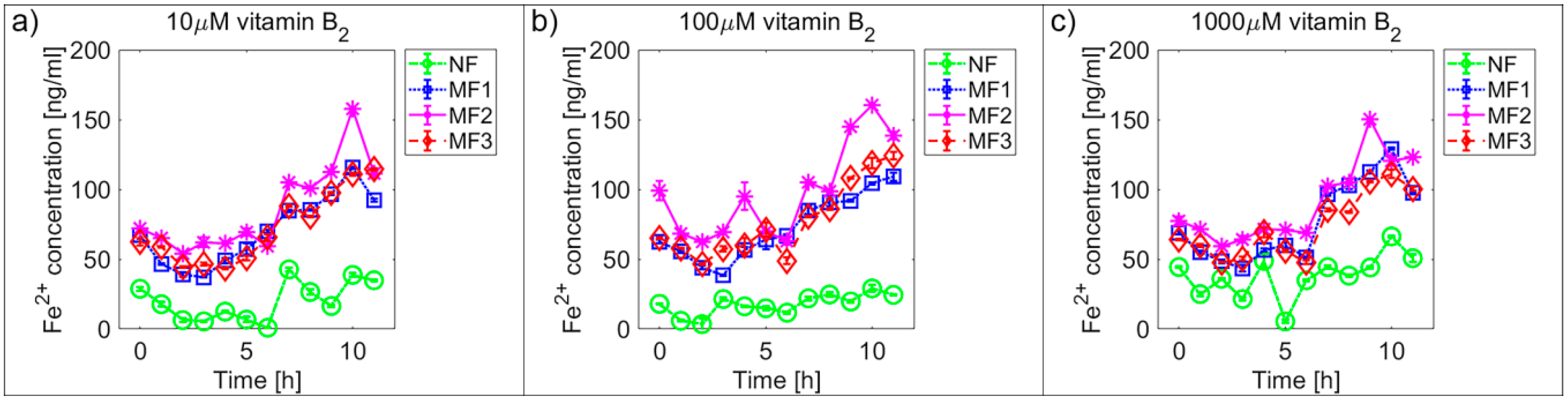
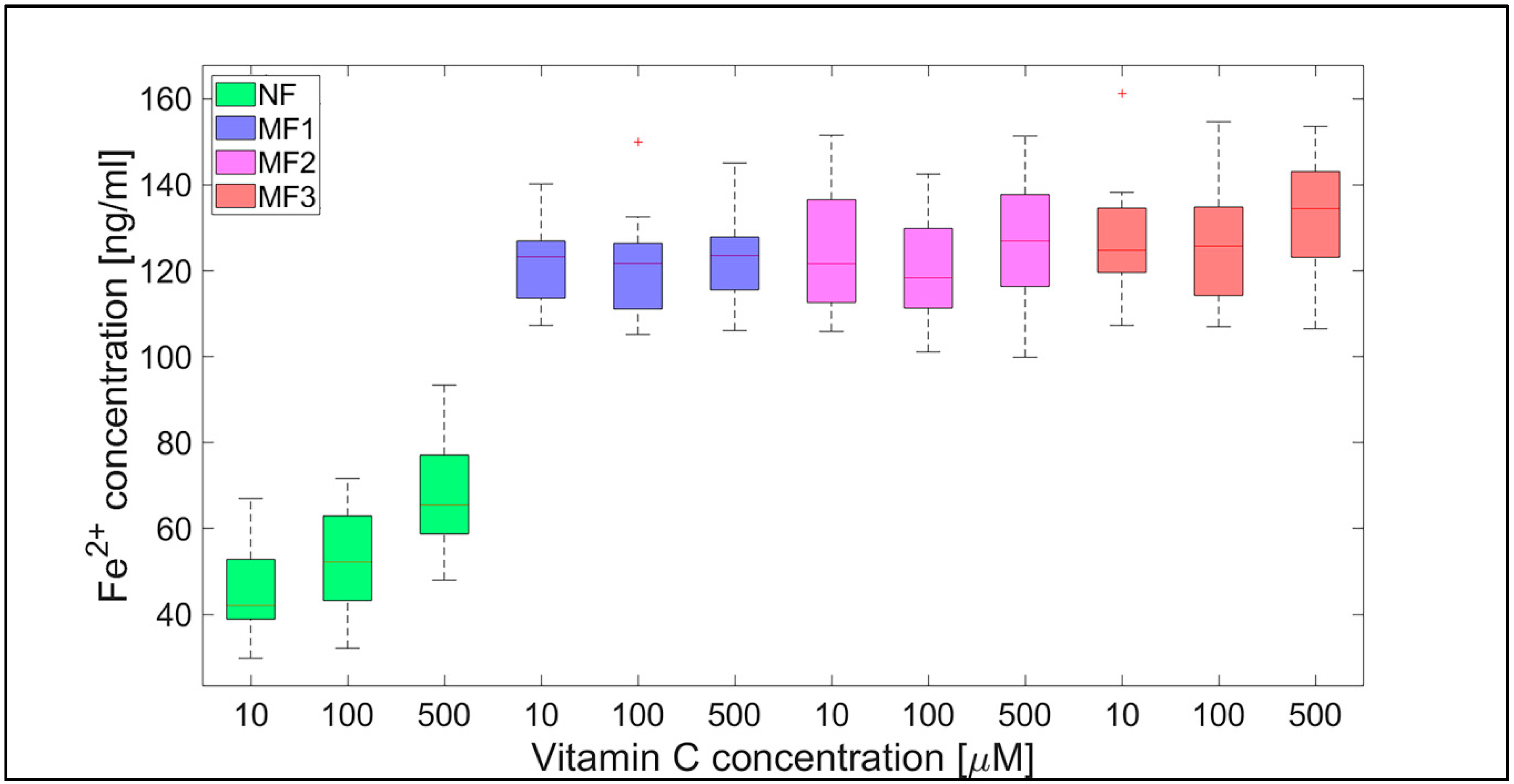
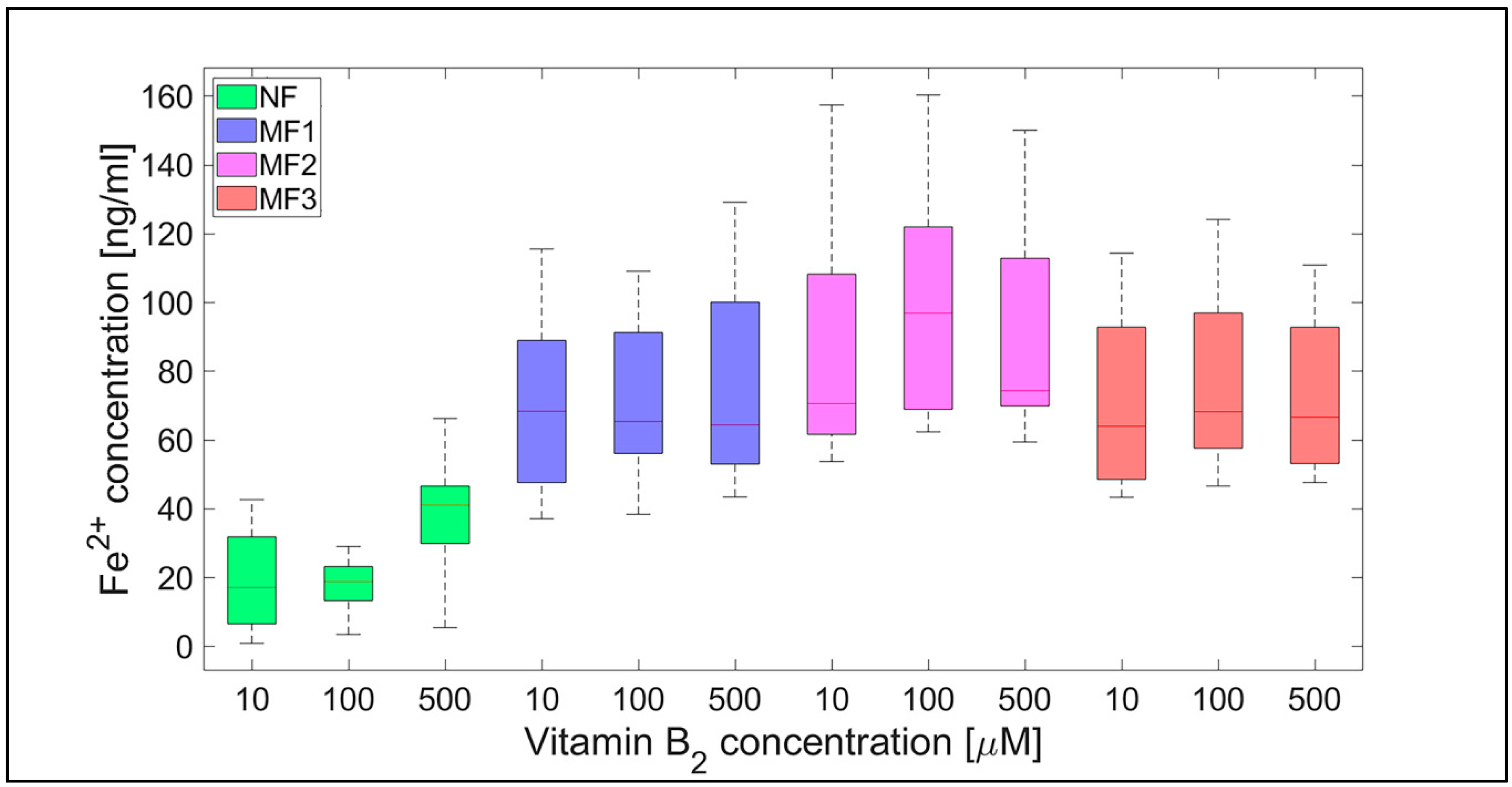
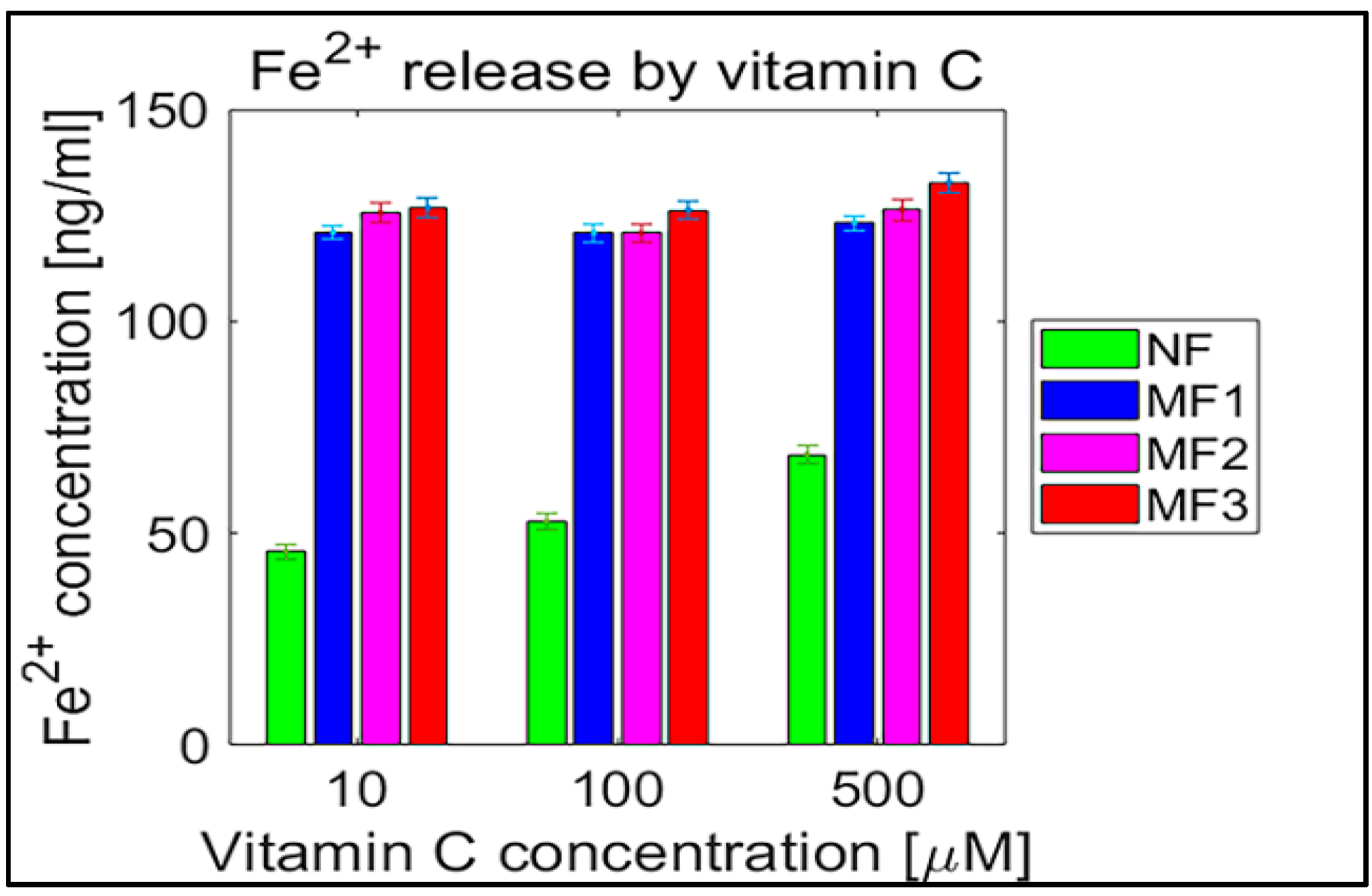
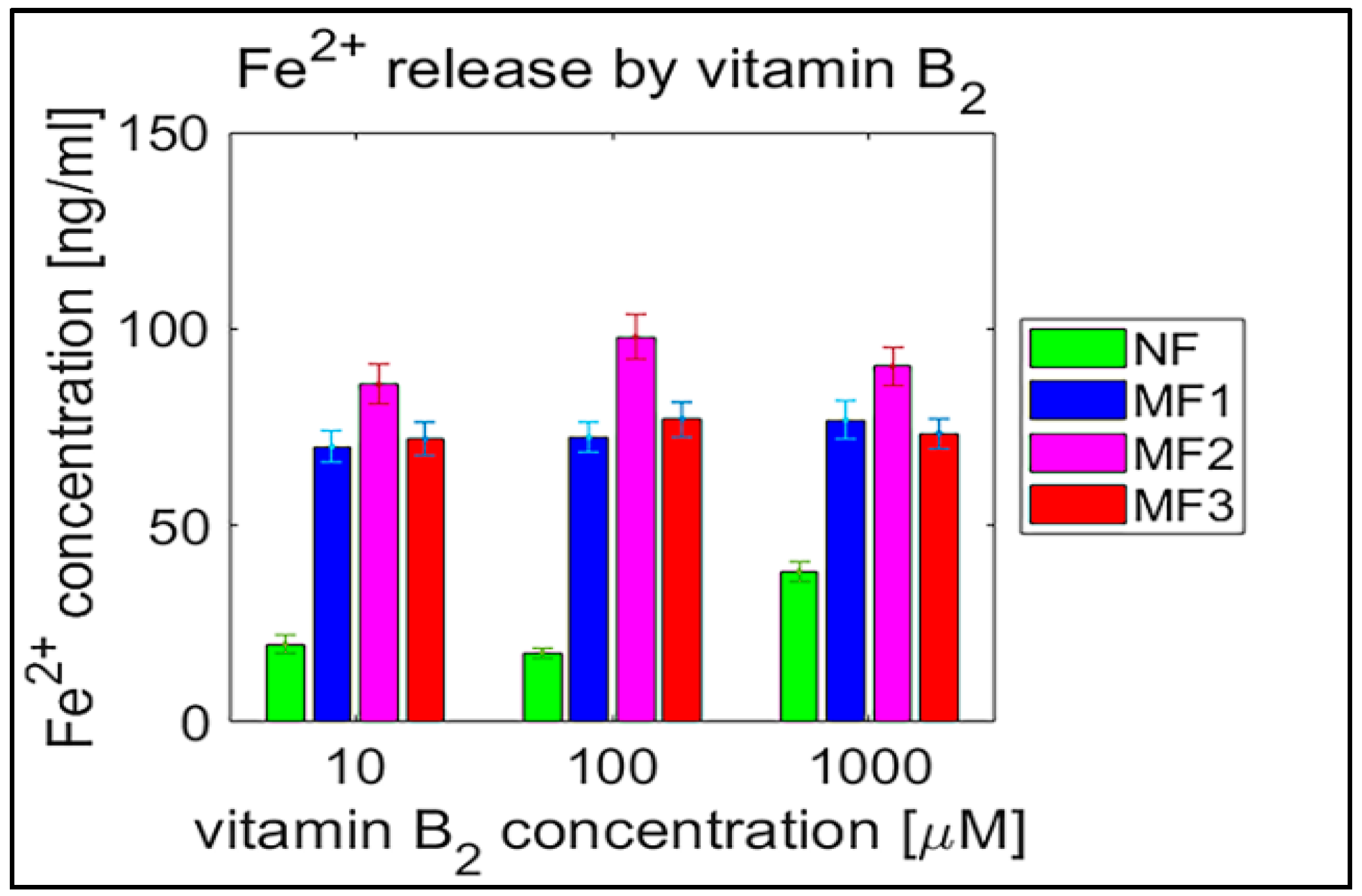
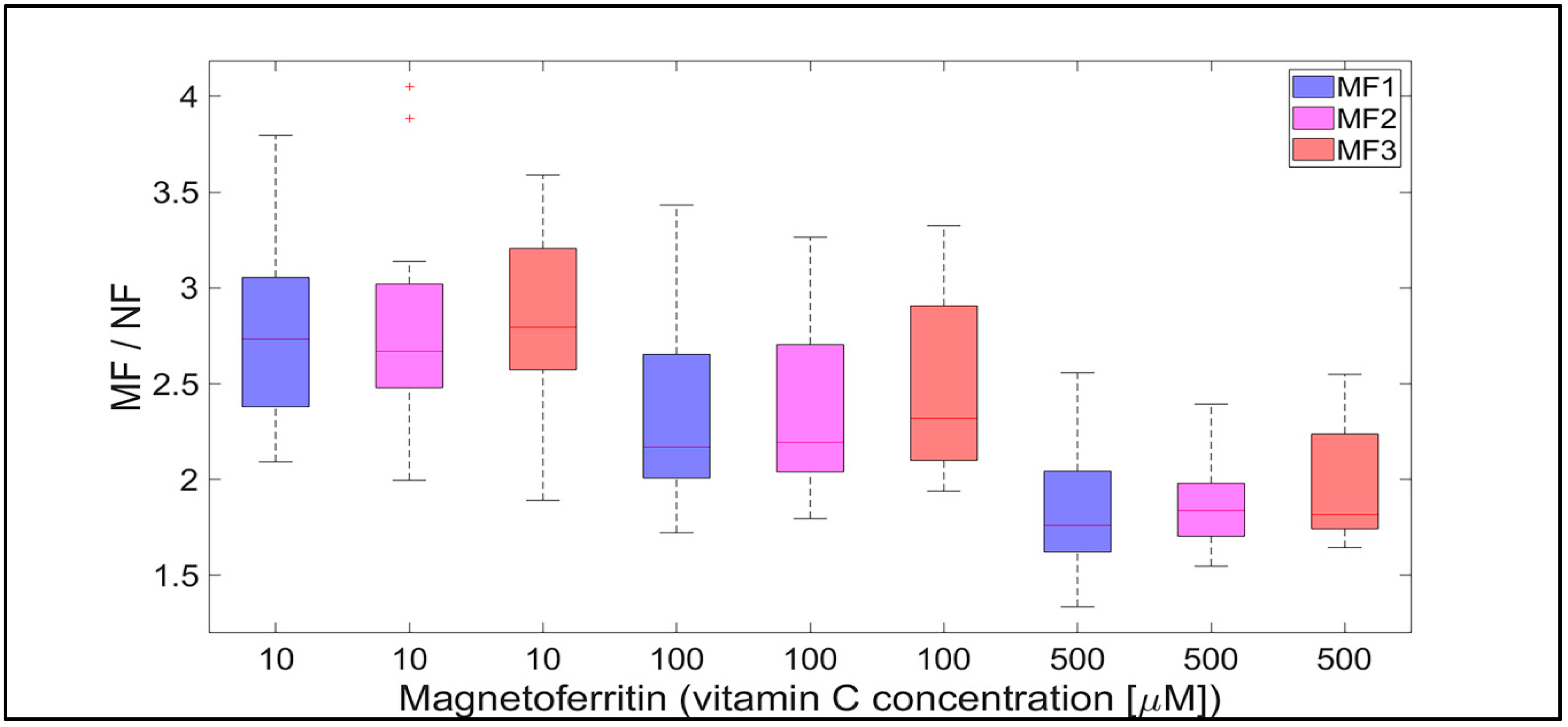
2.2. Mineral Core Reduction
3. Discussion
- Biologically essential vitamins (B2 and C) are capable of ferritin mineral core reduction at a rate that exceeds the rate of iron release from “untouched” ferritin by 20 to 68 times per second. From the long-term point of view, this can result in the accumulation of toxic iron (ferrous ions) in cells, giving rise to a higher risk of oxidative stress.
3.1. A New Proposal for the Origin of Magnetite in Human Tissue and the Initiation of Associated Pathology
4. Materials and Methods
4.1. Chemicals
4.2. Synthesis of Magnetoferritin
4.3. Quantitative Determination of the Loading Factor
4.4. Measurement of Hydrodynamic Diameters
4.5. Zeta Potential Determination
4.6. SQUID Magnetometry
4.7. The Study of Ferritin and Its Derivates’ Interaction with Ascorbic Acid and Riboflavin
4.8. Data Processing and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Williams, R.J.P. Iron and the origin of life. Nature 1990, 343, 213–214. [Google Scholar] [CrossRef] [PubMed]
- Archibald, F.S. Lactobacillus plantarum, an organism not requiring iron. FEMS Microbiol. Let. 1983, 19, 19–32. [Google Scholar] [CrossRef]
- Ilbert, M.; Bonnefoy, V. Insight into the evolution of the iron oxidation pathways. Biochim. Biophys. Acta 2013, 1827, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hentze, M.W.; Muckenthaler, M.U.; Andrews, N.C. Balancing acts: Molecular control of mammalian iron metabolism. Cell 2004, 117, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Frank-Cannon, T.C.; Alto, L.T.; McAlpine, F.E.; Tansey, M.G. Does neuroinflammation fan the flame in neurodegenerative disease? Mol. Neurodegener. 2009, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Hagemeier, J.; Geurts, J.J.G.; Zivadinov, R. Brain iron accumulation in ageing and neurodegenerative disorders. Expert. Rev. Neurother. 2012, 12, 1467–1480. [Google Scholar] [CrossRef]
- Kobayashi, M.; Suhara, T.; Baba, Y.; Kawasaki, N.K.; Higa, J.K.; Matsui, T. Pathological roles of iron in cardiovascular disease. Curr. Drug Targets 2018, 19, 1068–1076. [Google Scholar] [CrossRef]
- Jung, M.; Mertens, C.; Tomat, E.; Brune, B. Iron as a central player and promising target in cancer progression. Int. J. Mol. Sci. 2019, 20, 273. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Pantooulos, K. Regulation of cellular iron metabolism. Biochem. J. 2011, 434, 365–381. [Google Scholar] [CrossRef] [Green Version]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Das, T.K.; Wati, M.R.; Fatima-Shad, K. Oxidative stress gated by Fenton and Haber Weiss reactions and its association with Alzheimer’s disease. Arch. Neurosci. 2014, 2, e20078. [Google Scholar]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Knovich, M.A.; Storey, J.A.; Coffman, L.G.; Torti, S.V. Ferritin for the clinicians. Blood Rev. 2009, 23, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Stumm, W.; Lee, G.F. Oxygenation of ferrous iron. Ind. Eng. Chem. 1961, 53, 143–146. [Google Scholar] [CrossRef]
- Bjorklund, G.; Hofer, T.; Nurchi, V.M.; Aaseth, J. Iron and other metals in the pathogenesis of Parkinson’s disease: Toxic effects and possible detoxification. J. Inorg. Biochem. 2019, 199, 110717. [Google Scholar] [CrossRef] [PubMed]
- Hautot, D.; Pankhurst, Q.A.; Khan, N.; Dobson, J. Preliminary evaluation of nanoscale biogenic magnetite in Alzheimer’s disease brain tissue. Proc. R. Soc. Lond. B 2003, 270, S62–S64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschvink, J.L.; Kobayashi-Kirschvink, A.; Woodford, B.J. Magnetite biomineralization in the human brain. Proc. Natl. Acad. Sci. USA 1992, 89, 7683–7687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prozorov, T. Magnetic microbes: Bacterial magnetite biomineralization. Semin. Cell Dev. Biol. 2015, 46, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Dobson, J. On the structural form of iron in ferritin cores associated with progressive supranuclear palsy and Alzheimer’s disease. Cell. Mol. Biol. 2001, 47, OL49–OL50. [Google Scholar]
- Quintana, C.; Gutierrez, L. Could a dysfunction of ferritin be a determinant factor in the aetiology of some neurodegenerative disease? Biochim. Biophys Acta 2010, 1800, 770–782. [Google Scholar] [CrossRef]
- Chasteen, N.D.; Harrison, P.M. Mineralization in ferritin: An efficient means of iron storage. J. Struct. Biol. 1999, 126, 182–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bou-Abdallah, F.; Zhao, G.; Biasiotto, G.; Poli, M.; Arosio, M.; Chasteen, N.D. Facilitated diffusion of iron(II) and dioxygen substrates into human H-chain ferritin. A fluorescence and absorbance study employing the ferroxidase center substitution Y34W. J. Am. Chem. Soc. 2008, 130, 17801–17811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theil, E.C. Ferritin: The protein nanocage and iron biomineral in health and in disease. Inorg. Chem. 2013, 52, 12223–12233. [Google Scholar] [CrossRef] [PubMed]
- Quintana, C.; Cowley, J.M.; Marhic, C. Electron nanodiffraction and high-resolution electron microscopy studies of the structure and composition of physiological and pathological ferritin. J. Struct. Biol. 2004, 147, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Bossoni, L.; Moursel, L.G.; Bulk, M.; Simon, B.G.; Webb, A.; Weerd, A.; Weerd, L.; Huber, M.; Carretta, P.; Lascialfari, A.; et al. Human-brain ferritin studied by muon spin rotation: A pilot study. J. Phys.: Condens. Matter 2017, 29, 415801. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Deng, D.; Sun, J. Magnetoferritin: Process, prospects, and their biomedical applications. Int. J. Mol. Sci. 2019, 20, 2426. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.; Spencer, R.; Walsh, C. Mechanism and kinetics of iron release from ferritin by dihydroflavins and dihydroflavin analogs. Biochemistry 1978, 17, 4011–4017. [Google Scholar] [CrossRef] [PubMed]
- Sirivech, S.; Frieden, E.; Osaki, S. The release of iron from horse spleen ferritin by reduced flavins. Biochem. J. 1974, 143, 311–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, R.F.; Grabill, T.W.; Petrovich, R.M. Reductive release of ferritin iron: A kinetic assay. Anal. Biochem. 1988, 174, 17–22. [Google Scholar] [CrossRef]
- Sakurai, K.; Nabeyama, A.; Fujimoto, Y. Ascorbate-mediated iron release from ferritin in the presence of alloxan. Reductive mobilisation of ferritin iron. FEBS J. 1985, 152, 167–172. [Google Scholar]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophys. Acta 2012, 1826, 443–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melman, G.; Bou-Abdallah, F.; Vane, E.; Maura, P.; Arosio, P.; Melman, A. Iron release from ferritin by flavin nucleotides. Biochim. Biophys. Acta 2013, 1830, 4669–4674. [Google Scholar] [CrossRef] [PubMed]
- Hagel, A.F.; Albrecht, H.; Dauth, W.; Hagel, W.; Vitali, F.; Ganzleben, I.; Schultis, H.W.; Konturek, P.C.; Stein, J.; Neurath, M.F.; et al. Plasma concentrations of ascorbic adic in a cross section of the German population. J. Int. Med. Res. 2018, 46, 168–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischbach, F.T.; Fischbach, M.A. A Manual of Laboratory and Diagnostic Tests, 10th ed.; Wolters Kluwer: Alphen aan den Rijn, The Netherlands, 2018. [Google Scholar]
- Orino, K.; Kamura, S.; Natsuhori, M.; Yamamoto, S.; Watanabe, K. Two pathways of iron uptake in bovine spleen apoferritin dependent on iron concentration. Biometals 2002, 15, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Arosio, P.; Levi, S.; Chasteen, N.D. Ferroxidase kinetics of human liver apoferritin, recombinant H-chain apoferritin, and site-directed mutants. Biochemistry 1993, 32, 9362–9369. [Google Scholar] [CrossRef] [PubMed]
- Lui, X.; Theil, E.C. Ferritin reactions: Direct identification of the site for the diferric peroxide reaction intermediate. Proc. Natl. Acad. Sci. USA 2004, 101, 8557–8562. [Google Scholar]
- Liu, X.S.; Patterson, L.D.; Miller, M.J.; Theil, E.C. Peptides selected for the protein nanocage pores change the rate of iron recovery from the ferritin mineral. J. Biol. Chem. 2007, 282, 31821–31825. [Google Scholar] [CrossRef] [Green Version]
- Balejcikova, L.; Molcan, M.; Kovac, J.; Kubovcikova, M.; Saksl, K.; Mitroova, Z.; Timko, M.; Kopcansky, P. Hyperthermic effect in magnetoferritin aqueous colloidal solution. J. Mol. Liq. 2019, 283, 39–44. [Google Scholar] [CrossRef]
- Strbak, O.; Balejcikova, L.; Baciak, L.; Kovac, J.; Masarova-Kozelova, M.; Krafcik, A.; Dobrota, D.; Kopcansky, P. Low-field and high-field magnetic resonance contrast imaging of magnetoferritin as a pathological model system of iron accumulation. J. Phys. D Appl. Phys. 2017, 50, 365401. [Google Scholar] [CrossRef]
- Wong, K.K.W.; Douglas, T.; Gider, S.; Awschalom, D.D.; Mann, S. Biomimetic synthesis and characterisation of magnetic proteins (magnetoferritin). Chem. Mater. 1998, 10, 279–285. [Google Scholar] [CrossRef]
- Suwannasom, N.; Kao, I.; Prus, A.; Georgieva, R.; Baumler, H. Riboflavin: The health benefits of a forgotten natural vitamin. Int. J. Mol. Sci. 2020, 21, 950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Tu, H.; Wang, Y.; Levine, M. Vitamin C in mouse and human red blood cells: An HPLC assay. Anal. Biochem. 2012, 426, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.H.; Folson, A.R.; Harnack, L.; Halliwell, B.; Jacobs, D.R., Jr. Does supplemental vitamin C increase cardiovascular disease risk in women with diabetes? Am. J. Clin. Nutr. 2004, 80, 1194–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liguori, I.; Russo, G.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; Abete, P. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoll, A.H.; Bergmann, K.D.; Strauss, J.V. Life: The first two billion years. Phil. Trans. R. Soc. B 2016, 371, 20150493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A red carpet for iron metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Mercadante, C.J.; Prajapati, M.; Parmar, J.H.; Conboy, H.L.; Dash, M.E.; Pettiglio, M.A.; Herrera, C.; Bu, J.T.; Stopa, E.G.; Mendes, P.; et al. Gastrointestinal iron excretion and reversal of iron excess in a mouse model of inherited iron excess. Haematologica 2019, 104, 678–689. [Google Scholar] [CrossRef]
- James, S.A.; Roberts, B.R.; Hare, D.J.; de Jonge, M.D.; Birchall, I.E.; Jenkins, N.L.; Cherry, R.A.; Bush, A.I.; McColl, G. Direct in vivo imaging of ferrous iron dyshomeostasis in ageing Caenorhabditis elegans. Chem. Sci. 2015, 6, 2952. [Google Scholar] [CrossRef] [Green Version]
- Saito, H. Metabolism of iron stores. Nagoya J. Med. Sci. 2014, 76, 235–254. [Google Scholar]
- Cabantchik, Z.I. Labile iron in cells and body fluids: Physiology, pathology, and pharmacology. Front. Pharmacol. 2014, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Dugger, B.N.; Dickson, D.W. Pathology of neurodegenerative diseases. Cold Spring Harb. Perspect Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [PubMed]
- Maher, B.A.; Ahmed, I.A.M.; Karloukovski, V.; MacLaren, D.A.; Foulds, P.G.; Allsop, D.; Mann, D.M.A.; Torres-Jardon, R.; Calderon-Garcduenas, L. Magnetite pollution nanoparticles in the human brain. Proc. Natl. Acad. Sci. USA 2016, 113, 10797–10801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfsen, E.M.; Stormer, F.C.; Nja, A.; Walloe, L. A proposed tandem mechanism for memory storage in neurons involving magnetite and prions. Med. Hypotheses 2018, 119, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | LF | <DHYDR> [nm] | ζ Potential [mV] |
|---|---|---|---|
| MF1 | 553 | 46.00 | −29.3 |
| MF2 | 733 | 46.45 | −26.9 |
| MF3 | 873 | 42.20 | −20.8 |
| NF | 868 | 15.60 | −30.8 |
| Sample | Vitamin C [µM] | Vitamin B2 [µM] | ||||
|---|---|---|---|---|---|---|
| 10 | 100 | 500 | 10 | 100 | 1000 | |
| NF | 45.7 ± 0.9 | 52.7 ± 1.0 | 68.6 ± 1.1 | 19.9 ± 1.2 | 17.7 ± 0.6 | 38.3 ± 1.3 |
| MF1 | 121.0 ± 0.8 | 120.9 ± 1.1 | 123.3 ± 0.9 | 70.0 ± 2.1 | 72.4 ± 1.9 | 76.8 ± 2.4 |
| MF2 | 125.6 ± 1.2 | 120.8 ± 1.0 | 126.3 ± 1.3 | 85.9 ± 2.6 | 98.0 ± 2.8 | 90.4 ± 2.4 |
| MF3 | 126.7 ± 1.2 | 126.2 ± 1.2 | 132.7 ± 1.2 | 72.0 ± 2.1 | 77.0 ± 2.2 | 73.4 ± 1.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strbak, O.; Balejcikova, L.; Kmetova, M.; Gombos, J.; Trancikova, A.; Pokusa, M.; Kopcansky, P. Quantification of Iron Release from Native Ferritin and Magnetoferritin Induced by Vitamins B2 and C. Int. J. Mol. Sci. 2020, 21, 6332. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176332
Strbak O, Balejcikova L, Kmetova M, Gombos J, Trancikova A, Pokusa M, Kopcansky P. Quantification of Iron Release from Native Ferritin and Magnetoferritin Induced by Vitamins B2 and C. International Journal of Molecular Sciences. 2020; 21(17):6332. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176332
Chicago/Turabian StyleStrbak, Oliver, Lucia Balejcikova, Martina Kmetova, Jan Gombos, Alzbeta Trancikova, Michal Pokusa, and Peter Kopcansky. 2020. "Quantification of Iron Release from Native Ferritin and Magnetoferritin Induced by Vitamins B2 and C" International Journal of Molecular Sciences 21, no. 17: 6332. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176332