In Vitro Antagonistic Effect of Gut Bacteriota Isolated from Indigenous Honey Bees and Essential Oils against Paenibacillus Larvae

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results
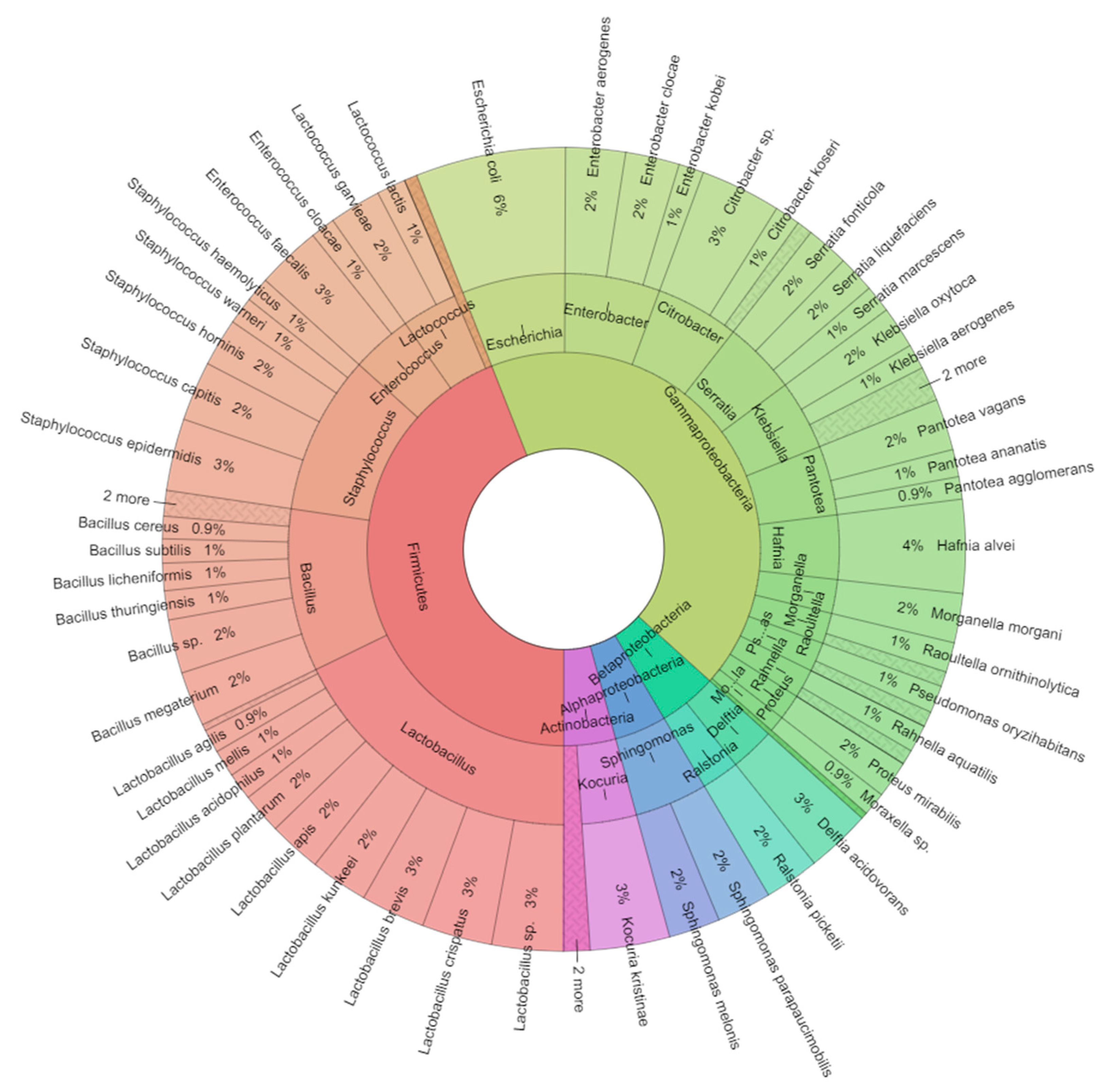
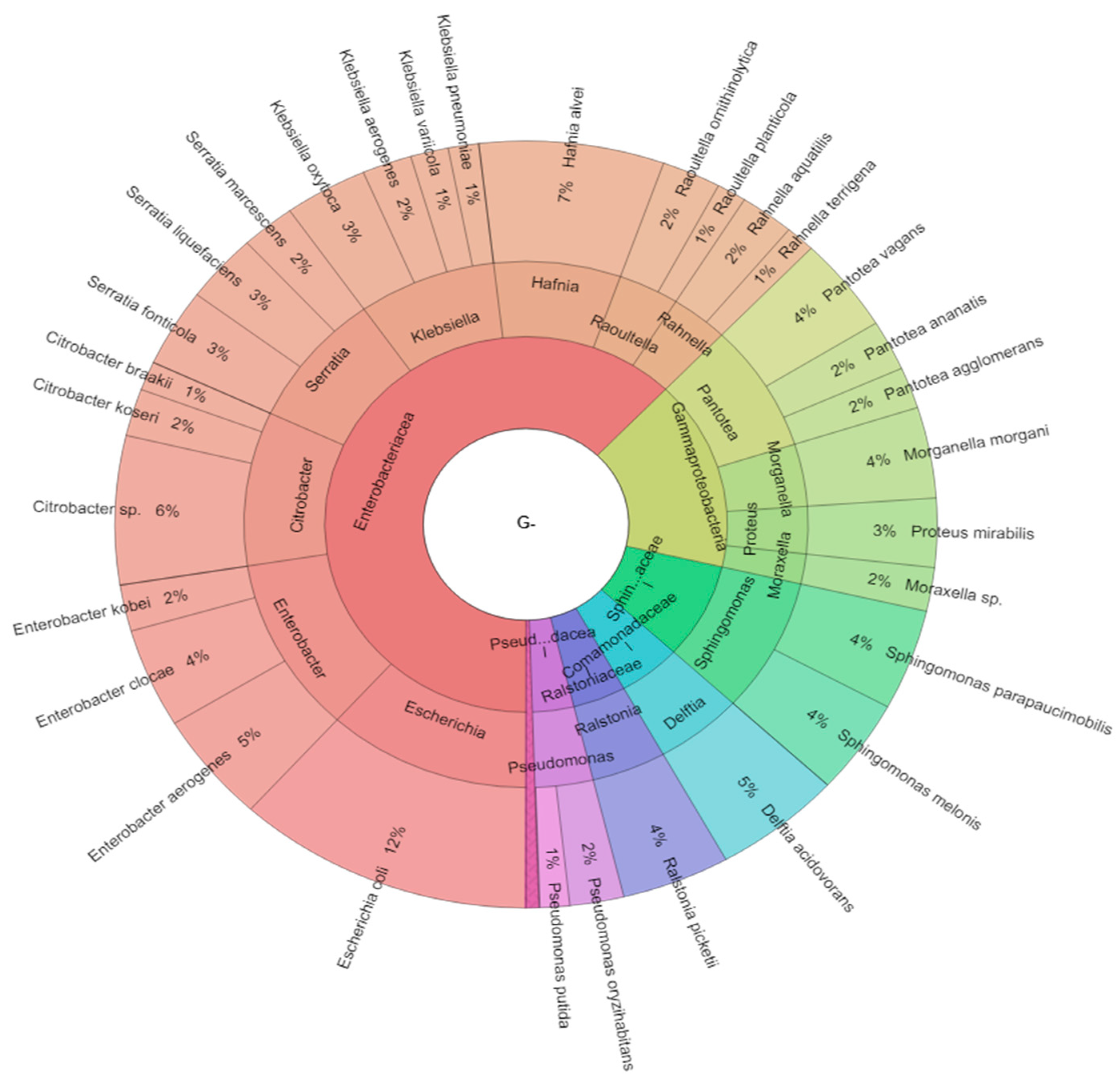
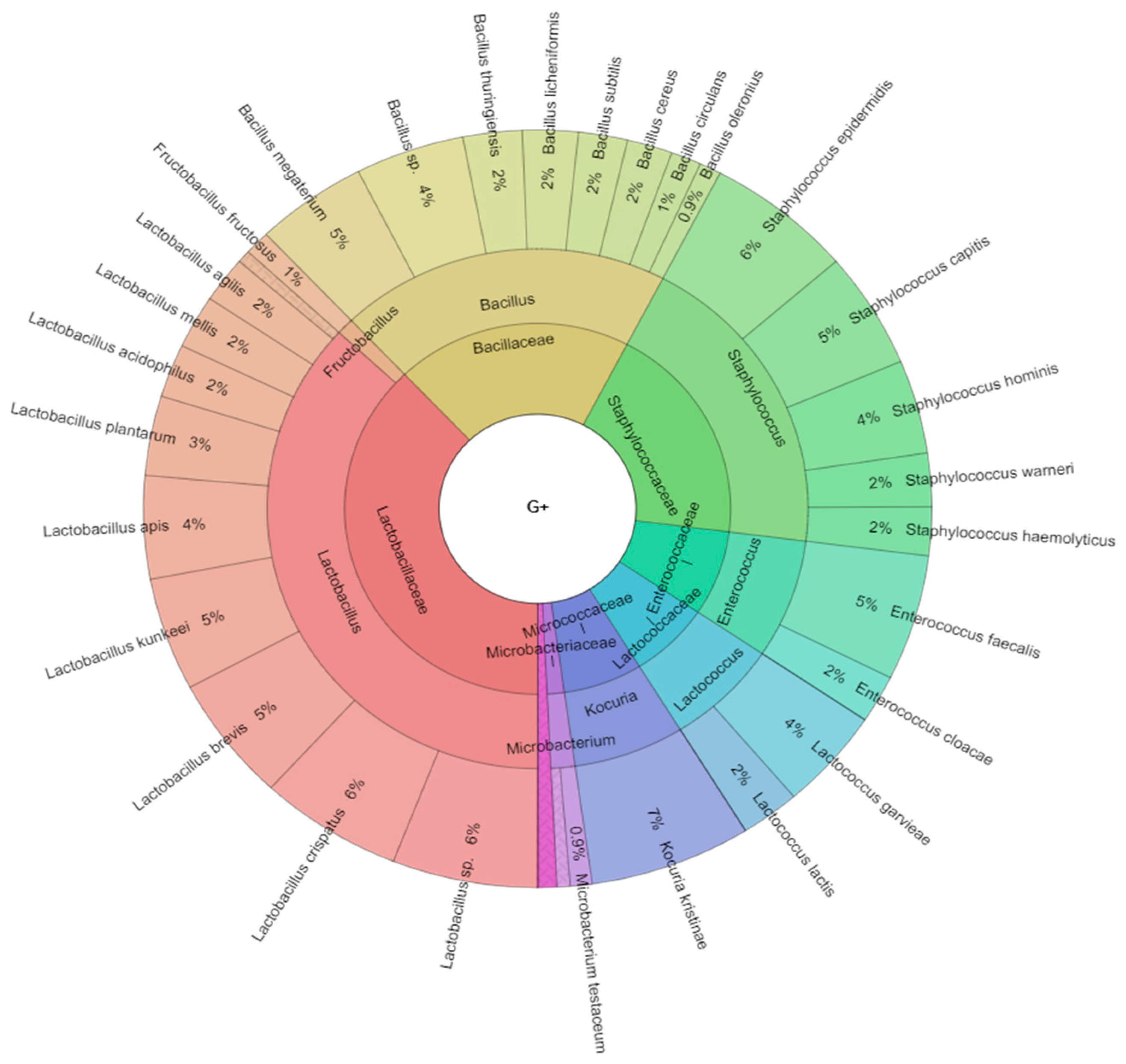
2.1. Bacteriota of Adult Worker Bees (Apis mellifera)
2.2. Isolated Bacteria from Bees Gut
2.3. Antibiotic Resistance of A. mellifera Gut Bacteriota
2.4. Antimicrobial Activity of Isolated Bee Digestive Tract Bacteriome against P. larvae
2.5. Antimicrobial Activity of Essential Oils against P. larvae
3. Discussion
4. Materials and Methods
4.1. Samples of Bees
4.2. Identification of Bacteria
4.3. Antimicrobial Resistance Testing
4.4. Antimicrobial Activity of Bacterial Suspensions against P. larvae
4.5. Antimicrobial Activity of Essential Oils against P. larvae
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jeyaprakash, A.; Hoy, M.A.; Allsopp, M.H. Bacterial diversity in worker adults of Apis mellifera capensis and Apis mellifera scutellata (Insecta: Hymenoptera) assessed using 16S rRNA sequences. J. Invertebr. Pathol. 2003, 84, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Tysset, C.; Durand, C. Contribution to the study of the intestinal microbism of healthy worker bees (Apis mellifica); reckoning and study of the constitutive groups. Bull Apic 1968, 2, 117–118. [Google Scholar]
- Tysset, G.; Rousseau, M.; Durand, G. La présence des streptocoques du groupe D de Lancefield chez les abeilles butineuses saines (Alpis mellifica L.) L’interprétation de leur présence en bactériologie alimentaire. Bull. Acad. Vet. Fr. 1969, XLII, 173–186. [Google Scholar] [CrossRef] [Green Version]
- Scardovi, V. Genus Bifidobacterium. In Bergey’s Manual of Systematic Bacteriology; Williams and Wilkins: Baltimore, MD, USA, 1986. [Google Scholar]
- Miyagi, T.; Peng, C.Y.S.; Chuang, R.Y.; Mussen, E.C.; Spivak, M.S.; Doi, R.H. Verification of Oxytetracycline-Resistant American Foulbrood Pathogen Paenibacillus larvae in the United States. J. Invertebr. Pathol. 2000, 75, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Shimanuki, H.; Knox, D.A. Susceptibility of Bacillus larvae to Terramycin. Am. bee J. 1994, 134, 125–126. [Google Scholar]
- Michaud, V. Antibiotic residues in honey-the FEEDM view. Apiacta 2005, 40, 52–54. [Google Scholar]
- Iwu, M.W.; Duncan, A.R.; Okunji, C.O. New antimicrobials of plant origin. In Perspectives on New Crops and New Uses; ASHS Press: Alexandria, VA, USA, 1999; pp. 457–462. [Google Scholar]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Rada, V.; Máchová, M.; Huk, J.; Marounek, M.; Dušková, D. Microflora in the honeybee digestive tract: Counts, characteristics and sensitivity to veterinary drugs. Apidologie 1997, 28, 357–365. [Google Scholar] [CrossRef] [Green Version]
- Killer, J.; Kopečný, J.; Mrázek, J.; Rada, V.; Dubná, S.; Marounek, M. Bifidobacteria in the digestive tract of bumblebees. Anaerobe 2010, 16, 165–170. [Google Scholar] [CrossRef]
- Kačániová, M.; Chlebo, R.; Kopernický, M.; Trakovická, A. Microflora of the honeybee gastrointestinal tract. Folia Microbiol. (Praha) 2004, 49, 169–171. [Google Scholar] [CrossRef]
- Grubbs, K.J.; Scott, J.J.; Budsberg, K.J.; Read, H.; Balser, T.C.; Currie, C.R. Unique Honey Bee (Apis mellifera) Hive Component-Based Communities as Detected by a Hybrid of Phospholipid Fatty-Acid and Fatty-Acid Methyl Ester Analyses. PLoS ONE 2015, 10, e0121697. [Google Scholar] [CrossRef]
- Popa, M.; Vica, M.; Axinte, R.; Glevitzky, M.; Varvara, S. Study concerning the honey qualities in Transylvania region. Ann. Univ. Apulensis Ser. Oeconomica 2009, 11, 1034–1040. [Google Scholar]
- Nora, C.; Mahamed, A.L. Contribution to identification of the microflora of the digestive tract and pollen of Algerian honeybees: Apis mellifera intermissa and Apis mellifera sahariensis. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 601–607. [Google Scholar]
- Yoshiyama, M.; Kimura, K. Bacteria in the gut of Japanese honeybee, Apis cerana japonica, and their antagonistic effect against Paenibacillus larvae, the causal agent of American foulbrood. J. Invertebr. Pathol. 2009, 102, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Mohr, K.I.; Tebbe, C.C. Diversity and phylotype consistency of bacteria in the guts of three bee species (Apoidea) at an oilseed rape field. Environ. Microbiol. 2006, 8, 258–272. [Google Scholar] [CrossRef] [PubMed]
- Babendreier, D.; Joller, D.; Romeis, J.; Bigler, F.; Widmer, F. Bacterial community structures in honeybee intestines and their response to two insecticidal proteins. FEMS Microbiol. Ecol. 2007, 59, 600–610. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. Science (80-) 2007, 318, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Martinson, V.G.; Danforth, B.N.; Minckley, R.L.; Rueppell, O.; Tingek, S.; Moran, N.A. A simple and distinctive microbiota associated with honey bees and bumble bees. Mol. Ecol. 2011, 20, 619–628. [Google Scholar] [CrossRef]
- Disayathanoowat, T.; Young, J.P.W.; Helgason, T.; Chantawannakul, P. T-RFLP analysis of bacterial communities in the midguts of Apis mellifera and Apis cerana honey bees in Thailand. FEMS Microbiol. Ecol. 2012, 79, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.-H.; Hong, I.-P.; Bok, J.-I.; Kim, B.-Y.; Song, J.; Weon, H.-Y. Pyrosequencing analysis of the bacterial communities in the guts of honey bees Apis cerana and Apis mellifera in Korea. J. Microbiol. 2012, 50, 735–745. [Google Scholar] [CrossRef]
- Engel, P.; Martinson, V.G.; Moran, N.A. Functional diversity within the simple gut microbiota of the honey bee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, R.S.; Moran, N.A.; Evans, J.D. Early gut colonizers shape parasite susceptibility and microbiota composition in honey bee workers. Proc. Natl. Acad. Sci. USA 2016, 113, 9345–9350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabree, Z.L.; Hansen, A.K.; Moran, N.A. Independent Studies Using Deep Sequencing Resolve the Same Set of Core Bacterial Species Dominating Gut Communities of Honey Bees. PLoS ONE 2012, 7, e41250. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Saini, K.; Jain, K.L. Quantitative comparison of lipids in some pollens and their phagostimulatory effects in honey bees. J. Apic. Res. 1999, 38, 87–92. [Google Scholar] [CrossRef]
- Singh, R.; Levitt, A.L.; Rajotte, E.G.; Holmes, E.C.; Ostiguy, N.; VanEngelsdorp, D.; Lipkin, W.I.; DePamphilis, C.W.; Toth, A.L.; Cox-Foster, D.L. RNA Viruses in Hymenopteran Pollinators: Evidence of Inter-Taxa Virus Transmission via Pollen and Potential Impact on Non-Apis Hymenopteran Species. PLoS ONE 2010, 5, e14357. [Google Scholar] [CrossRef] [PubMed]
- McFrederick, Q.S.; Wcislo, W.T.; Taylor, D.R.; Ishak, H.D.; Dowd, S.E.; Mueller, U.G. Environment or kin: Whence do bees obtain acidophilic bacteria? Mol. Ecol. 2012, 21, 1754–1768. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, T.C.; Vásquez, A. Detection and Identification of a Novel Lactic Acid Bacterial Flora Within the Honey Stomach of the Honeybee Apis mellifera. Curr. Microbiol. 2008, 57, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G. The Lactobacillus and Bifidobacterium microflora of the human intestine: Composition and succession. Curr. Issues Intest. Microbiol. 2001, 2, 43–53. [Google Scholar]
- Ouwehand, A.C.; Salminen, S.; Isolauri, E. Probiotics: An overview of beneficial effects. Antonie Van Leeuwenhoek 2002, 82, 279–289. [Google Scholar] [CrossRef]
- Sazawal, S.; Hiremath, G.; Dhingra, U.; Malik, P.; Deb, S.; Black, R.E. Efficacy of probiotics in prevention of acute diarrhoea: A meta-analysis of masked, randomised, placebo-controlled trials. Lancet Infect. Dis. 2006, 6, 374–382. [Google Scholar] [CrossRef]
- Rafter, J.; Bennett, M.; Caderni, G.; Clune, Y.; Hughes, R.; Karlsson, P.C.; Klinder, A.; O’Riordan, M.; O’Sullivan, G.C.; Pool-Zobel, B.; et al. Dietary synbiotics reduce cancer risk factors in polypectomized and colon cancer patients. Am. J. Clin. Nutr. 2007, 85, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Younts-Dahl, S.M.; Galyean, M.L.; Loneragan, G.H.; Elam, N.A.; Brashears, M.M. Dietary Supplementation with Lactobacillus- and Propionibacterium-Based Direct-Fed Microbials and Prevalence of Escherichia coli O157 in Beef Feedlot Cattle and on Hides at Harvest. J. Food Prot. 2004, 67, 889–893. [Google Scholar] [CrossRef] [PubMed]
- Younts-Dahl, S.M.; Osborn, G.D.; Galyean, M.L.; Rivera, J.D.; Loneragan, G.H.; Brashears, M.M. Reduction of Escherichia coli O157 in Finishing Beef Cattle by Various Doses of Lactobacillus acidophilus in Direct-Fed Microbials. J. Food Prot. 2005, 68, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Nocek, J.E.; Kautz, W.P. Direct-Fed Microbial Supplementation on Ruminal Digestion, Health, and Performance of Pre- and Postpartum Dairy Cattle. J. Dairy Sci. 2006, 89, 260–266. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Madic, J.; Doudin, F.; Martin, C. Biotic and Abiotic Factors Influencing In Vitro Growth of Escherichia coli O157:H7 in Ruminant Digestive Contents. Appl. Environ. Microbiol. 2006, 72, 4136–4142. [Google Scholar] [CrossRef] [Green Version]
- Kačániová, M.; Gasper, J.; Brindza, J.; Schubertová, Z.; Ivanišová, E. Bacteria Of Apis Mellifera Gastrointestinal Tract: Counts, Identification And Their Antibiotic Resistance. In Agrobiodiversity for Improving Nutrition, Health and Life Quality; Agrobionet: Nitra, Slovakia, 2017; pp. 210–215. [Google Scholar]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Powell, J.E.; Guo, J.; Evans, J.D.; Wu, J.; Williams, P.; Lin, Q.; Moran, N.A.; Zhang, Z. Two gut community enterotypes recur in diverse bumblebee species. Curr. Biol. 2015, 25, R652–R653. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.M.; Wcislo, W.T.; Robinson, G.E. Developmental Transcriptome for a Facultatively Eusocial Bee, Megalopta genalis. G3 Genes Genomes Genet. 2015, 5, 2127–2135. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Genersch, E.; Forsgren, E.; Pentikäinen, J.; Ashiralieva, A.; Rauch, S.; Kilwinski, J.; Fries, I. Reclassification of Paenibacillus larvae subsp. pulvifaciens and Paenibacillus larvae subsp. larvae as Paenibacillus larvae without subspecies differentiation. Int. J. Syst. Evol. Microbiol. 2006, 56, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Genersch, E.; Ashiralieva, A.; Fries, I. Strain- and Genotype-Specific Differences in Virulence of Paenibacillus larvae subsp. larvae, a Bacterial Pathogen Causing American Foulbrood Disease in Honeybees. Appl. Environ. Microbiol. 2005, 71, 7551–7555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.D. Diverse origins of tetracycline resistance in the honey bee bacterial pathogen Paenibacillus larvae. J. Invertebr. Pathol. 2003, 83, 46–50. [Google Scholar] [CrossRef]
- Mutinelli, F.; Rademacher, E. The use of drugs to control varroosis in honey bee colonies and European legislation: The current situation. Bee World 2003, 84, 55–59. [Google Scholar] [CrossRef]
- Thompson, H.M.; Waite, R.J.; Wilkins, S.; Brown, M.A.; Bigwood, T.; Shaw, M.; Ridgway, C.; Sharman, M. Effects of European foulbrood treatment regime on oxytetracycline levels in honey extracted from treated honeybee (Apis mellifera) colonies and toxicity to brood. Food Addit. Contam. 2005, 22, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as Major Modulators of Insect Health: Lactic Acid Bacteria and Honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- D Evans, J.; Armstrong, T.-N. Inhibition of the American foulbrood bacterium, Paenibacillus larvae larvae, by bacteria isolated from honey bees. J. Apic. Res. 2005, 44, 168–171. [Google Scholar] [CrossRef]
- Evans, J.D.; Armstrong, T.-N. Antagonistic interactions between honey bee bacterial symbionts and implications for disease. BMC Ecol. 2006, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-W.; Wang, C.-H.; An, J.; Kai-Kuang, H. Susceptibility of the Asian honey bee, Apis cerana, to American foulbrood, Paenibacillus larvae larvae. J. Apic. Res. 2000, 39, 169–175. [Google Scholar] [CrossRef]
- De Vuyst, L.; Leroy, F. Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food Applications. J. Mol. Microbiol. Biotechnol. 2007, 13, 194–199. [Google Scholar] [CrossRef]
- Alippi, A.M. Is Terramycin® losing its effectiveness against AFB? Bee Biz 2000, 11, 27–29. [Google Scholar]
- Martirani, L.; Varcamonti, M.; Naclerio, G.; De Felice, M. Purification and partial characterization of bacillocin 490, a novel bacteriocin produced by a thermophilic strain of Bacillus licheniformis. Microb. Cell Fact. 2002, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Alqarni, A.; Hannan, M.; Owayss, A.; Engel, M. The indigenous honey bees of Saudi Arabia (Hymenoptera, Apidae, Apis mellifera jemenitica Ruttner): Their natural history and role in beekeeping. Zookeys 2011, 134, 83–98. [Google Scholar] [CrossRef] [Green Version]
- Alghamdi, A. The Comprehensive Study of the Mite, Varroa Destructor on Honeybees Apis mellifera; Indigenous and Imported; Bee Research Unit, PhD Department, College of Food and Agriculture Sciences, King Saud University: Riyadh, Saudi Arabia, 2002. [Google Scholar]
- Ansari, M.J.; Al-Ghamdi, A.; Usmani, S.; Al-Waili, N.; Nuru, A.; Sharma, D.; Khan, K.A.; Kaur, M.; Omer, M. In vitro evaluation of the effects of some plant essential oils on Paenibacillus larvae, the causative agent of American foulbrood. Biotechnol. Biotechnol. Equip. 2016, 30, 49–55. [Google Scholar] [CrossRef]
- Erler, S.; Moritz, R.F.A. Pharmacophagy and pharmacophory: Mechanisms of self-medication and disease prevention in the honeybee colony (Apis mellifera). Apidologie 2016, 47, 389–411. [Google Scholar] [CrossRef] [Green Version]
- Kuzyšinová, K.; Mudroňová, D.; Toporčák, J.; Molnár, L.; Javorský, P. The use of probiotics, essential oils and fatty acids in the control of American foulbrood and other bee diseases. J. Apic. Res. 2016, 55, 386–395. [Google Scholar] [CrossRef]
- Kaufman, P.; Dayanandan, P.; Li, C.; McKenzie, M.; Hoyt, J.; Kirakosyan, A. The Uses of Plant Natural Products by Humans and Risks Associated with Their Use. In Natural Products from Plants, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 441–473. [Google Scholar]
- Fuselli, S.R. Antimicrobial activity of some Argentinean wild plant essential oils against Paenibacillus larvae larvae, causal agent of American foulbrood (AFB). J. Apic. Res. 2006, 6, 2–7. [Google Scholar] [CrossRef]
- Fuselli, S.R.; de la Rosa, S.B.G.; Eguaras, M.J.; Fritz, R. Susceptibility of the Honeybee Bacterial Pathogen Paenibacillus larvae to Essential Oils Distilled from Exotic and Indigenous Argentinean Plants. J. Essent. Oil Res. 2008, 20, 464–470. [Google Scholar] [CrossRef]
- Gende, L.B.; Floris, I.; Fritz, R.; Eguaras, M.J. Antimicrobial activity of cinnamon (Cinnamomum zeylanicum) essential oil and its main components against Paenibacillus larvae from Argentine. Bull. Insectol. 2008, 61, 1–4. [Google Scholar]
- Antúnez, K.; Harriet, J.; Gende, L.; Maggi, M.; Eguaras, M.; Zunino, P. Efficacy of natural propolis extract in the control of American Foulbrood. Vet. Microbiol. 2008, 131, 324–331. [Google Scholar] [CrossRef]
- Alippi, A.M.; Reynaldi, F.J. Inhibition of the growth of Paenibacillus larvae, the causal agent of American foulbrood of honeybees, by selected strains of aerobic spore-forming bacteria isolated from apiarian sources. J. Invertebr. Pathol. 2006, 91, 141–146. [Google Scholar] [CrossRef]
- Colin, M.E.; de Lahitte, J.D.; Larribau, E.; Boué, T. Activité des huiles essentielles de Labiées sur Ascophaera apis et traitement d’un rucher. Apidologie 1989, 20, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Floris, I.; Carta, C. In vivo activity of Cinnamomum zeylanicum Nees essential oil against Bacillus larvae White. Apicoltura 1990, 57–61. [Google Scholar]
- Fuselli, S.R.; García de la Rosa, B.; Eguaras, M.J.; Fritz, R. In vitro antibacterial effect of exotic plants essential oils on the honeybee pathogen Paenibacillus larvae, causal agent of American foulbrood. Spanish J. Agric. Res. 2010, 8, 651–657. [Google Scholar] [CrossRef] [Green Version]
- González, M.J.; Marioli, J.M. Antibacterial activity of water extracts and essential oils of various aromatic plants against Paenibacillus larvae, the causative agent of American Foulbrood. J. Invertebr. Pathol. 2010, 104, 209–213. [Google Scholar] [CrossRef]
- Rovná, K.; Ivanišová, E.; Žiarovská, J.; Ferus, P.; Terentjeva, M.; Kowalczewski, P.Ł.; Kačániová, M. Characterization of Rosa canina Fruits Collected in Urban Areas of Slovakia. Genome Size, iPBS Profiles and Antioxidant and Antimicrobial Activities. Molecules 2020, 25, 1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kačániová, M.; Gasper, J.; Terentjeva, M.; Kunová, S.; Kluz, M.; Puchalski, C. Antibacterial Activity of Bees Gut Lactobacilli against Paenibacillus Larvae In Vitro. Adv. Res. Life Sci. 2018, 2, 7–10. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Bee Gut from Intestine | Bee Gut from Rectum | |||
|---|---|---|---|---|
| Winter Bees Samples | Summer Bees Samples | Winter Bees Samples | Summer Bees Samples | |
| TCAM * | 5.39 ± 0.14 a | 5.03 ± 0.16 ab | 5.00 ± 0.22 abc | 4.48 ± 0.13 abc |
| TCANM | 8.38 ± 0.11 a | 8.12 ± 0.06 b | 9.25 ± 0.15 ab | 9.05 ± 0.09 ab |
| AG+ | 6.49 ± 0.13 a | 6.13 ± 0.09 ab | 7.10 ± 0.12 abc | 6.77 ± 0.11 abc |
| BS | 2.48 ± 0.09 a | 3.43 ± 0.16 ab | 3.53 ± 0.07 ac | 3.22 ± 0.10 abc |
| LS | 7.14 ± 0.06 a | 7.66 ± 0.14 ab | 8.27 ± 0.11 ab | 8.12 ± 0.06 ab |
| PS | 2.55 ± 0.06 a | 2.29 ± 0.13 ab | 3.12 ± 0.07 abc | 2.85 ± 0.15 abc |
| ES | 3.21 ± 0.08 a | 3.42 ± 0.12 ab | 2.24 ± 0.10 abc | 2.53 ± 0.15 abc |
| SS | 3.22 ± 0.09 a | 3.45 ± 0.08 ab | 2.56 ± 0.19 abc | 2.25 ± 0.07 abc |
| CB | 2.52 ± 0.11 a | 3.25 ± 0.13 ab | 3.57 ± 0.13 abc | 3.37 ± 0.14 ac |
| Class | Genus | Species |
|---|---|---|
| Gammaproteobacteria | Aeromonas | Aeromonas salmonicida |
| Actinobacteria | Arthrobacter | Arthrobacter tumbae |
| Firmicutes | Bacillus | Bacillus cereus |
| Firmicutes | Bacillus | Bacillus circulans |
| Firmicutes | Bacillus | Bacillus licheniformis |
| Firmicutes | Bacillus | Bacillus megaterium |
| Firmicutes | Bacillus | Bacillus oleronius |
| Firmicutes | Bacillus | Bacillus spp. |
| Firmicutes | Bacillus | Bacillus subtilis |
| Firmicutes | Bacillus | Bacillus thuringiensis |
| Gammaproteobacteria | Citrobacter | Citrobacter spp. |
| Gammaproteobacteria | Citrobacter | Citrobacter braakii |
| Gammaproteobacteria | Citrobacter | Citrobacter koseri |
| Betaproteobacteria | Delftia | Delftia acidovorans |
| Gammaproteobacteria | Enterobacter | Enterobacter aerogenes |
| Gammaproteobacteria | Enterobacter | Enterobacter clocae |
| Gammaproteobacteria | Enterobacter | Enterobacter kobei |
| Firmicutes | Enterococcus | Enterococcus cloacae |
| Firmicutes | Enterococcus | Enterococcus faecalis |
| Gammaproteobacteria | Escherichia | Escherichia coli |
| Firmicutes | Fructobacillus | Fructobacillus fructosus |
| Gammaproteobacteria | Hafnia | Hafnia alvei |
| Gammaproteobacteria | Klebsiella | Klebsiella aerogenes |
| Gammaproteobacteria | Klebsiella | Klebsiella oxytoca |
| Gammaproteobacteria | Klebsiella | Klebsiella pneumoniae |
| Gammaproteobacteria | Klebsiella | Klebsiella variicola |
| Actinobacteria | Kocuria | Kocuria kristinae |
| Firmicutes | Lactobacillus | Lactobacillus acidophilus |
| Firmicutes | Lactobacillus | Lactobacillus agilis |
| Firmicutes | Lactobacillus | Lactobacillus apis |
| Firmicutes | Lactobacillus | Lactobacillus brevis |
| Firmicutes | Lactobacillus | Lactobacillus crispatus |
| Firmicutes | Lactobacillus | Lactobacillus jensenii |
| Firmicutes | Lactobacillus | Lactobacillus kunkeei |
| Firmicutes | Lactobacillus | Lactobacillus mellis |
| Firmicutes | Lactobacillus | Lactobacillus plantarum |
| Firmicutes | Lactobacillus | Lactobacillus spp. |
| Firmicutes | Lactococcus | Lactococcus garvieae |
| Firmicutes | Lactococcus | Lactococcus lactis |
| Actinobacteria | Microbacterium | Microbacterium pumilum |
| Actinobacteria | Microbacterium | Microbacterium testaceum |
| Gammaproteobacteria | Moraxella | Moraxella spp. |
| Gammaproteobacteria | Morganella | Morganella morgani |
| Firmicutes | Paenibacillus | Paenibacillus larvae |
| Gammaproteobacteria | Pantotea | Pantotea agglomerans |
| Gammaproteobacteria | Pantotea | Pantotea ananatis |
| Gammaproteobacteria | Pantotea | Pantotea vagans |
| Gammaproteobacteria | Proteus | Proteus mirabilis |
| Gammaproteobacteria | Pseudomonas | Pseudomonas marginalis |
| Gammaproteobacteria | Pseudomonas | Pseudomonas oryzihabitans |
| Gammaproteobacteria | Pseudomonas | Pseudomonas putida |
| Gammaproteobacteria | Rahnella | Rahnella aquatilis |
| Gammaproteobacteria | Rahnella | Rahnella terrigena |
| Betaproteobacteria | Ralstonia | Ralstonia picketii |
| Gammaproteobacteria | Raoultella | Raoultella ornithinolytica |
| Gammaproteobacteria | Raoultella | Raoultella planticola |
| Gammaproteobacteria | Serratia | Serratia fonticola |
| Gammaproteobacteria | Serratia | Serratia liquefaciens |
| Gammaproteobacteria | Serratia | Serratia marcescens |
| Alphaproteobacteria | Sphingomonas | Sphingomonas parapaucimobilis |
| Alphaproteobacteria | Sphingomonas | Sphingomonas melonis |
| Firmicutes | Staphylococcus | Staphylococcus capitis |
| Firmicutes | Staphylococcus | Staphylococcus epidermidis |
| Firmicutes | Staphylococcus | Staphylococcus hemolyticus |
| Firmicutes | Staphylococcus | Staphylococcus hominis |
| Firmicutes | Staphylococcus | Staphylococcus warneri |
| Species | No. of Isolates/No. of Samples | No. of Positive Samples (%) |
|---|---|---|
| Aeromonas salmonicida | 15/12 | 6.00 |
| Arthrobacter tumbae | 21/15 | 7.50 |
| Bacillus cereus | 51/25 | 12.50 |
| Bacillus circulans | 35/10 | 5.00 |
| Bacillus licheniformis | 64/25 | 12.50 |
| Bacillus megaterium | 128/96 | 48.00 |
| Bacillus oleronius | 25/20 | 10.00 |
| Bacillus spp. | 125/52 | 26.00 |
| Bacillus subtilis | 56/35 | 17.50 |
| Bacillus thuringiensis | 68/42 | 21.00 |
| Citrobacter spp. | 188/112 | 56.00 |
| Citrobacter braakii | 37/15 | 7.50 |
| Citrobacter koseri | 60/30 | 15.00 |
| Delftia acidovorans | 150/200 | 100.00 |
| Enterobacter aerogenes | 136/110 | 55.00 |
| Enterobacter clocae | 126/99 | 49.50 |
| Enterobacter kobei | 59/32 | 16.00 |
| Enterococcus cloacae | 56/15 | 7.50 |
| Enterococcus faecalis | 150/100 | 50.00 |
| Escherichia coli | 350/200 | 100.00 |
| Fructobacillus fructosus | 29/11 | 5.50 |
| Hafnia alvei | 218/169 | 84.50 |
| Klebsiella aerogenes | 59/28 | 14.00 |
| Klebsiella oxytoca | 98/58 | 29.00 |
| Klebsiella pneumoniae | 36/12 | 6.00 |
| Klebsiella variicola | 45/15 | 7.50 |
| Kocuria kristinae | 186/125 | 62.50 |
| Lactobacillus acidophilus | 64/30 | 15.00 |
| Lactobacillus agilis | 55/20 | 10.00 |
| Lactobacillus apis | 123/69 | 34.50 |
| Lactobacillus brevis | 150//100 | 50.00 |
| Lactobacillus crispatus | 164//88 | 44.00 |
| Lactobacillus jensenii | 15/10 | 5.00 |
| Lactobacillus kunkeei | 135/120 | 60.00 |
| Lactobacillus mellis | 64/35 | 17.50 |
| Lactobacillus plantarum | 95/80 | 40.00 |
| Lactobacillus spp. | 167/150 | 75.00 |
| Lactococcus garvieae | 121/90 | 45.00 |
| Lactococcus lactis | 68/39 | 19.50 |
| Microbacterium pumilum | 15/5 | 2.50 |
| Microbacterium testaceum | 25/10 | 5.00 |
| Moraxella spp. | 55/15 | 7.50 |
| Morganella morgani | 115/100 | 50.00 |
| Paenibacillus larvae | 1/1 | 0.50 |
| Pantotea agglomerans | 52/40 | 20.00 |
| Pantotea ananatis | 65/30 | 15.00 |
| Pantotea vagans | 87/58 | 29.00 |
| Proteus mirabilis | 120/95 | 47.50 |
| Pseudomonas marginalis | 12/3 | 1.50 |
| Pseudomonas oryzihabitans | 65/50 | 25.00 |
| Pseudomonas putida | 35/15 | 7.50 |
| Rahnella aquatilis | 65/40 | 20.00 |
| Rahnella terrigena | 35/22 | 11.00 |
| Ralstonia picketii | 126/110 | 55.00 |
| Raoultella ornithinolytica | 69/52 | 26.00 |
| Raoultella planticola | 35/15 | 7.50 |
| Serratia fonticola | 95/95 | 47.50 |
| Serratia liquefaciens | 87/58 | 29.00 |
| Serratia marcescens | 64/30 | 15.00 |
| Sphingomonas parapaucimobilis | 125/100 | 50.00 |
| Sphingomonas melonis | 120/60 | 30.00 |
| Staphylococcus capitis | 136/120 | 60.00 |
| Staphylococcus epidermidis | 168/62 | 31.00 |
| Staphylococcus haemolyticus | 58/35 | 17.50 |
| Staphylococcus hominis | 112/90 | 45.00 |
| Staphylococcus warneri | 64/52 | 26.00 |
| Antimicrobial | CEF | CIP | – | – |
|---|---|---|---|---|
| Resistance/Sensitivity | R/S | R/S | – | – |
| Aeromonas salmonicida | 0/15 | 0/15 | – | – |
| Arthrobacter tumbae | ND | ND | – | – |
| Bacillus cereus | ND | ND | – | – |
| Bacillus circulans | ND | ND | – | – |
| Bacillus licheniformis | ND | ND | – | – |
| Bacillus megaterium | ND | ND | – | – |
| Bacillus oleronius | ND | ND | – | – |
| Bacillus spp. | ND | ND | – | – |
| Bacillus subtilis | ND | ND | – | – |
| Bacillus thuringiensis | ND | ND | – | – |
| TIC | IMI | CIP | CHL | |
| R/S | R/S | R/S | R/S | |
| Citrobacter spp. | 8/188 | 25/188 | 0/188 | 45/188 |
| Citrobacter braakii | 6/37 | 15/37 | 5/37 | 10/37 |
| Citrobacter koseri | 16/60 | 10/60 | 5/60 | 14/60 |
| Delftia acidovorans | ND | ND | ND | ND |
| TIC | IMI | CIP | CHL | |
| R/S | R/S | R/S | R/S | |
| Enterobacter aerogenes | 61/136 | 25/136 | 10/136 | 22/136 |
| Enterobacter clocae | 28/126 | 5/136 | 1/136 | 6/136 |
| Enterobacter kobei | 9/59 | 5/59 | 0/59 | 0/59 |
| IMI | TEI | TIG | – | |
| R/S | R/S | R/S | – | |
| Enterococcus cloacae | 5/56 | 6/56 | 11/56 | – |
| Enterococcus faecalis | 58/150 | 10/150 | 25/150 | – |
| TIC | IMI | CIP | CHL | |
| R/S | R/S | R/S | R/S | |
| Escherichia coli | 53/350 | 26/350 | 12/350 | 10/350 |
| Fructobacillus fructosus | ND | ND | ND | ND |
| Hafnia alvei | 15/218 | 12/218 | 5/218 | 5/218 |
| Klebsiella aerogenes | 42/59 | 25/59 | 15/59 | 5/59 |
| Klebsiella oxytoca | 63/98 | 35/98 | 15/98 | 10/98 |
| Klebsiella pneumoniae | 14/36 | 10/36 | 5/36 | 1/36 |
| Klebsiella variicola | 5/45 | 10/45 | 4/45 | 5/45 |
| Kocuria kristinae | ND | ND | ND | ND |
| AMP | IMI | MER | CHL | |
| R/S | R/S | R/S | R/S | |
| Lactobacillus acidophilus | 4/64 | 0/64 | 0/64 | 0/64 |
| Lactobacillus agilis | 2/55 | 3/55 | 2/55 | 0/55 |
| Lactobacillus apis | 16/123 | 10/123 | 8/123 | 5/123 |
| Lactobacillus brevis | 15/150 | 20/150 | 10/150 | 15/150 |
| Lactobacillus crispatus | 25/164 | 38/164 | 5/164 | 6/164 |
| Lactobacillus jensenii | 0/15 | 0/15 | 0/15 | 0/15 |
| Lactobacillus kunkeei | 52/135 | 25/135 | 15/135 | 10/135 |
| Lactobacillus mellis | 2/64 | 0/64 | 1/64 | 0/64 |
| Lactobacillus plantarum | 50/95 | 20/95 | 10/95 | 10/95 |
| Lactobacillus spp. | 0/167 | 0/167 | 0/167 | 0/167 |
| Lactococcus garvieae | ND | ND | ND | ND |
| Lactococcus lactis | ND | ND | ND | ND |
| Microbacterium pumilum | ND | ND | ND | ND |
| Microbacterium testaceum | ND | ND | ND | ND |
| Moraxella spp. | ND | ND | ND | ND |
| TIC | IMI | CIP | CHL | |
| R/S | R/S | R/S | R/S | |
| Morganella morgani | 65/115 | 35/115 | 25/115 | 15/115 |
| Paenibacillus larvae | ND | ND | ND | ND |
| Pantotea agglomerans | 15/52 | 15/52 | 10/52 | 10/52 |
| Pantotea ananatis | 10/65 | 15/65 | 15/65 | 10/65 |
| Pantotea vagans | 37/87 | 30/87 | 15/87 | 10/87 |
| Proteus mirabilis | 25/120 | 15/120 | 16/120 | 10/120 |
| TIC | IMI | CIP | TOB | |
| R/S | R/S | R/S | R/S | |
| Pseudomonas marginalis | 5/12 | 4/12 | 2/12 | 0/12 |
| Pseudomonas oryzihabitans | 30/65 | 20/65 | 10/65 | 10/65 |
| Pseudomonas putida | 5/35 | 5/35 | 5/35 | 5/35 |
| TIC | IMI | CIP | CHL | |
| R/S | R/S | R/S | R/S | |
| Rahnella aquatilis | 24/65 | 20/65 | 12/65 | 8/65 |
| Rahnella terrigena | 5/35 | 0/35 | 0/35 | 0/35 |
| Ralstonia picketii | ND | ND | ND | ND |
| Raoultella ornithinolytica | 29/69 | 20/69 | 10/69 | 10/69 |
| Raoultella planticola | 15/35 | 20/35 | 10/35 | 5/35 |
| Serratia fonticola | 45/95 | 30/95 | 15/95 | 5/95 |
| Serratia liquefaciens | 25/87 | 32/87 | 16/87 | 10/87 |
| Serratia marcescens | 16/64 | 12/64 | 5/64 | 2/64 |
| Sphingomonas parapaucimobilis | ND | ND | ND | ND |
| Sphingomonas melonis | ND | ND | ND | ND |
| TIG | LIN | CIP | CHL | |
| R/S | R/S | R/S | R/S | |
| Staphylococcus capitis | 15/136 | 25/136 | 20/136 | 10/136 |
| Staphylococcus epidermidis | 60/168 | 30/168 | 15/168 | 5/168 |
| Staphylococcus haemolyticus | 28/58 | 15/58 | 10/58 | 5/58 |
| Staphylococcus hominis | 41/112 | 23/112 | 16/112 | 7/112 |
| Staphylococcus warneri | 5/64 | 15/64 | 10/64 | 5/64 |
| Species | P. larvae | P. larvae CCM 4483 |
|---|---|---|
| Aeromonas salmonicida | 10.67 ± 0.58 | 10.33 ± 0.58 |
| Arthrobacter tumbae | 9.67 ± 1.15 | 8.67 ± 0.58 |
| Bacillus cereus | 14.33 ± 0.58 | 13.67 ± 0.58 |
| Bacillus circulans | 14.67 ± 1.15 | 14.33 ± 0.58 |
| Bacillus licheniformis | 15.67 ± 0.58 | 16.33 ± 1.15 |
| Bacillus megaterium | 11.67 ± 0.58 | 11.33 ± 0.58 |
| Bacillus oleronius | 10.33 ± 1.15 | 10.67 ± 0.58 |
| Bacillus spp. | 9.33 ± 0.58 | 8.67 ± 0.58 |
| Bacillus subtilis | 12.33 ± 0.58 | 11.67 ± 0.58 |
| Bacillus thuringiensis | 12.33 ± 1.15 | 11.67 ± 1.15 |
| Citrobacter spp. | 8.67 ± 0.58 | 6.67 ± 1.53 |
| Citrobacter braakii | 8.33 ± 1.53 | 7.33 ± 1.15 |
| Citrobacter koseri | 6.33 ± 1.53 | 7.67 ± 0.58 |
| Delftia acidovorans | 11.67 ± 1.15 | 11.33 ± 0.58 |
| Enterobacter aerogenes | 8.67 ± 0.58 | 6.67 ± 1.53 |
| Enterobacter clocae | 8.33 ± 1.53 | 7.33 ± 1.15 |
| Enterobacter kobei | 6.33 ± 1.53 | 7.67 ± 0.58 |
| Enterococcus cloacae | 14.67 ± 0.58 | 14.33 ± 0.58 |
| Enterococcus faecalis | 16.33 ± 1.53 | 16.33 ± 0.58 |
| Escherichia coli | 15.67 ± 0.58 | 15.33 ± 0.58 |
| Fructobacillus fructosus | 18.67 ± 0.58 | 18.33 ± 0.58 |
| Hafnia alvei | 8.33 ± 1.53 | 7.33 ± 1.15 |
| Klebsiella aerogenes | 6.33 ± 1.53 | 7.67 ± 0.58 |
| Klebsiella oxytoca | 7.67 ± 0.58 | 8.33 ± 0.58 |
| Klebsiella pneumoniae | 7.33 ± 0.58 | 6.67 ± 0.58 |
| Klebsiella variicola | 5.33 ± 0.58 | 4.67 ± 0.58 |
| Kocuria kristinae | 11.33 ± 0.58 | 10.67 ± 0.58 |
| Lactobacillus acidophilus | 23.33 ± 0.58 | 22.67 ± 0.58 |
| Lactobacillus agilis | 18.67 ± 0.58 | 18.33 ± 0.58 |
| Lactobacillus apis | 20.33 ± 0.58 | 20.67 ± 0.58 |
| Lactobacillus brevis | 19.33 ± 0.58 | 19.00 ± 1.00 |
| Lactobacillus crispatus | 20.33 ± 1.15 | 19.67 ± 1.15 |
| Lactobacillus jensenii | 20.33 ± 0.58 | 20.33 ± 1.15 |
| Lactobacillus kunkeei | 25.67 ± 1.15 | 24.33 ± 0.58 |
| Lactobacillus mellis | 18.67 ± 1.15 | 17.67 ± 0.58 |
| Lactobacillus plantarum | 22.33 ± 0.58 | 21.67 ± 0.58 |
| Lactobacillus spp. | 17.00 ± 1.00 | 17.33 ± 0.58 |
| Lactococcus garvieae | 16.67 ± 0.58 | 16.33 ± 0.58 |
| Lactococcus lactis | 17.67 ± 0.58 | 17.33 ± 0.58 |
| Microbacterium pumilum | 13.67 ± 0.58 | 13.33 ± 0.58 |
| Microbacterium testaceum | 12.67 ± 0.58 | 12.33 ± 0.58 |
| Moraxella spp. | 8.67 ± 0.58 | 6.67 ± 1.53 |
| Morganella morgani | 8.33 ± 1.53 | 7.33 ± 1.15 |
| Pantotea agglomerans | 6.33 ± 1.53 | 7.67 ± 0.58 |
| Pantotea ananatis | 8.67 ± 0.58 | 6.67 ± 1.53 |
| Proteus mirabilis | 8.33 ± 1.53 | 7.33 ± 1.15 |
| Pantotea vagans | 6.33 ± 1.53 | 7.67 ± 0.58 |
| Pseudomonas marginalis | 11.33 ± 0.58 | 10.67 ± 0.58 |
| Pseudomonas oryzihabitans | 11.33 ± 1.15 | 11.00 ± 1.00 |
| Pseudomonas putida | 10.67 ± 0.58 | 10.33 ± 0.58 |
| Rahnella aquatilis | 8.67 ± 0.58 | 6.67 ± 1.53 |
| Rahnella terrigena | 8.33 ± 1.53 | 7.33 ± 1.15 |
| Ralstonia picketii | 6.33 ± 1.53 | 7.67 ± 0.58 |
| Raoultella ornithinolytica | 8.67 ± 0.58 | 6.67 ± 1.53 |
| Raoultella planticola | 8.33 ± 1.53 | 7.33 ± 1.15 |
| Serratia fonticola | 8.67 ± 0.58 | 6.67 ± 1.53 |
| Serratia liquefaciens | 8.33 ± 1.53 | 7.33 ± 1.15 |
| Serratia marcescens | 6.33 ± 1.53 | 7.67 ± 0.58 |
| Sphingomonas parapaucimobilis | 11.67 ± 1.15 | 11.33 ± 0.58 |
| Sphingomonas melonis | 10.67 ± 0.58 | 10.33 ± 0.58 |
| Staphylococcus capitis | 13.67 ± 0.58 | 13.33 ± 0.58 |
| Staphylococcus epidermidis | 14.67 ± 0.58 | 14.33 ± 0.58 |
| Staphylococcus haemolyticus | 13.67 ± 0.58 | 13.33 ± 0,58 |
| Staphylococcus hominis | 12.67 ± 0.58 | 12.33 ± 0.58 |
| Staphylococcus warneri | 11.67 ± 0.58 | 11.33 ± 0.58 |
| Essential Oil | P. larvae | P. larvae CCM 4483 |
|---|---|---|
| Lavandula angustifolia Mill. | 14.33 ± 1.15 | 15.33 ± 0.58 |
| Cinnamomum zeylanicum L. | 10.00 ± 1.00 | 12.33 ± 2.52 |
| Pinus montana Mill. | 17.67 ± 0.58 | 17.33 ± 0.58 |
| Mentha piperita L. | 7.33 ± 0.58 | 7.00 ± 2.00 |
| Foeniculum vulgare Mill. | 14.66 ± 0.58 | 14.00 ± 0.57 |
| Pinus sylvestris L. | 17.00 ± 1.00 | 17.67 ± 0.57 |
| Satureja hortensis L. | 12.33 ± 0.58 | 17.67 ± 1.53 |
| Origanum vulgare L. | 18.67 ± 1.15 | 19.00 ± 1.00 |
| Pimpinella anisum L. | 12.33 ± 0.58 | 11.67 ± 0.58 |
| Rosmarinus officinalis L. | 14.67 ± 0.58 | 10.00 ± 1.00 |
| Salvia officinalis L. | 14.33 ± 0.58 | 13.00 ± 1.00 |
| Abies alba Mill. | 17.33 ± 0.58 | 18.00 ± 1.00 |
| Citrus aurantium var. dulce L. | 4.33 ± 0.58 | 3.00 ± 1.00 |
| Citrus sinensis L. Osbeck. | 2.00 ± 1.00 | 5.33 ± 0.58 |
| Cymbopogon nardus L. | 8.67 ± 0.58 | 8.00 ± 1.00 |
| Mentha spicata var. crispa L. | 9.67 ± 1.53 | 9.33 ± 0.57 |
| Thymus vulgaris L. | 19.67 ± 1.53 | 15.67 ± 1.53 |
| Carvum carvi L. | 7.67 ± 0.58 | 5.00 ± 0.58 |
| Thymus serpyllum L. | 4.33 ± 0.58 | 7.33 ± 0.58 |
| Amyris balsamifera | 9.33 ± 0.58 | 9.67 ± 0.58 |
| Ocimum basilicum | 13.67 ± 1.15 | 14.00 ± 1.00 |
| Canarium luzonicum Miq. | 11.33 ± 1.15 | 12.33 ± 0.58 |
| Eucalyptus globulus | 16.33 ± 1.15 | 17.33 ± 0.58 |
| Gaultheria procumbens | 8.33 ± 0.58 | 7.33 ± 0.58 |
| Pelargonium graveolens | 6.67 ± 0.58 | 7.33 ± 0.58 |
| Cinnamomum caphora var. linalolifera | 16.00 ± 1.73 | 15.67 ± 1.15 |
| Boswellia carterii | 7.67 ± 1.15 | 7.00 ± 1.00 |
| Melaleuca leucadendron | 9.67 ± 0.58 | 9.33 ± 0.58 |
| Litsea cubeba Pers. | 10.33 ± 0.58 | 10.66 ± 0.58 |
| Melaleuca ericifolia Smith. | 9.67 ± 0.58 | 10.00 ± 1.00 |
| Microorganisms Group | Dilution | Agar | Inoculation Method | Cultivation Condition | ||
|---|---|---|---|---|---|---|
| Relation of O2 | Temperature | Time | ||||
| TCAM | 10−5–10−7 | PCA | surface | aerobic | 30 °C | 48 h |
| TCANM | 10−5–10−7 | PCA | surface | anaerobic | 25 °C | 48 h |
| AG+ | 10−3–10−6 | AA | surface | anaerobic | 37 °C | 48 h |
| Bacillus spp. | 10−3–10−5 | PCA | surface | aerobic | 30 °C | 48 h |
| Lactobacillus spp. | 10−2–10−6 | MRS | surface | aerobic | 37 °C | 48 h |
| Pseudomonas spp. | 10−3–10−5 | Pseudomonas agar | surface | aerobic | 30 °C | 48 h |
| Enterococcus spp. | 10−3–10−5 | Enterococcus selective agar | surface | aerobic | 37 °C | 48 h |
| Staphylococcus spp. | 10−2–10−4 | Blood agar | surface | aerobic | 37 °C | 48 h |
| CB | 10−4–10−6 | McC | surface | aerobic | 37 °C | 48 h |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kačániová, M.; Terentjeva, M.; Žiarovská, J.; Kowalczewski, P.Ł. In Vitro Antagonistic Effect of Gut Bacteriota Isolated from Indigenous Honey Bees and Essential Oils against Paenibacillus Larvae. Int. J. Mol. Sci. 2020, 21, 6736. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186736
Kačániová M, Terentjeva M, Žiarovská J, Kowalczewski PŁ. In Vitro Antagonistic Effect of Gut Bacteriota Isolated from Indigenous Honey Bees and Essential Oils against Paenibacillus Larvae. International Journal of Molecular Sciences. 2020; 21(18):6736. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186736
Chicago/Turabian StyleKačániová, Miroslava, Margarita Terentjeva, Jana Žiarovská, and Przemysław Łukasz Kowalczewski. 2020. "In Vitro Antagonistic Effect of Gut Bacteriota Isolated from Indigenous Honey Bees and Essential Oils against Paenibacillus Larvae" International Journal of Molecular Sciences 21, no. 18: 6736. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186736