Cooperativity between the Ribosome-Associated Chaperone Ssb/RAC and the Ubiquitin Ligase Ltn1 in Ubiquitination of Nascent Polypeptides


Abstract
:1. Introduction
2. Results
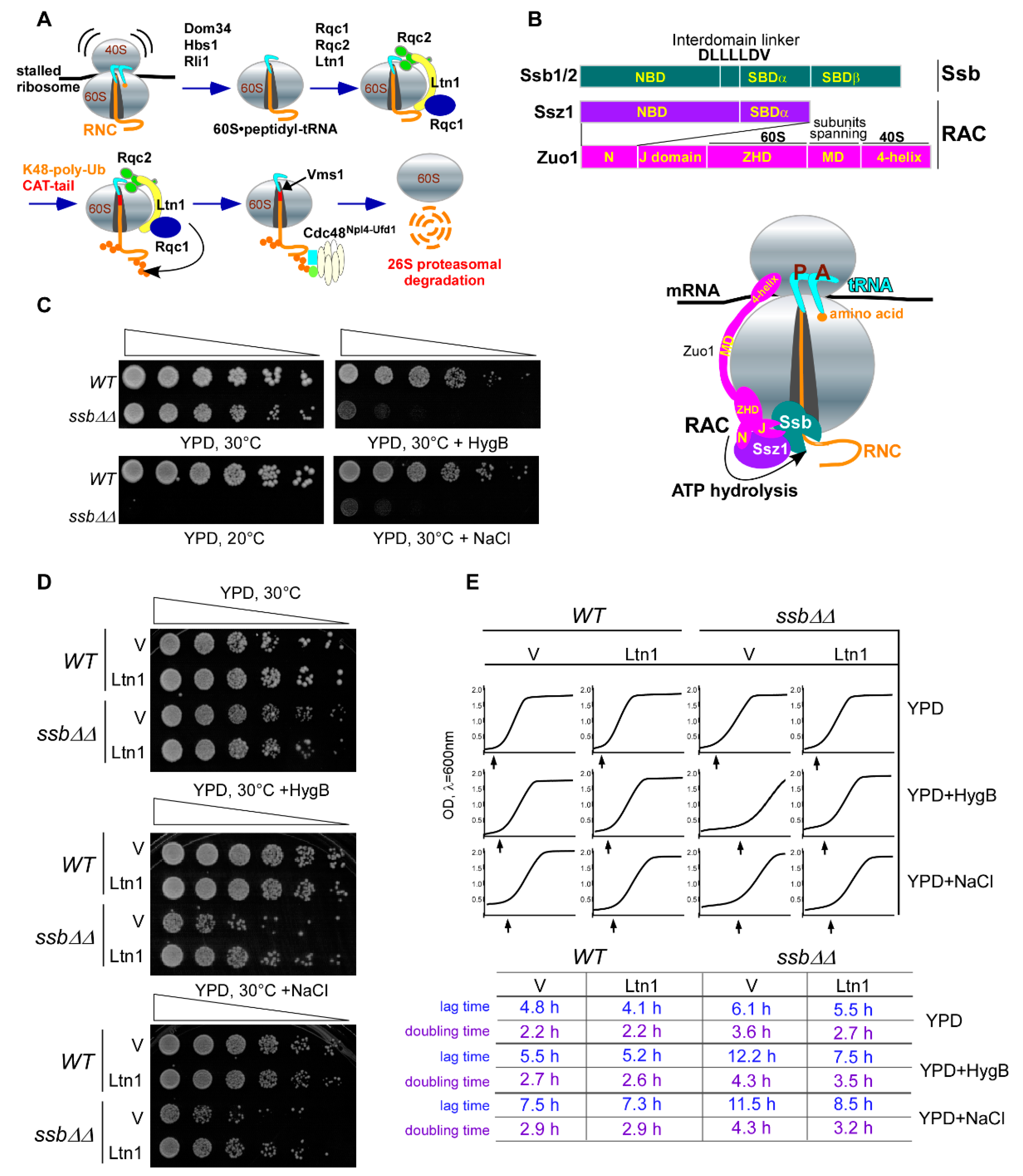
2.1. Ubiquitin Ligase Ltn1 and Ribosome-Associated Chaperone Ssb Demonstrate Genetic Interaction
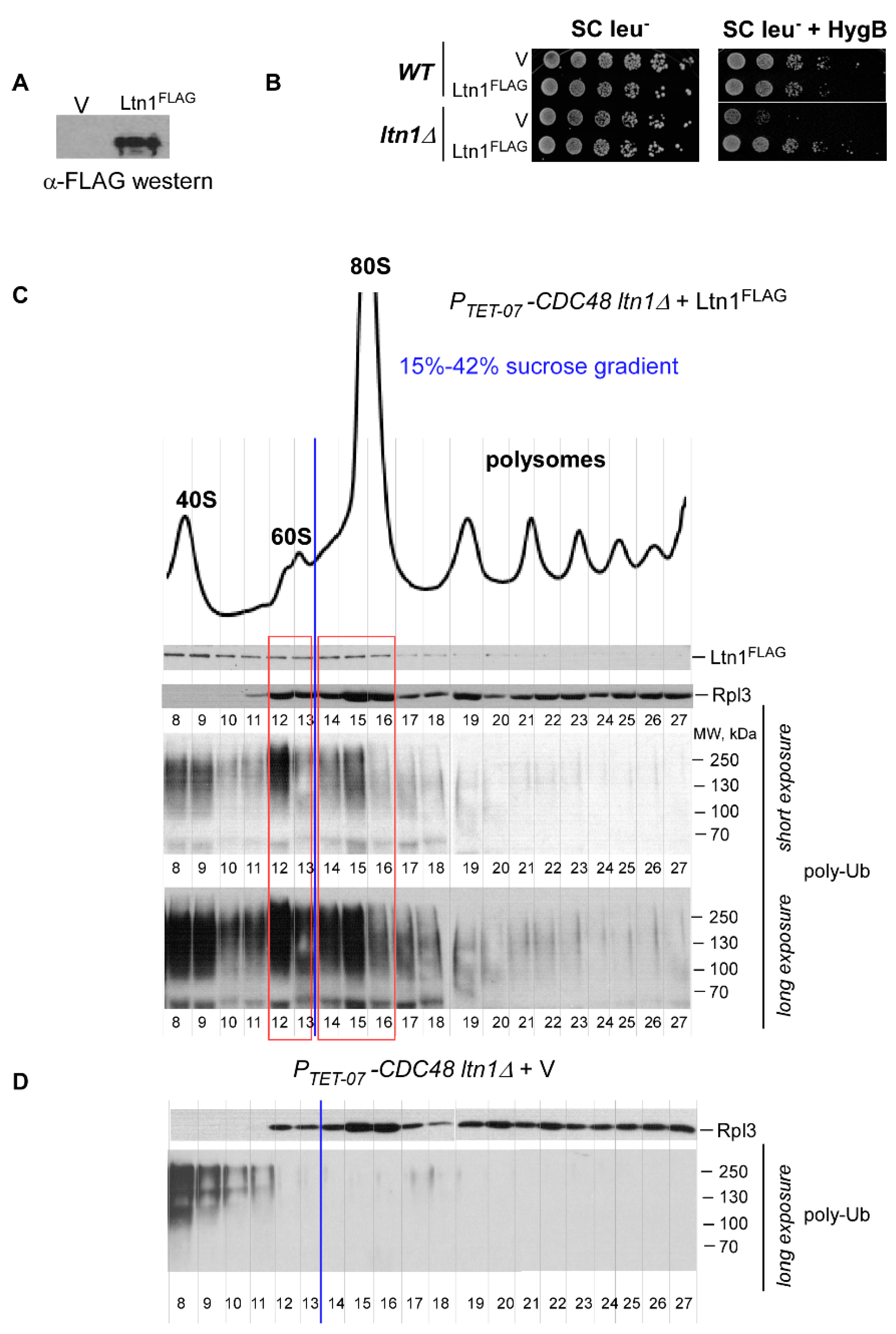
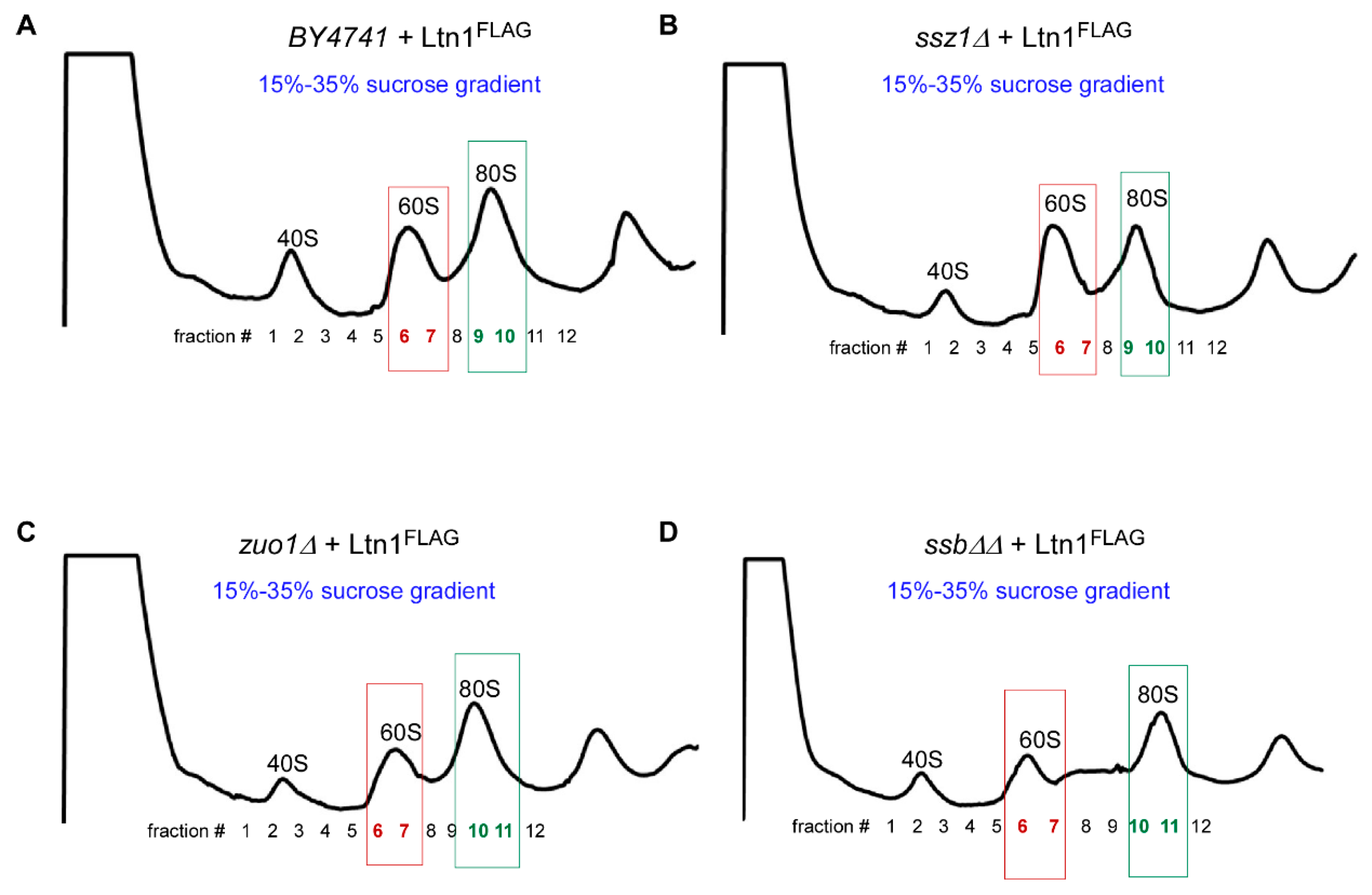
2.2. Ltn1FLAG Associates with Both 60S and 80S, Promoting Polyubiquitination of Associated RNCs
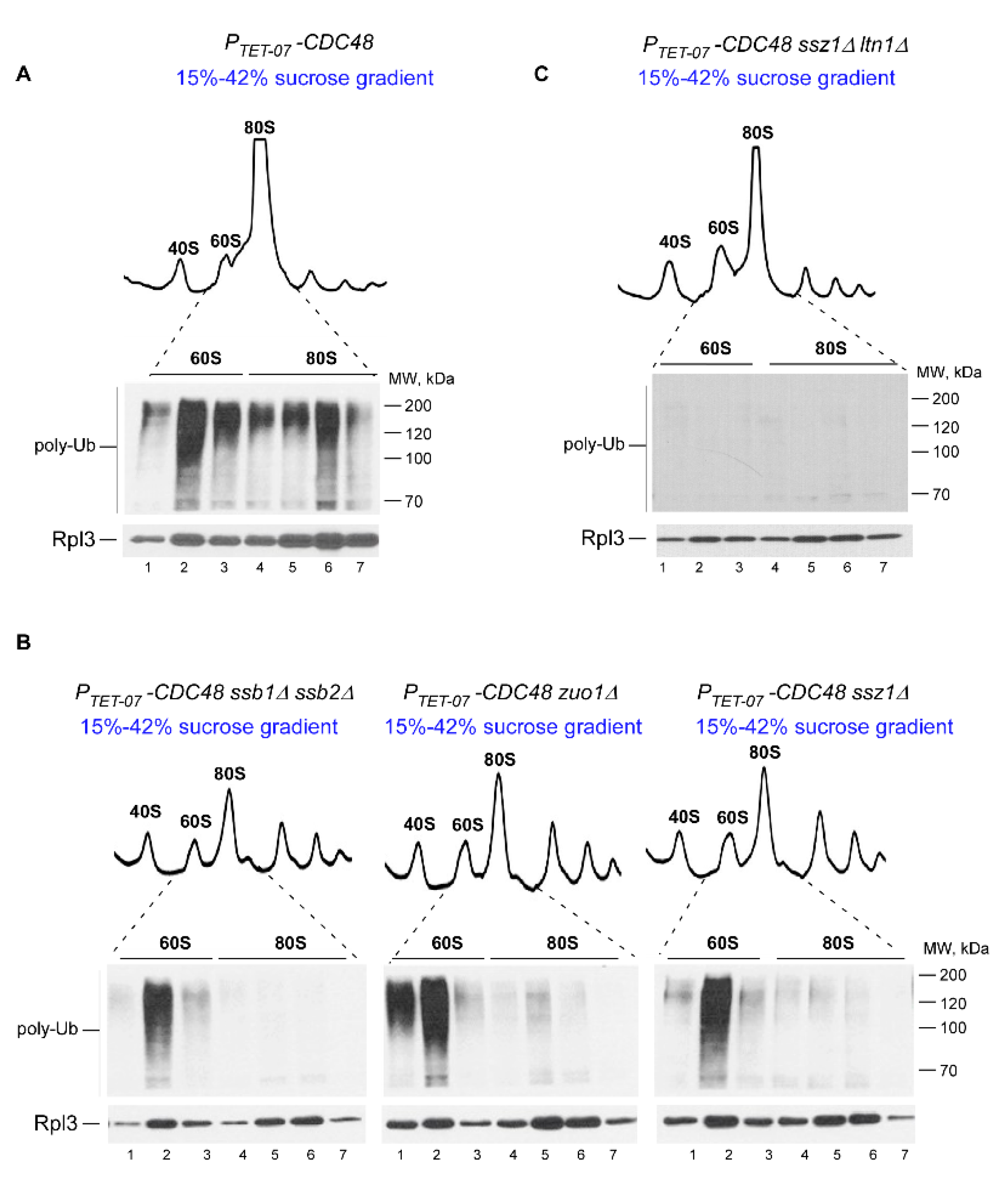
2.3. Ssb/RAC Promotes Ubiquitination of Endogenous RNCs on 80S Species
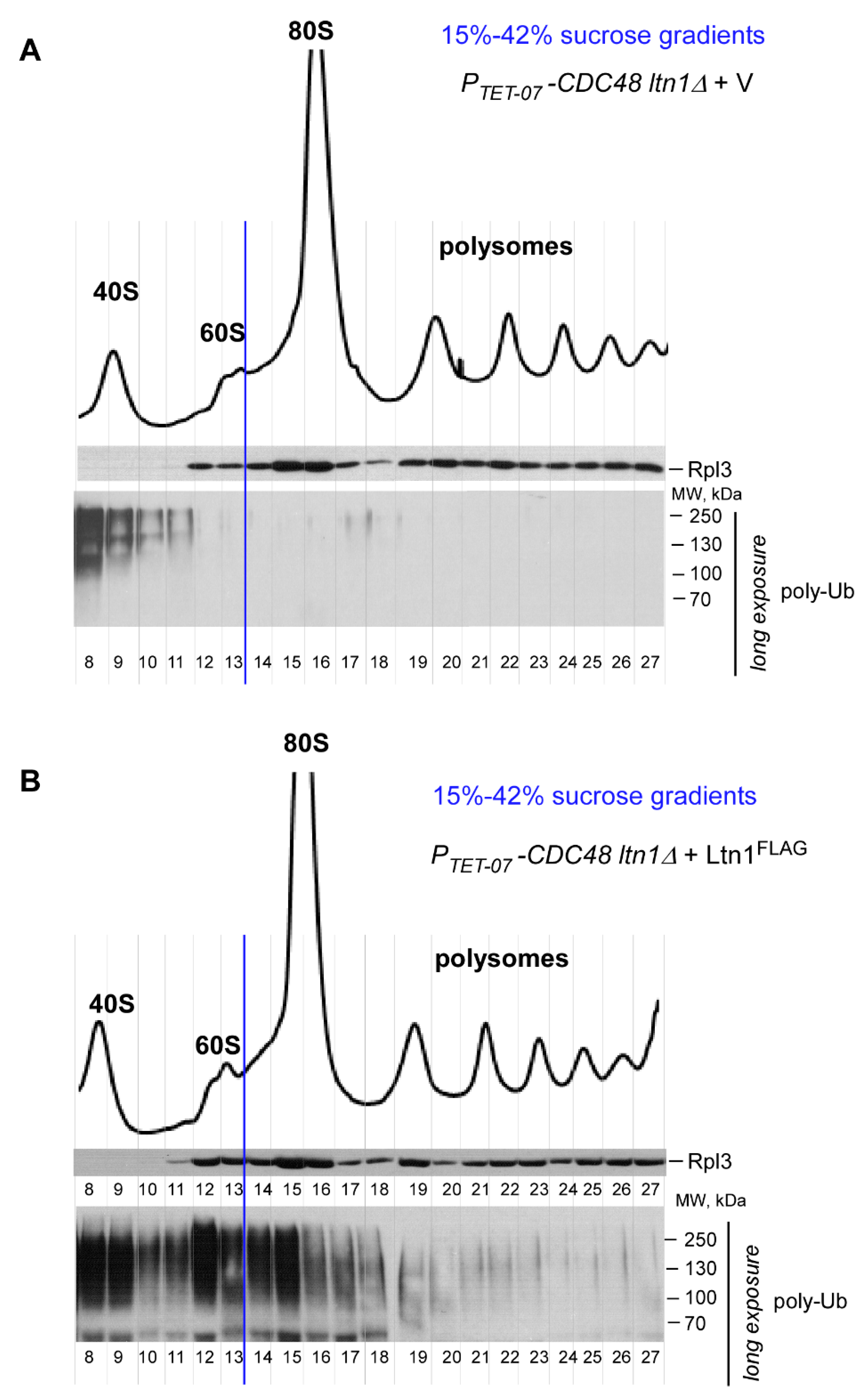
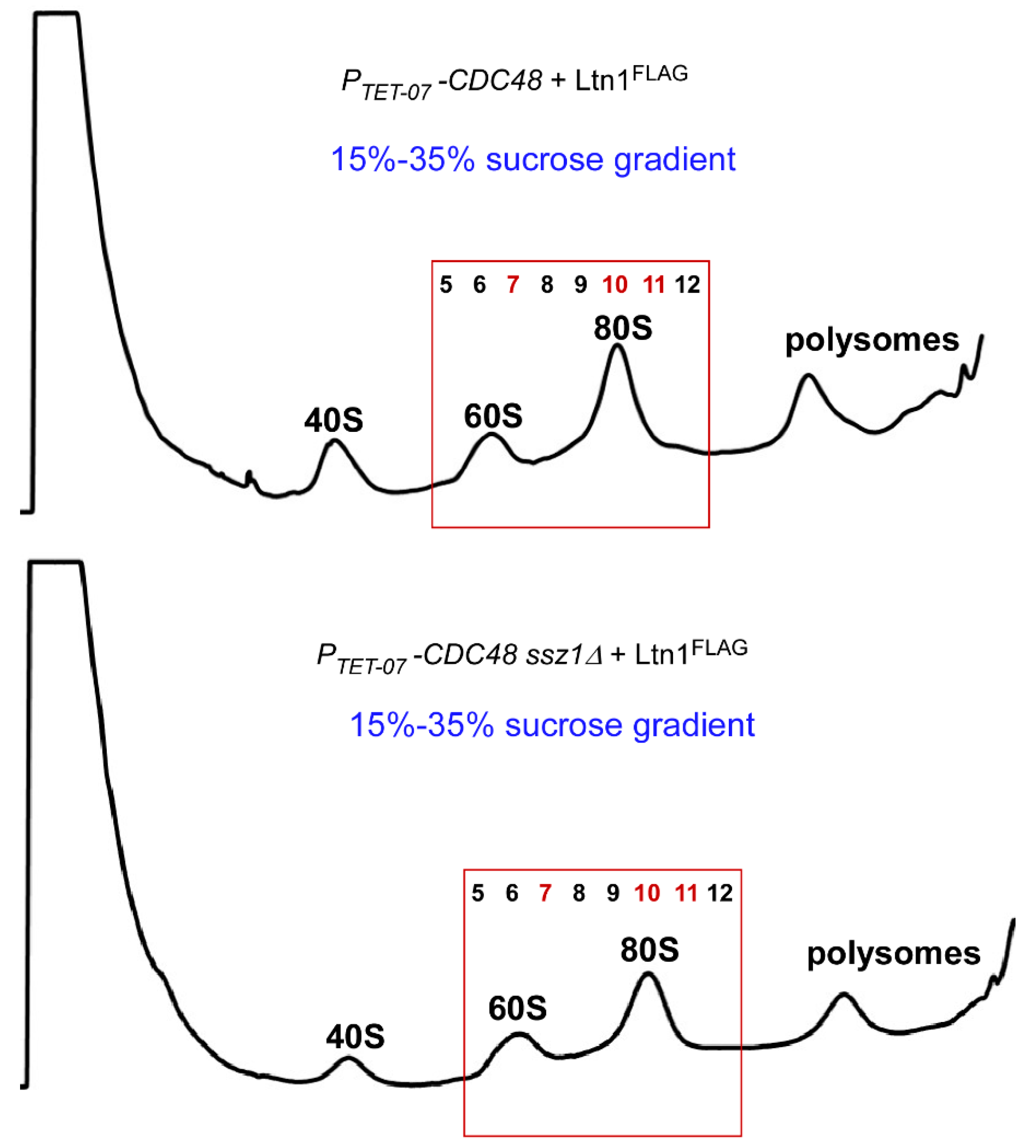
2.4. Disrupting the Ssb/RAC Triad Abrogates Ltn1-Dependent Ubiquitination of 80S-Associated RNCs upon Depletion of Cdc48
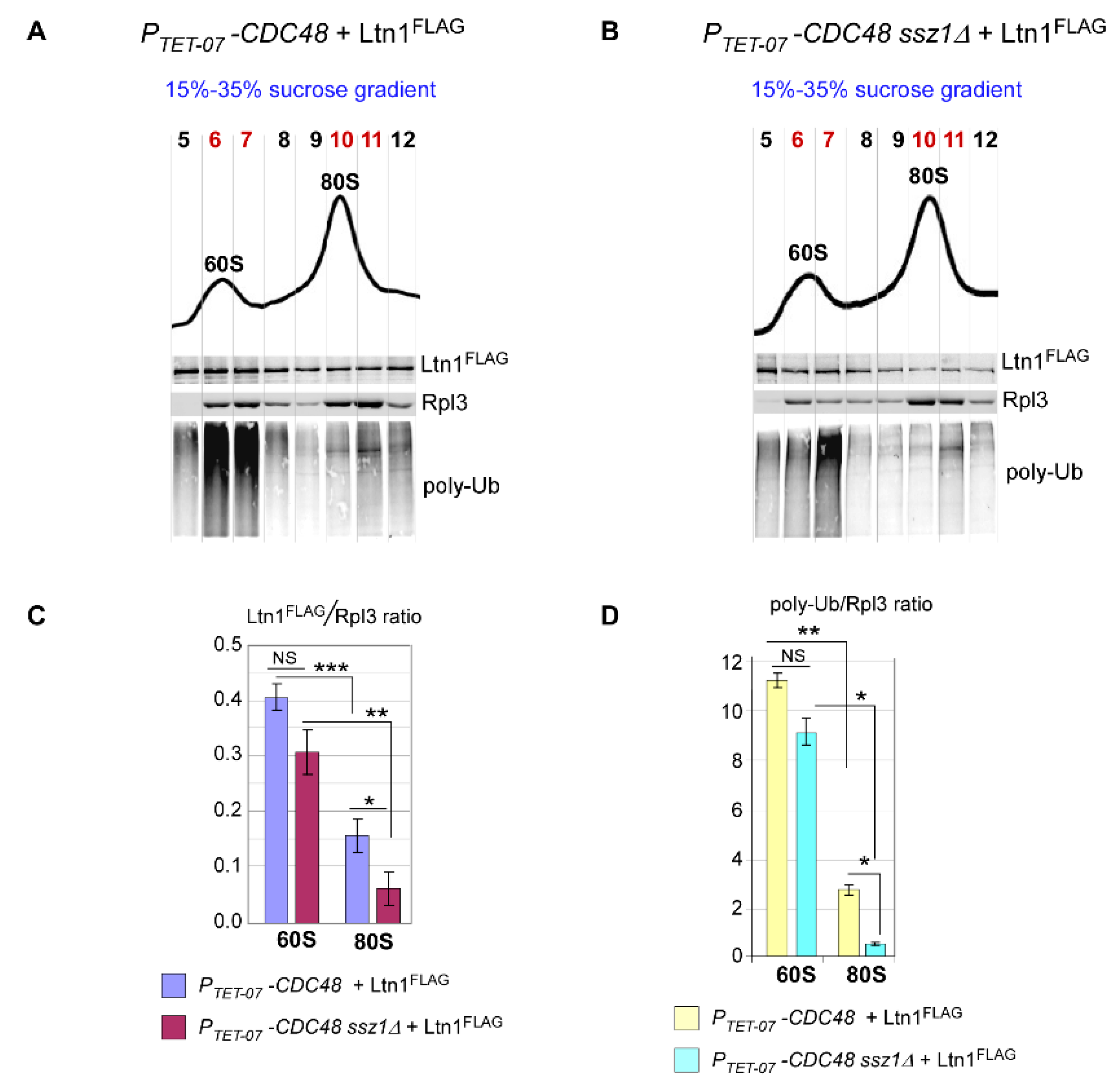
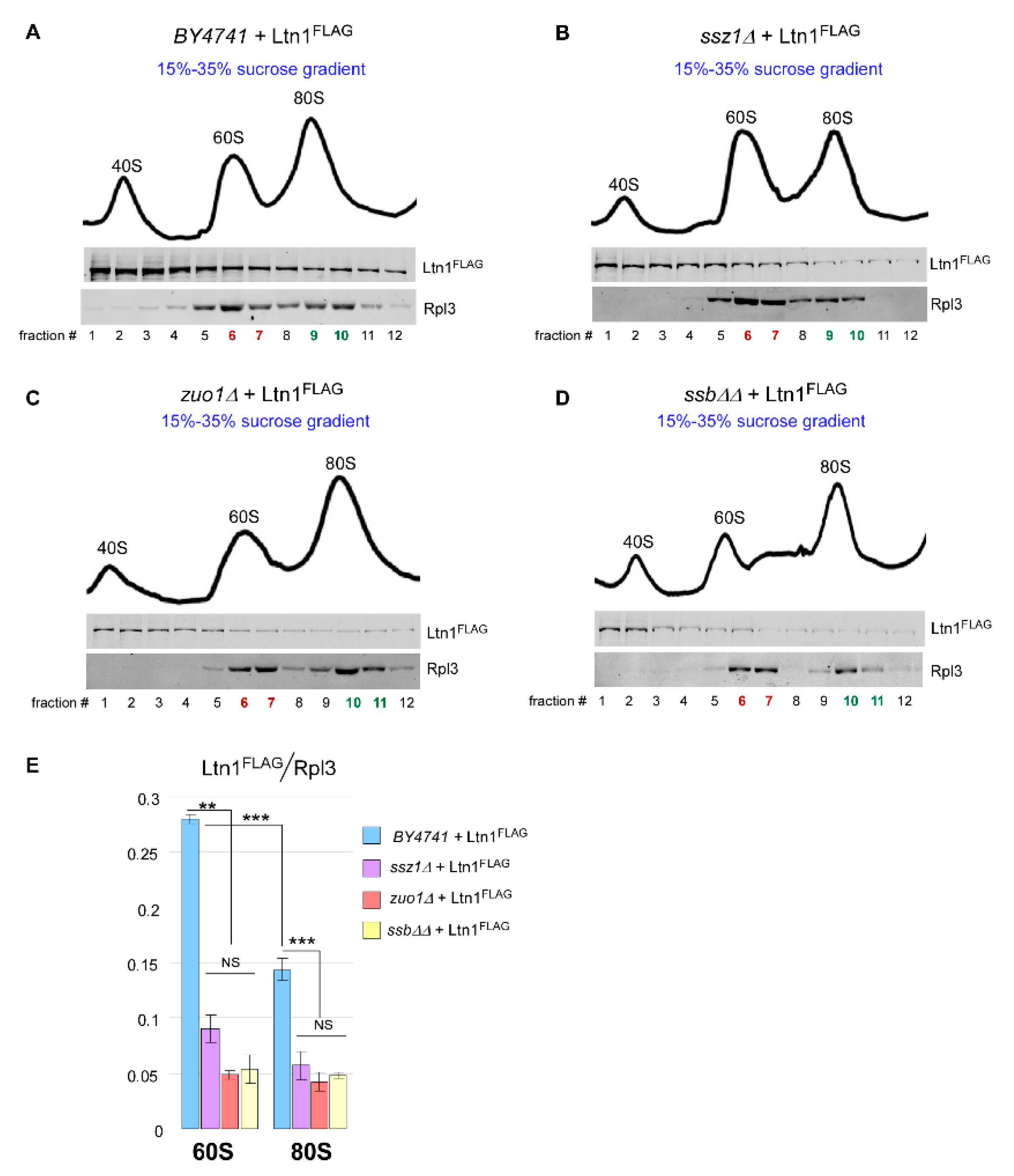
2.5. Loss of Ssb/RAC Triad Integrity Decreases Ltn1 Association with Both 60S and 80S in Wild-Type Cells
3. Discussion
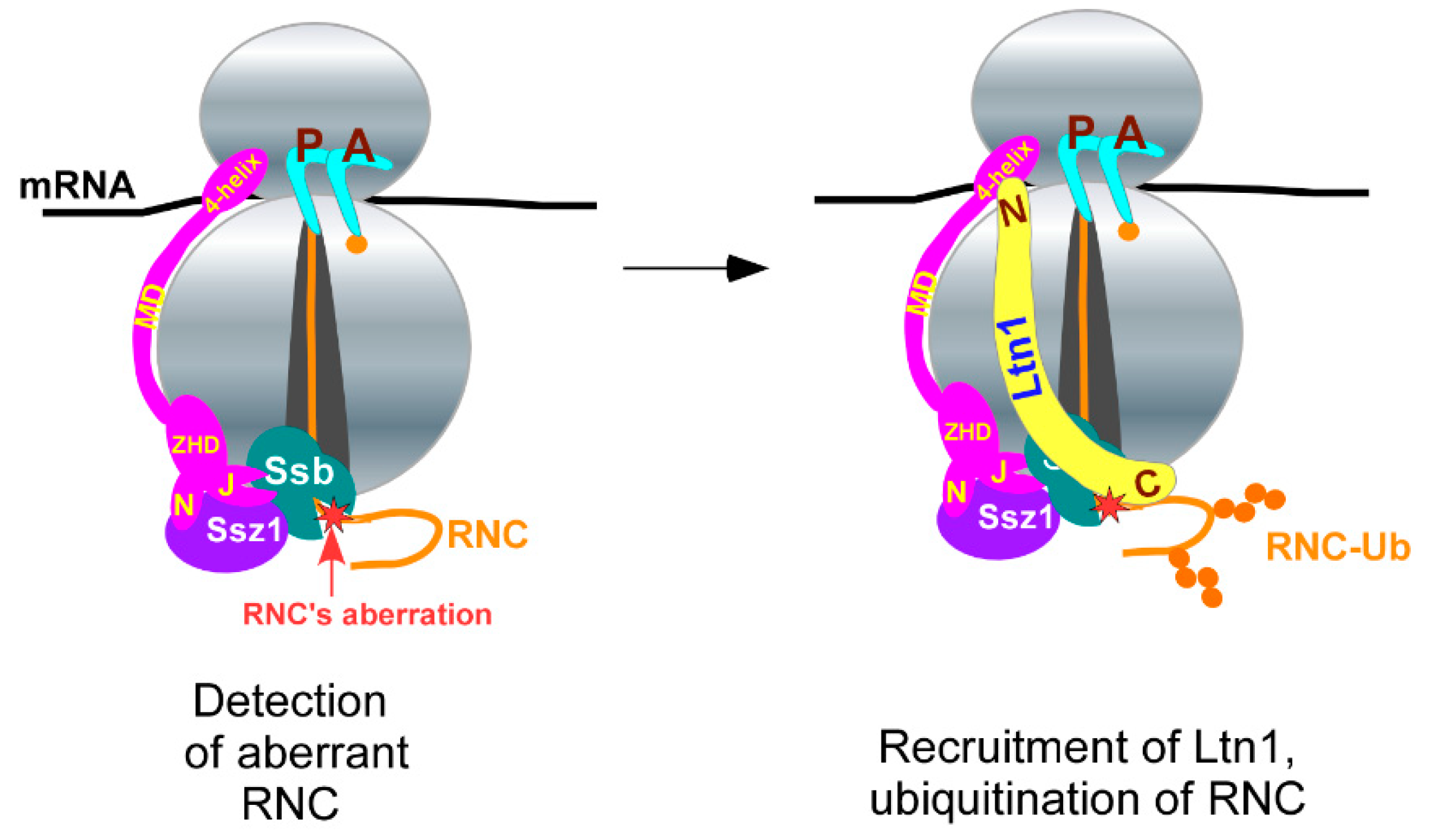
3.1. Non-Conventional 80S-RQC Pathway
3.2. Hsp70-Ssb and Ltn1 Binding to Ribosomes
3.3. Ssb/RAC Activity Is Associated with Free 60S Subunit
4. Materials and Methods
4.1. Antibodies and Chemicals
4.2. Plasmids
4.3. Yeast Strains, Medium, and Growth Conditions
4.4. Cell Viability and Growth Assays
4.5. Sucrose Gradient Centrifugation
4.6. Protein Extraction and Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RAC | Ribosome-associated complex |
| RNC | Ribosome-bound nascent chain |
| RQC | Ribosome-associated protein quality control |
Appendix A



References
- Ciechanover, A.; Kwon, Y.T. Protein Quality Control by Molecular Chaperones in Neurodegeneration. Front. Neurosci. 2017, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, S.-O. Guiding tail-anchored membrane proteins to the endoplasmic reticulum in a chaperone cascade. J. Biol. Chem. 2019, 294, 16577–16586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joazeiro, C.A.P. Mechanisms and functions of ribosome-associated protein quality control. Nat. Rev. Mol. Cell Biol. 2019, 20, 368–383. [Google Scholar] [CrossRef] [PubMed]
- Bengtson, M.H.; Joazeiro, C.A.P. Role of a ribosome-associated E3 ubiquitin ligase in protein quality control. Nature 2010, 467, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Brandman, O.; Stewart-Ornstein, J.; Wong, D.; Larson, A.; Williams, C.C.; Li, G.-W.; Zhou, S.; King, D.; Shen, P.S.; Weibezahn, J.; et al. A ribosome-bound quality control complex triggers degradation of nascent peptides and signals translation stress. Cell 2012, 151, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Defenouillère, Q.; Yao, Y.; Mouaikel, J.; Namane, A.; Galopier, A.; Decourty, L.; Doyen, A.; Malabat, C.; Saveanu, C.; Jacquier, A.; et al. Cdc48-associated complex bound to 60S particles is required for the clearance of aberrant translation products. Proc. Natl. Acad. Sci. USA 2013, 110, 5046–5051. [Google Scholar] [CrossRef] [Green Version]
- Pisareva, V.P.; Skabkin, M.A.; Hellen, C.U.T.; Pestova, T.V.; Pisarev, A.V. Dissociation by Pelota, Hbs1 and ABCE1 of mammalian vacant 80S ribosomes and stalled elongation complexes. EMBO J. 2011, 30, 1804–1817. [Google Scholar] [CrossRef] [Green Version]
- Shoemaker, C.J.; Eyler, D.E.; Green, R. Dom34:Hbs1 promotes subunit dissociation and peptidyl-tRNA drop-off to initiate no-go decay. Science 2010, 330, 369–372. [Google Scholar] [CrossRef] [Green Version]
- Tsuboi, T.; Kuroha, K.; Kudo, K.; Makino, S.; Inoue, E.; Kashima, I.; Inada, T. Dom34:hbs1 plays a general role in quality-control systems by dissociation of a stalled ribosome at the 3′ end of aberrant mRNA. Mol. Cell 2012, 46, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Kuroha, K.; Zinoviev, A.; Hellen, C.U.T.; Pestova, T.V. Release of Ubiquitinated and Non-ubiquitinated Nascent Chains from Stalled Mammalian Ribosomal Complexes by ANKZF1 and Ptrh1. Mol. Cell 2018, 72, 286–302.e8. [Google Scholar] [CrossRef] [Green Version]
- Shen, P.S.; Park, J.; Qin, Y.; Li, X.; Parsawar, K.; Larson, M.H.; Cox, J.; Cheng, Y.; Lambowitz, A.M.; Weissman, J.S.; et al. Rqc2p and 60S ribosomal subunits mediate mRNA-independent elongation of nascent chains. Science 2015, 347, 75–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, R.; Reichermeier, K.M.; Burroughs, A.M.; Oania, R.S.; Reitsma, J.M.; Aravind, L.; Deshaies, R.J. Vms1 and ANKZF1 peptidyl-tRNA hydrolases release nascent chains from stalled ribosomes. Nature 2018, 557, 446–451. [Google Scholar] [CrossRef]
- Verma, R.; Oania, R.S.; Kolawa, N.J.; Deshaies, R.J. Cdc48/p97 promotes degradation of aberrant nascent polypeptides bound to the ribosome. eLife 2013, 2, e00308. [Google Scholar] [CrossRef] [PubMed]
- Shorter, J.; Houry, W.A. Editorial: The Role of AAA+ Proteins in Protein Repair and Degradation. Front. Mol. Biosci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Jentsch, S.; Rumpf, S. Cdc48 (p97): A “molecular gearbox” in the ubiquitin pathway? Trends Biochem. Sci. 2007, 32, 6–11. [Google Scholar] [CrossRef]
- Meyer, H.; Bug, M.; Bremer, S. Emerging functions of the VCP/p97 AAA-ATPase in the ubiquitin system. Nat. Cell Biol. 2012, 14, 117–123. [Google Scholar] [CrossRef]
- Ye, Y. Diverse functions with a common regulator: Ubiquitin takes command of an AAA ATPase. J. Struct. Biol. 2006, 156, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, Y.-J.; Park, S.-H.; Hassemer, T.; Körner, R.; Vincenz-Donnelly, L.; Hayer-Hartl, M.; Hartl, F.U. Failure of RQC machinery causes protein aggregation and proteotoxic stress. Nature 2016, 531, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Defenouillère, Q.; Zhang, E.; Namane, A.; Mouaikel, J.; Jacquier, A.; Fromont-Racine, M. Rqc1 and Ltn1 Prevent C-terminal Alanine-Threonine Tail (CAT-tail)-induced Protein Aggregation by Efficient Recruitment of Cdc48 on Stalled 60S Subunits. J. Biol. Chem. 2016, 291, 12245–12253. [Google Scholar] [CrossRef] [Green Version]
- Klaips, C.L.; Jayaraj, G.G.; Hartl, F.U. Pathways of cellular proteostasis in aging and disease. J. Cell Biol. 2018, 217, 51–63. [Google Scholar] [CrossRef]
- Deuerling, E.; Gamerdinger, M.; Kreft, S.G. Chaperone Interactions at the Ribosome. Cold Spring Harb. Perspect. Biol. 2019, 11, a033977. [Google Scholar] [CrossRef] [PubMed]
- Gamerdinger, M. Protein quality control at the ribosome: Focus on RAC, NAC and RQC. Essays Biochem. 2016, 60, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frydman, J. Folding of Newly Translated Proteins In Vivo: The Role of Molecular Chaperones. Annu. Rev. Biochem. 2001, 70, 603–647. [Google Scholar] [CrossRef] [PubMed]
- Preissler, S.; Deuerling, E. Ribosome-associated chaperones as key players in proteostasis. Trends Biochem. Sci. 2012, 37, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Sinning, I.; Rospert, S. Two chaperones locked in an embrace: Structure and function of the ribosome-associated complex RAC. Nat. Struct. Mol. Biol. 2017, 24, 611–619. [Google Scholar] [CrossRef]
- Gautschi, M.; Lilie, H.; Fünfschilling, U.; Mun, A.; Ross, S.; Lithgow, T.; Rücknagel, P.; Rospert, S. RAC, a stable ribosome-associated complex in yeast formed by the DnaK-DnaJ homologs Ssz1p and zuotin. Proc. Natl. Acad. Sci. USA 2001, 98, 3762–3767. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; Gautschi, M.; Walter, W.; Rospert, S.; Craig, E.A. The Hsp70 Ssz1 modulates the function of the ribosome-associated J-protein Zuo1. Nat. Struct. Mol. Biol. 2005, 12, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Leidig, C.; Bange, G.; Kopp, J.; Amlacher, S.; Aravind, A.; Wickles, S.; Witte, G.; Hurt, E.; Beckmann, R.; Sinning, I. Structural characterization of a eukaryotic chaperone—The ribosome-associated complex. Nat. Struct. Mol. Biol. 2013, 20, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Sharma, R.; Shrestha, O.K.; Bingman, C.A.; Craig, E.A. Dual interaction of the Hsp70 J-protein cochaperone Zuotin with the 40S and 60S ribosomal subunits. Nat. Struct. Mol. Biol. 2016, 23, 1003–1010. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Susanto, T.T.; Saurabh, S.; Barna, M. Decoding the Function of Expansion Segments in Ribosomes. Mol. Cell 2018, 72, 1013–1020.e6. [Google Scholar] [CrossRef] [Green Version]
- Peisker, K.; Braun, D.; Wölfle, T.; Hentschel, J.; Fünfschilling, U.; Fischer, G.; Sickmann, A.; Rospert, S. Ribosome-associated complex binds to ribosomes in close proximity of Rpl31 at the exit of the polypeptide tunnel in yeast. Mol. Biol. Cell 2008, 19, 5279–5288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan-Minogue, H.; Bedwell, D.M. Eukaryotic ribosomal RNA determinants of aminoglycoside resistance and their role in translational fidelity. RNA 2008, 14, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiaux, J.; Horst, J.; Scior, A.; Preissler, S.; Koplin, A.; Bukau, B.; Deuerling, E. Structural analysis of the ribosome-associated complex (RAC) reveals an unusual Hsp70/Hsp40 interaction. J. Biol. Chem. 2010, 285, 3227–3234. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ma, C.; Yuan, Y.; Zhu, J.; Li, N.; Chen, C.; Wu, S.; Yu, L.; Lei, J.; Gao, N. Structural basis for interaction of a cotranslational chaperone with the eukaryotic ribosome. Nat. Struct. Mol. Biol. 2014, 21, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Willmund, F.; del Alamo, M.; Pechmann, S.; Chen, T.; Albanèse, V.; Dammer, E.B.; Peng, J.; Frydman, J. The cotranslational function of ribosome-associated Hsp70 in eukaryotic protein homeostasis. Cell 2013, 152, 196–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautschi, M.; Mun, A.; Ross, S.; Rospert, S. A functional chaperone triad on the yeast ribosome. Proc. Natl. Acad. Sci. USA 2002, 99, 4209–4214. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Hanebuth, M.A.; Kityk, R.; Fries, S.J.; Jain, A.; Kriel, A.; Albanese, V.; Frickey, T.; Peter, C.; Mayer, M.P.; Frydman, J.; et al. Multivalent contacts of the Hsp70 Ssb contribute to its architecture on ribosomes and nascent chain interaction. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Raue, U.; Oellerer, S.; Rospert, S. Association of Protein Biogenesis Factors at the Yeast Ribosomal Tunnel Exit Is Affected by the Translational Status and Nascent Polypeptide Sequence. J. Biol. Chem. 2007, 282, 7809–7816. [Google Scholar] [CrossRef] [Green Version]
- Döring, K.; Ahmed, N.; Riemer, T.; Suresh, H.G.; Vainshtein, Y.; Habich, M.; Riemer, J.; Mayer, M.P.; O’Brien, E.P.; Kramer, G.; et al. Profiling Ssb-Nascent Chain Interactions Reveals Principles of Hsp70-Assisted Folding. Cell 2017, 170, 298–311.e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gribling-Burrer, A.-S.; Chiabudini, M.; Zhang, Y.; Qiu, Z.; Scazzari, M.; Wölfle, T.; Wohlwend, D.; Rospert, S. A dual role of the ribosome-bound chaperones RAC/Ssb in maintaining the fidelity of translation termination. Nucleic Acids Res. 2019, 47, 7018–7034. [Google Scholar] [CrossRef] [PubMed]
- Rakwalska, M.; Rospert, S. The ribosome-bound chaperones RAC and Ssb1/2p are required for accurate translation in Saccharomyces cerevisiae. Mol. Cell. Biol. 2004, 24, 9186–9197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, R.J.; Ziegelhoffer, T.; Nicolet, C.; Werner-Washburne, M.; Craig, E.A. The translation machinery and 70 kd heat shock protein cooperate in protein synthesis. Cell 1992, 71, 97–105. [Google Scholar] [CrossRef]
- Gumiero, A.; Conz, C.; Gesé, G.V.; Zhang, Y.; Weyer, F.A.; Lapouge, K.; Kappes, J.; von Plehwe, U.; Schermann, G.; Fitzke, E.; et al. Interaction of the cotranslational Hsp70 Ssb with ribosomal proteins and rRNA depends on its lid domain. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Stein, K.C.; Kriel, A.; Frydman, J. Nascent Polypeptide Domain Topology and Elongation Rate Direct the Cotranslational Hierarchy of Hsp70 and TRiC/CCT. Mol. Cell 2019, 75, 1117–1130.e5. [Google Scholar] [CrossRef] [PubMed]
- Chiabudini, M.; Conz, C.; Reckmann, F.; Rospert, S. Ribosome-associated complex and Ssb are required for translational repression induced by polylysine segments within nascent chains. Mol. Cell. Biol. 2012, 32, 4769–4779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiabudini, M.; Tais, A.; Zhang, Y.; Hayashi, S.; Wölfle, T.; Fitzke, E.; Rospert, S. Release factor eRF3 mediates premature translation termination on polylysine-stalled ribosomes in Saccharomyces cerevisiae. Mol. Cell. Biol. 2014, 34, 4062–4076. [Google Scholar] [CrossRef] [Green Version]
- Peisker, K.; Chiabudini, M.; Rospert, S. The ribosome-bound Hsp70 homolog Ssb of Saccharomyces cerevisiae. Biochim. Biophys. Acta 2010, 1803, 662–672. [Google Scholar] [CrossRef] [Green Version]
- Zuzow, N.; Ghosh, A.; Leonard, M.; Liao, J.; Yang, B.; Bennett, E.J. Mapping the mammalian ribosome quality control complex interactome using proximity labeling approaches. Mol. Biol. Cell 2018, 29, 1258–1269. [Google Scholar] [CrossRef]
- Shcherbik, N.; Chernova, T.A.; Chernoff, Y.O.; Pestov, D.G. Distinct types of translation termination generate substrates for ribosome-associated quality control. Nucleic Acids Res. 2016, 44, 6840–6852. [Google Scholar] [CrossRef] [Green Version]
- Mnaimneh, S.; Davierwala, A.P.; Haynes, J.; Moffat, J.; Peng, W.-T.; Zhang, W.; Yang, X.; Pootoolal, J.; Chua, G.; Lopez, A.; et al. Exploration of essential gene functions via titratable promoter alleles. Cell 2004, 118, 31–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyumkis, D.; Oliveira dos Passos, D.; Tahara, E.B.; Webb, K.; Bennett, E.J.; Vinterbo, S.; Potter, C.S.; Carragher, B.; Joazeiro, C.A.P. Structural basis for translational surveillance by the large ribosomal subunit-associated protein quality control complex. Proc. Natl. Acad. Sci. USA 2014, 111, 15981–15986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczanowska, M.; Rydén-Aulin, M. Ribosome biogenesis and the translation process in Escherichia coli. Microbiol. Mol. Biol. Rev. 2007, 71, 477–494. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.; Brown, A.; Santhanam, B.; Hegde, R.S. Structure and assembly pathway of the ribosome quality control complex. Mol. Cell 2015, 57, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.; Hegde, R.S. Reconstitution of a minimal ribosome-associated ubiquitination pathway with purified factors. Mol. Cell 2014, 55, 880–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, S.; von der Malsburg, K.; Hegde, R.S. Listerin-dependent nascent protein ubiquitination relies on ribosome subunit dissociation. Mol. Cell 2013, 50, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Prunuske, A.J.; Waltner, J.K.; Kuhn, P.; Gu, B.; Craig, E.A. Role for the molecular chaperones Zuo1 and Ssz1 in quorum sensing via activation of the transcription factor Pdr1. Proc. Natl. Acad. Sci. USA 2012, 109, 472–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosis, E.; Salomon, D.; Ohayon, O.; Sivan, G.; Bar-Nun, S.; Rabinovich, E. Ssz1 restores endoplasmic reticulum-associated protein degradation in cells expressing defective cdc48-ufd1-npl4 complex by upregulating cdc48. Genetics 2010, 184, 695–706. [Google Scholar] [CrossRef] [Green Version]
- Shorter, J.; Lindquist, S. Hsp104, Hsp70 and Hsp40 interplay regulates formation, growth and elimination of Sup35 prions. EMBO J. 2008, 27, 2712–2724. [Google Scholar] [CrossRef] [Green Version]
- Dimitrova, L.N.; Kuroha, K.; Tatematsu, T.; Inada, T. Nascent peptide-dependent translation arrest leads to Not4p-mediated protein degradation by the proteasome. J. Biol. Chem. 2009, 284, 10343–10352. [Google Scholar] [CrossRef] [Green Version]
- Zurita Rendón, O.; Fredrickson, E.K.; Howard, C.J.; Van Vranken, J.; Fogarty, S.; Tolley, N.D.; Kalia, R.; Osuna, B.A.; Shen, P.S.; Hill, C.P.; et al. Vms1p is a release factor for the ribosome-associated quality control complex. Nat. Commun. 2018, 9, 2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyumkis, D.; Doamekpor, S.K.; Bengtson, M.H.; Lee, J.-W.; Toro, T.B.; Petroski, M.D.; Lima, C.D.; Potter, C.S.; Carragher, B.; Joazeiro, C.A.P. Single-particle EM reveals extensive conformational variability of the Ltn1 E3 ligase. Proc. Natl. Acad. Sci. USA 2013, 110, 1702–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joazeiro, C.A.P. Ribosomal Stalling During Translation: Providing Substrates for Ribosome-Associated Protein Quality Control. Annu. Rev. Cell Dev. Biol. 2017, 33, 343–368. [Google Scholar] [CrossRef] [PubMed]
- Sitron, C.S.; Park, J.H.; Brandman, O. Asc1, Hel2, and Slh1 couple translation arrest to nascent chain degradation. RNA 2017, 23, 798–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shcherbik, N. Golgi-mediated glycosylation determines residency of the T2 RNase Rny1p in Saccharomyces cerevisiae. Traffic 2013, 14, 1209–1227. [Google Scholar] [CrossRef]
- Longtine, M.S.; McKenzie, A.; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Reference |
|---|---|---|
| BY4741 | MATa his3-1 leu2-0 met15-0 ura3-0 | Thermo Fisher |
| zuo1Δ | MATa his3-1 leu2-0 met15-0 ura3-0 zuo1Δ::KANMX6 | Thermo Fisher |
| ssz1Δ | MATa his3-1 leu2-0 met15-0 ura3-0 ssz1Δ::KANMX6 | Thermo Fisher |
| ssb1Δ ssb2Δ | MATa his3-1 leu2-0 met15-0 ura3-0 ssb1Δ::KANMX6 ssb2Δ::LEU2 | This study |
| PTET-O7-CDC48 | MATa URA3::CMV-tTA his3-1 leu2-0 met15-0 pCDC48::KanR-tet07-TATA | Thermo Fisher |
| PTET-O7-CDC48 ltn1Δ | MATa URA3::CMV-tTA his3-1 leu2-0 met15-0 pCDC48::KanR-tet07-TATA ltn1Δ::his5+ | This study |
| PTET-O7-CDC48 ssb1Δ ssb2Δ | MATa URA3::CMV-tTA his3-1 leu2-0 met15-0 pCDC48::KanR-tet07-TATA ssb1Δ::his5+ ssb2Δ::LEU2 | This study |
| PTET-O7-CDC48 ssz1Δ | MATa URA3::CMV-tTA his3-1 leu2-0 met15-0 pCDC48::KanR-tet07-TATA ssz1Δ::his5+ | This study |
| PTET-O7-CDC48 zuo1Δ | MATa URA3::CMV-tTA his3-1 leu2-0 met15-0 pCDC48::KanR-tet07-TATA zuo1Δ::LEU2 | This study |
| PTET-O7-CDC48 ssz1Δ ltn1Δ | MATa URA3::CMV-tTA his3-1 leu2-0 met15-0 pCDC48::KanR-tet07-TATA ssz1Δ::his5+ ltn1Δ::LEU2 | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, A.; Shcherbik, N. Cooperativity between the Ribosome-Associated Chaperone Ssb/RAC and the Ubiquitin Ligase Ltn1 in Ubiquitination of Nascent Polypeptides. Int. J. Mol. Sci. 2020, 21, 6815. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186815
Ghosh A, Shcherbik N. Cooperativity between the Ribosome-Associated Chaperone Ssb/RAC and the Ubiquitin Ligase Ltn1 in Ubiquitination of Nascent Polypeptides. International Journal of Molecular Sciences. 2020; 21(18):6815. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186815
Chicago/Turabian StyleGhosh, Arnab, and Natalia Shcherbik. 2020. "Cooperativity between the Ribosome-Associated Chaperone Ssb/RAC and the Ubiquitin Ligase Ltn1 in Ubiquitination of Nascent Polypeptides" International Journal of Molecular Sciences 21, no. 18: 6815. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186815