Interactions among Genetic Background, Anesthetic Agent, and Oxygen Concentration Shape Blunt Traumatic Brain Injury Outcomes in Drosophila melanogaster



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Genetic Background Modulates the Effect of Anesthetic Exposure on Mortality Following TBI
2.2. Effects of ISO/O2 Exposure on Mortality Following TBI Stratifies Effects of O2 and SEVO/O2 Exposure on Mortality
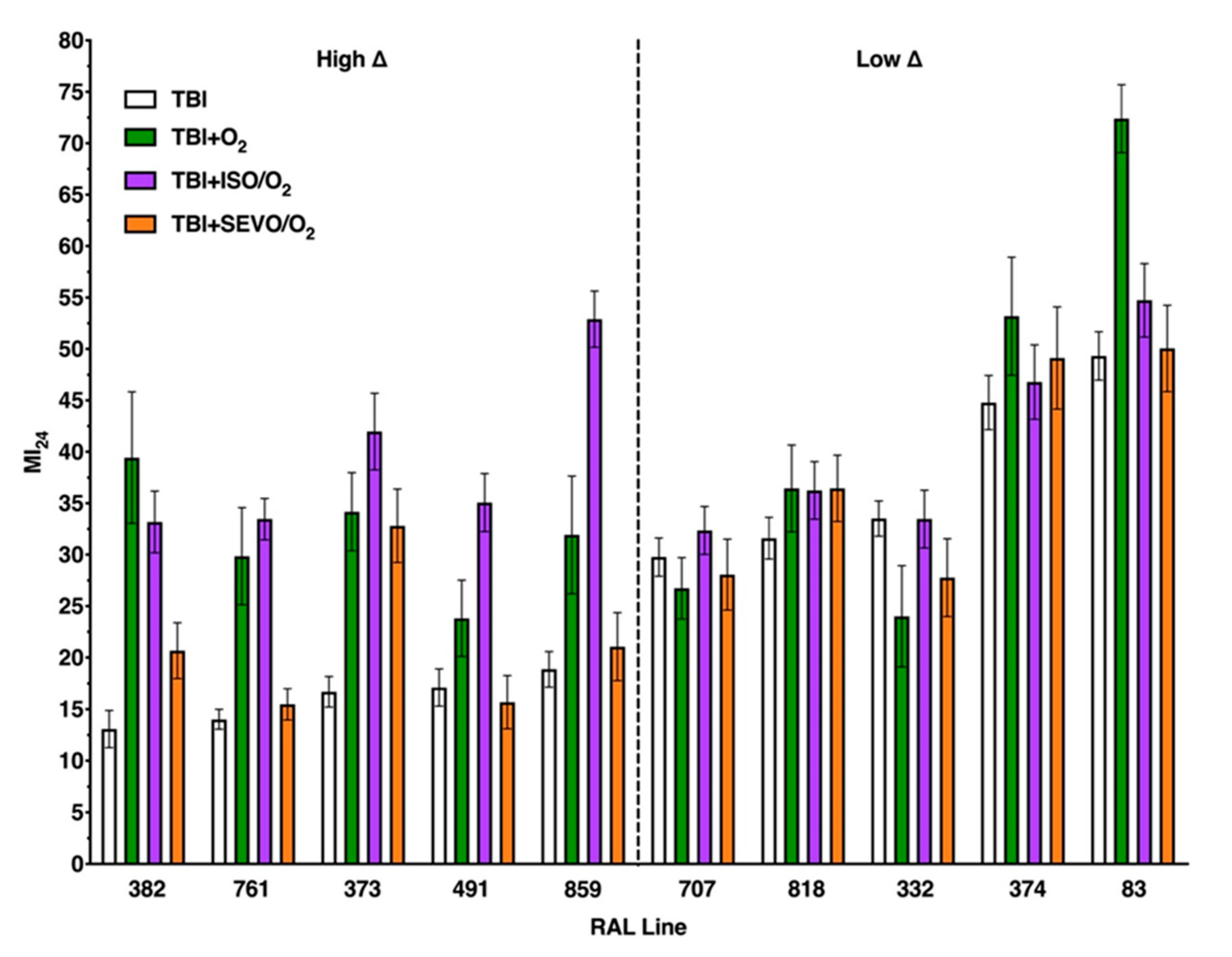
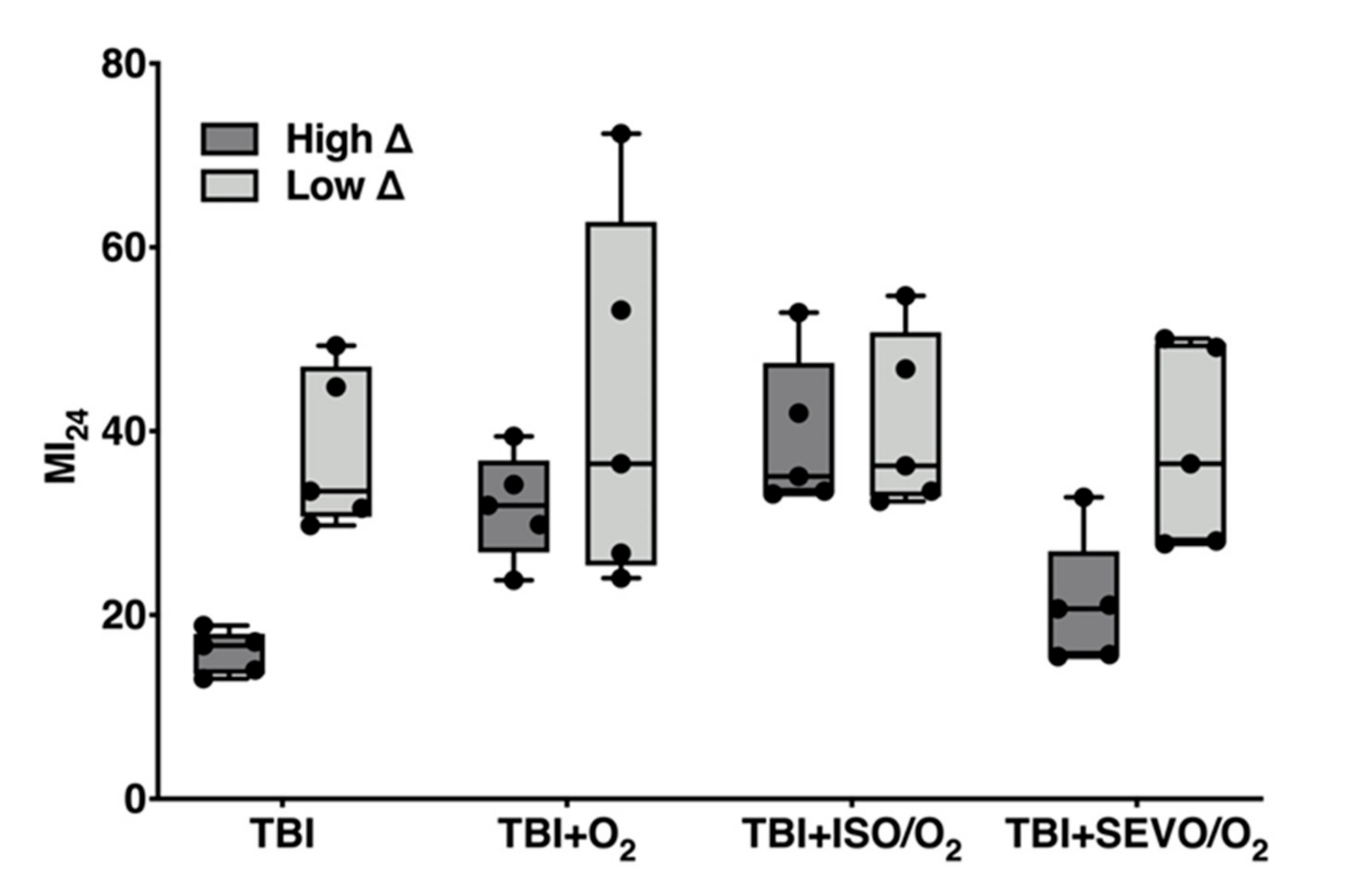
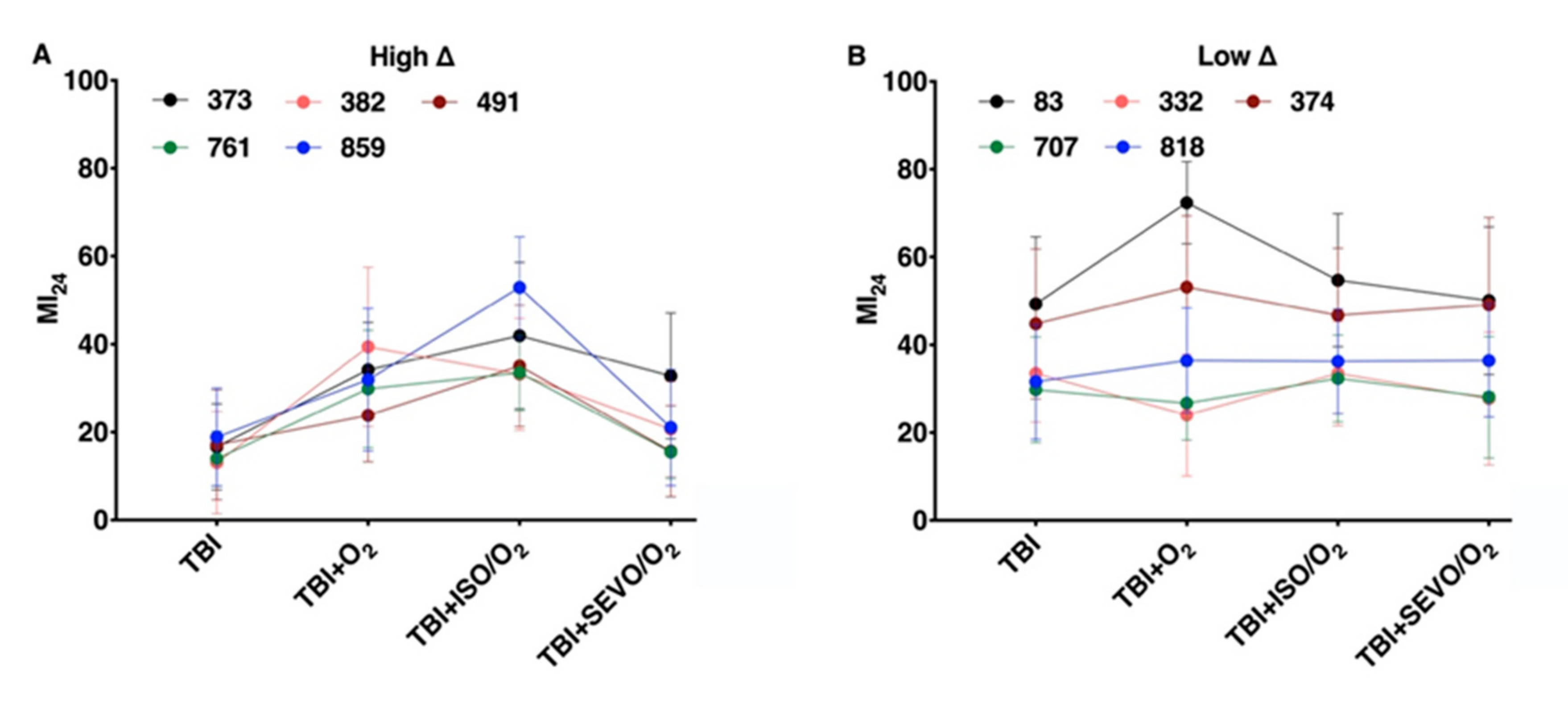
2.3. Variation in the MI24 of Individual Lines Following O2, ISO/O2, and SEVO/O2 Exposure after TBI Is More Common in the High ∆ Group than the Low ∆ Group
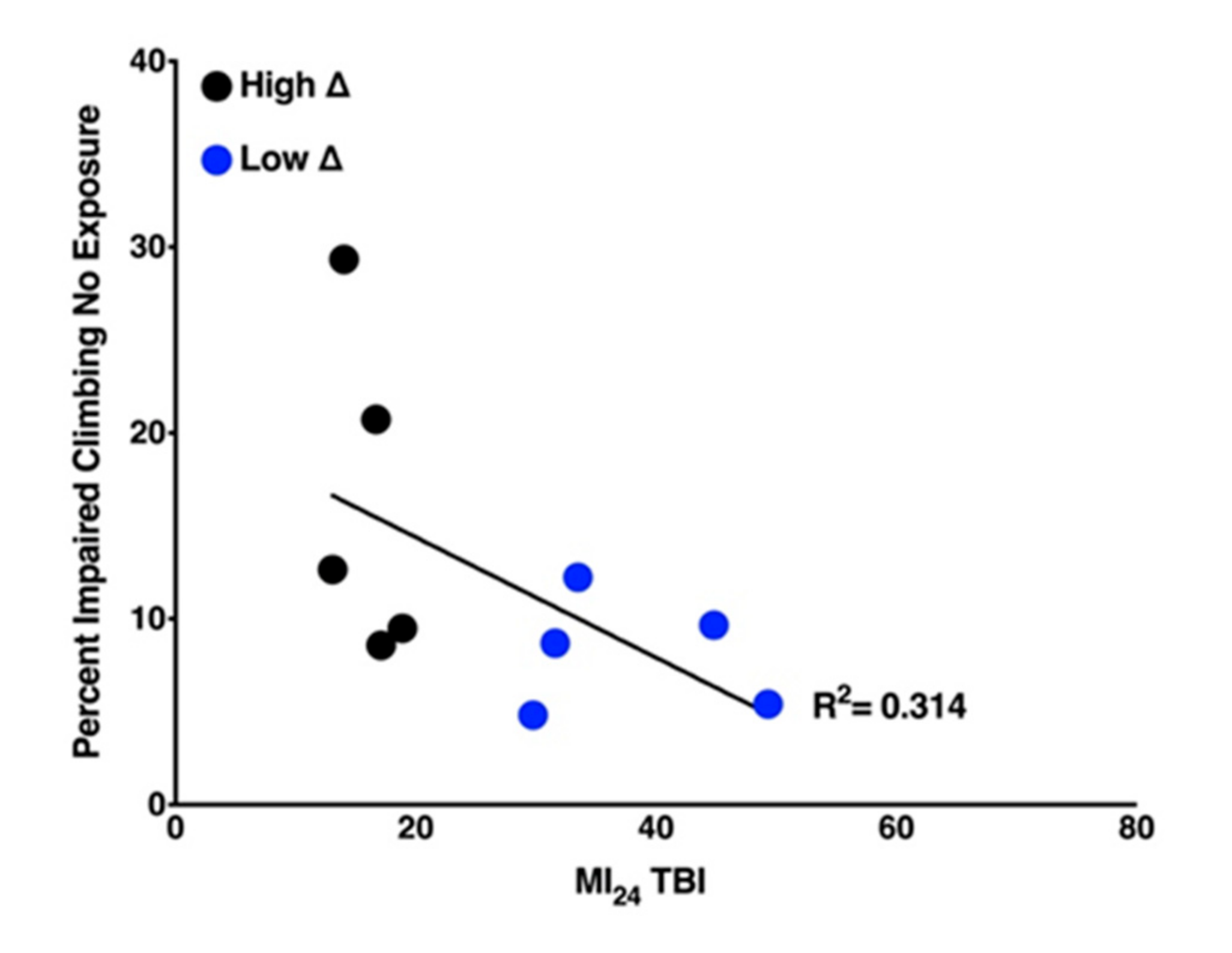
2.4. Locomotion of Uninjured Flies Has a Weak Negative Correlation with Mortality of Flies Following TBI
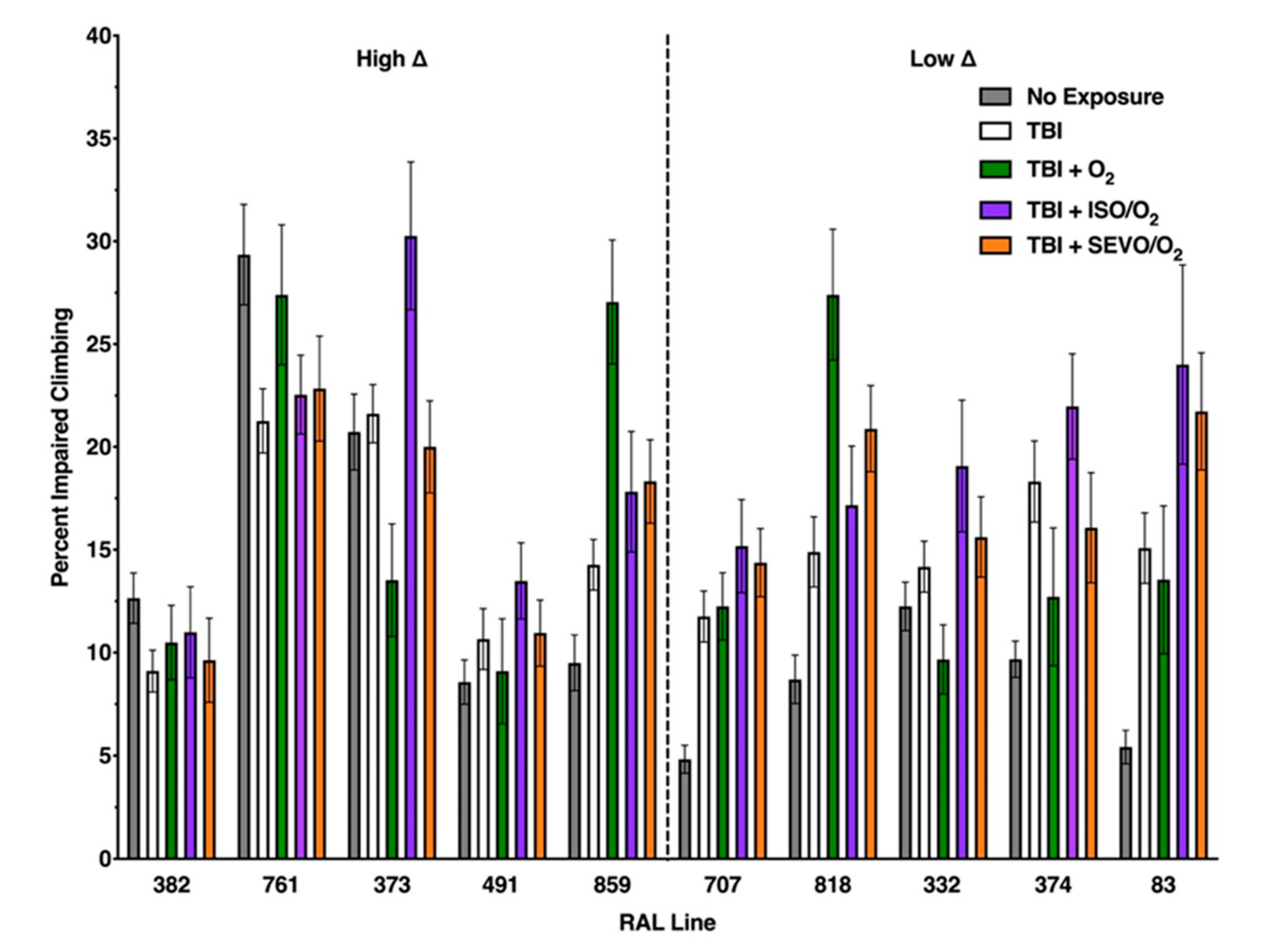
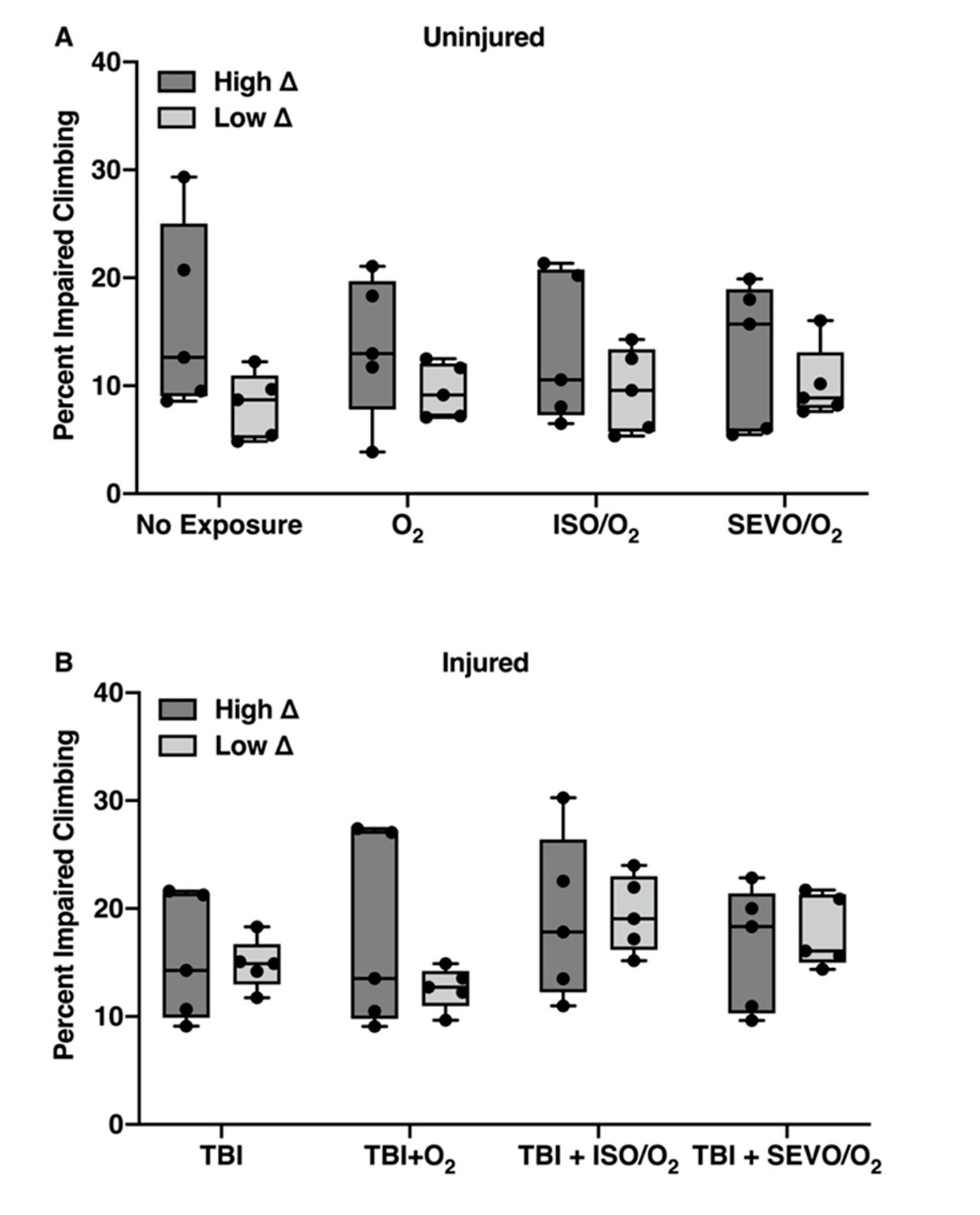
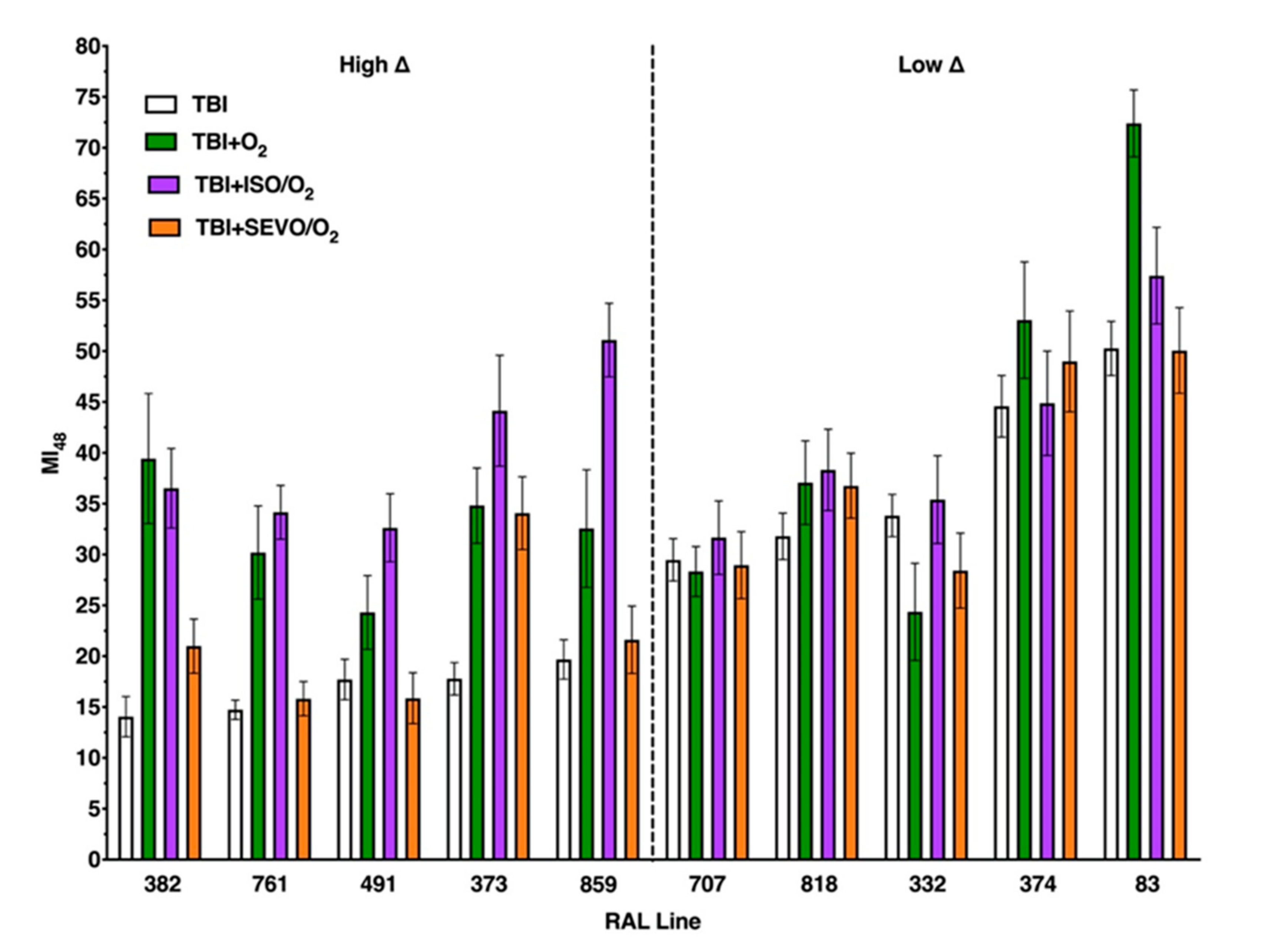
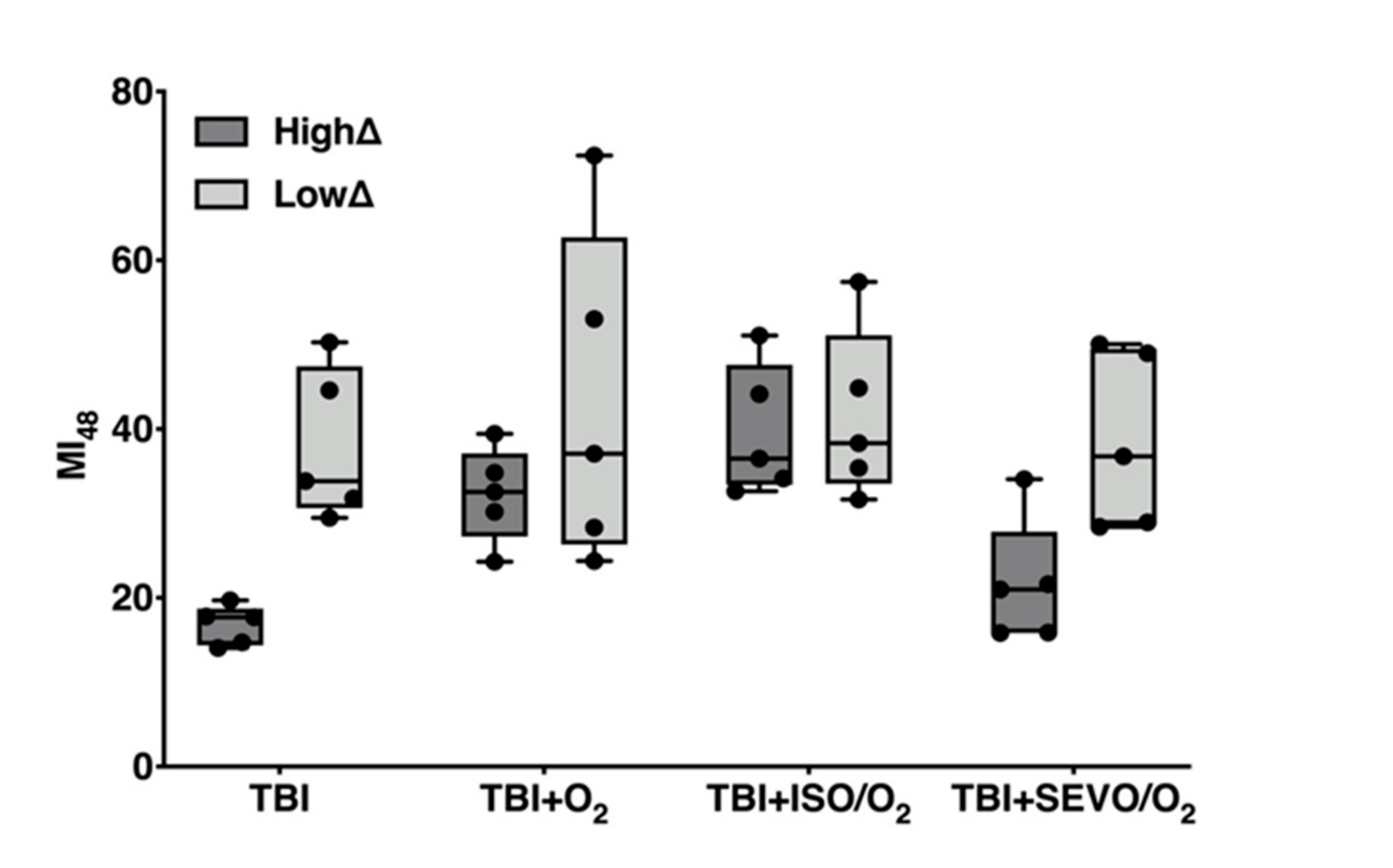
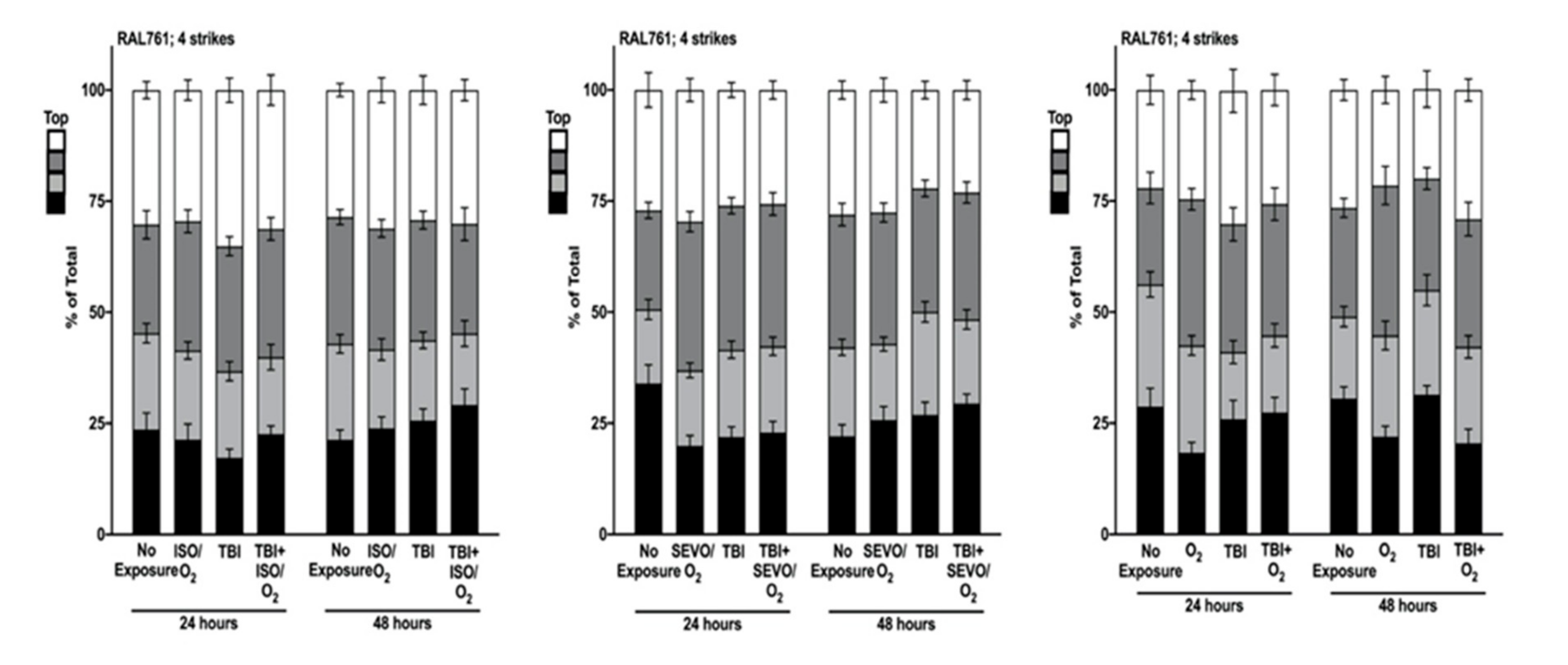
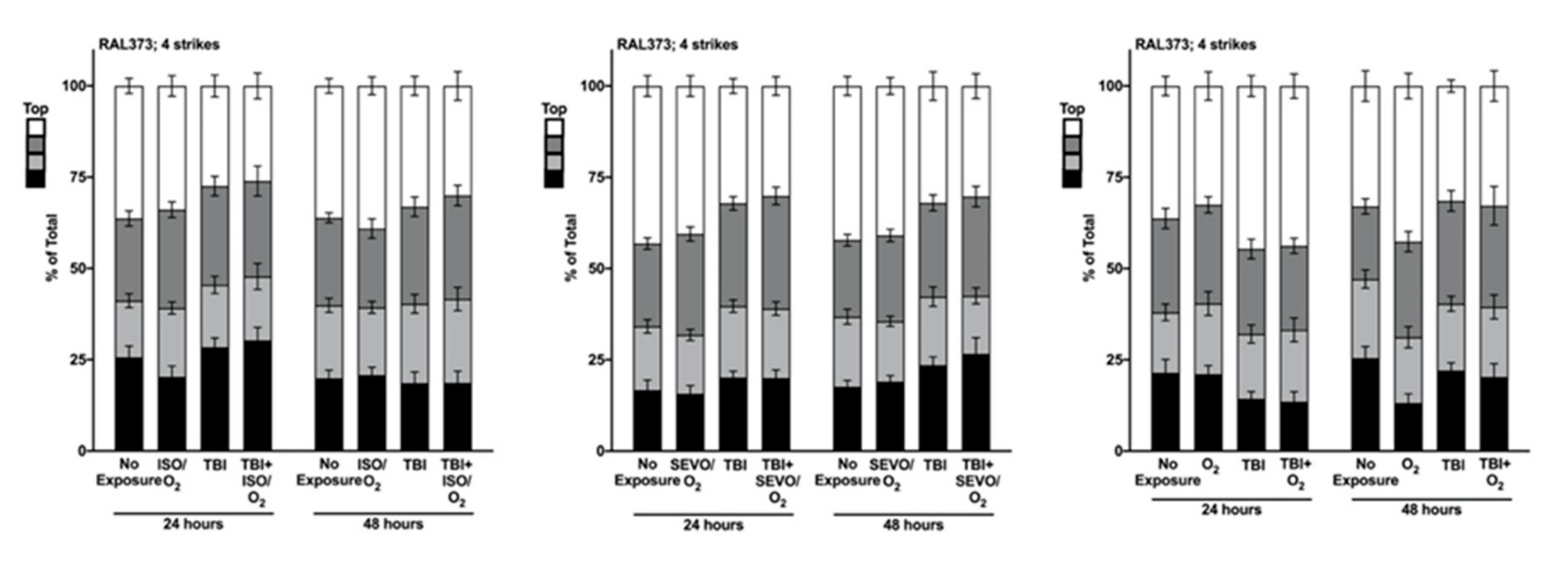
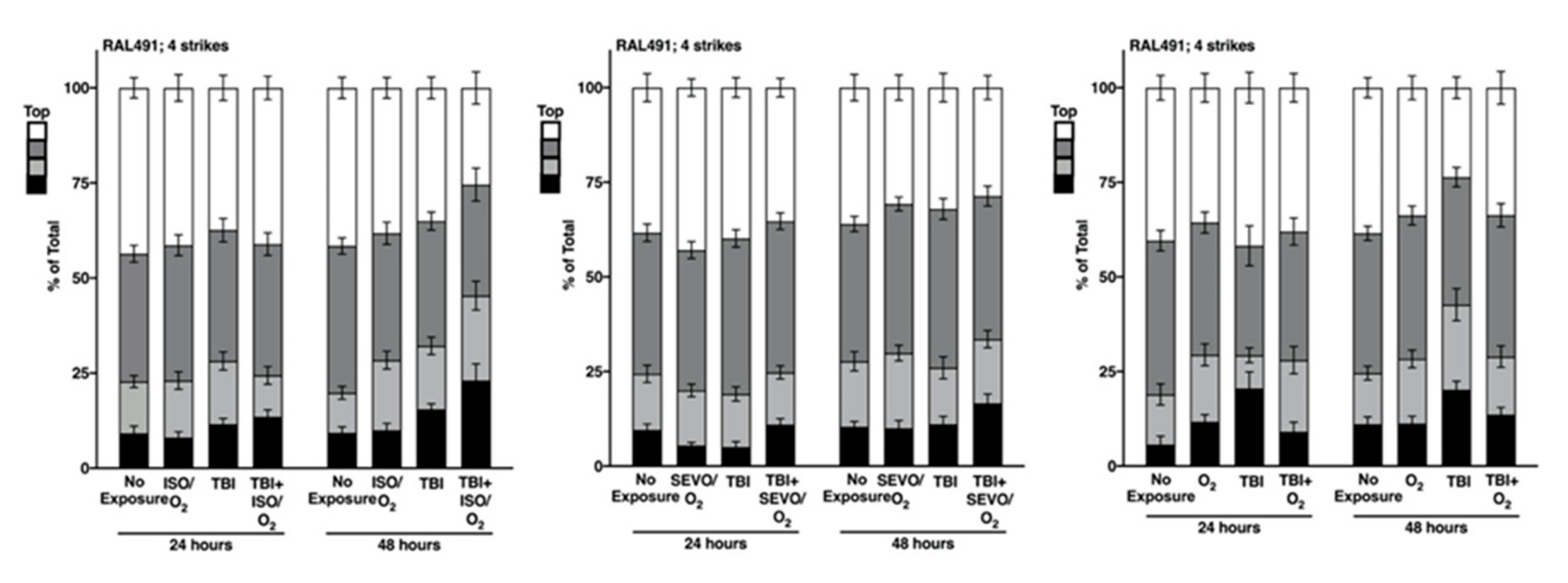
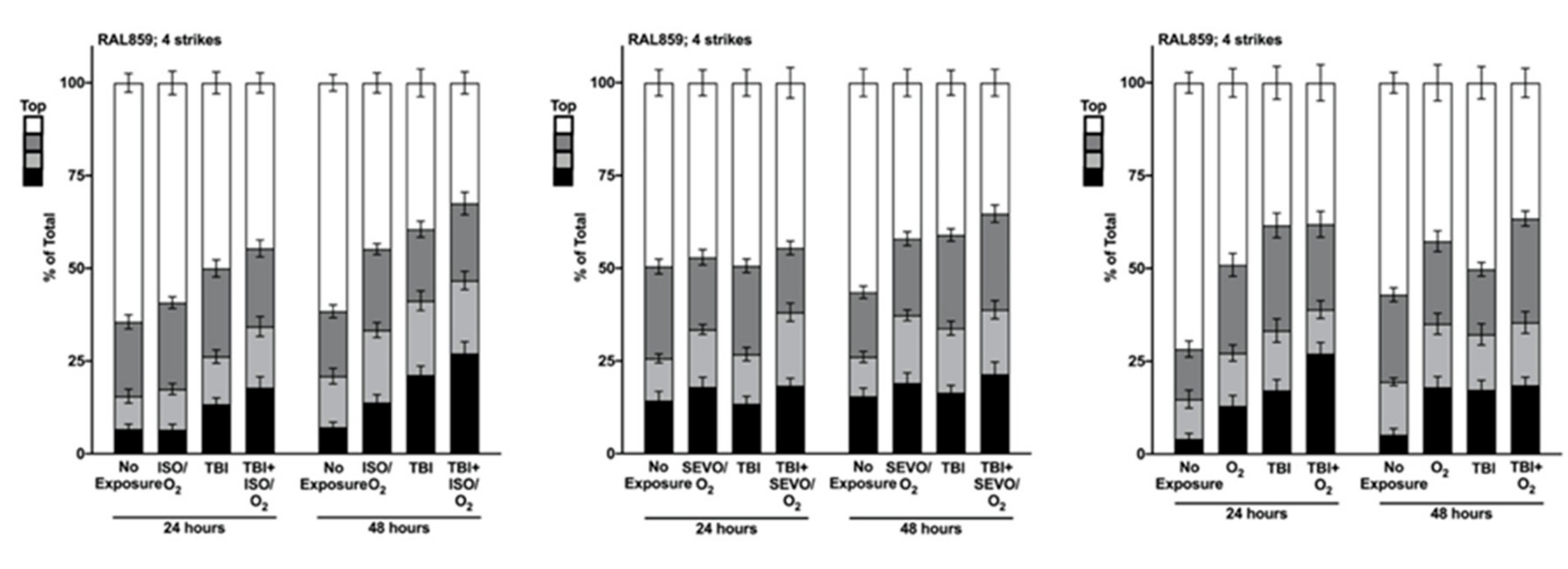
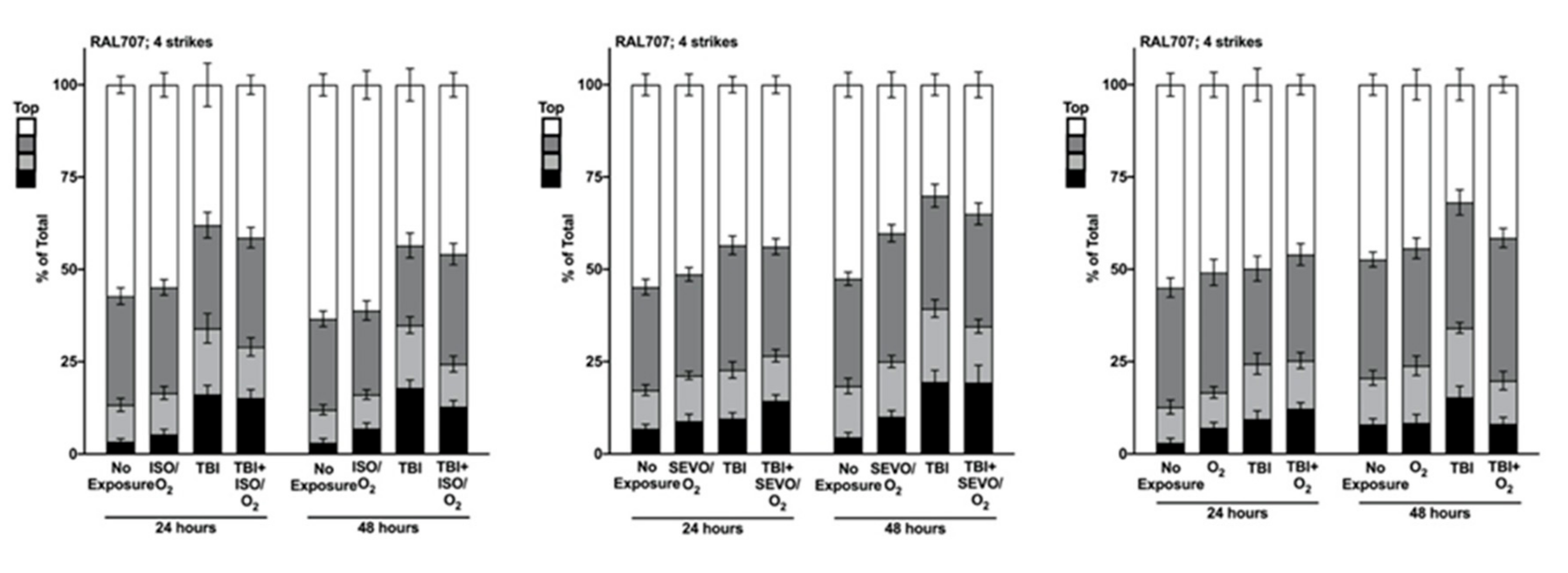
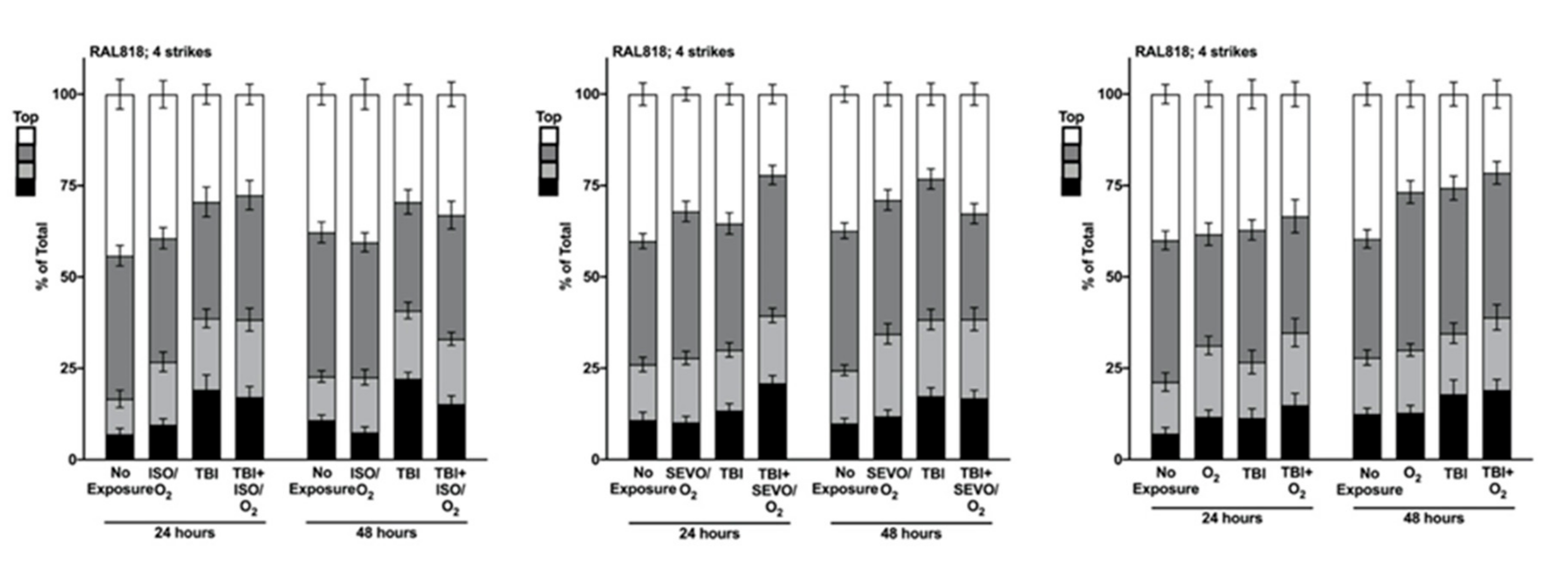
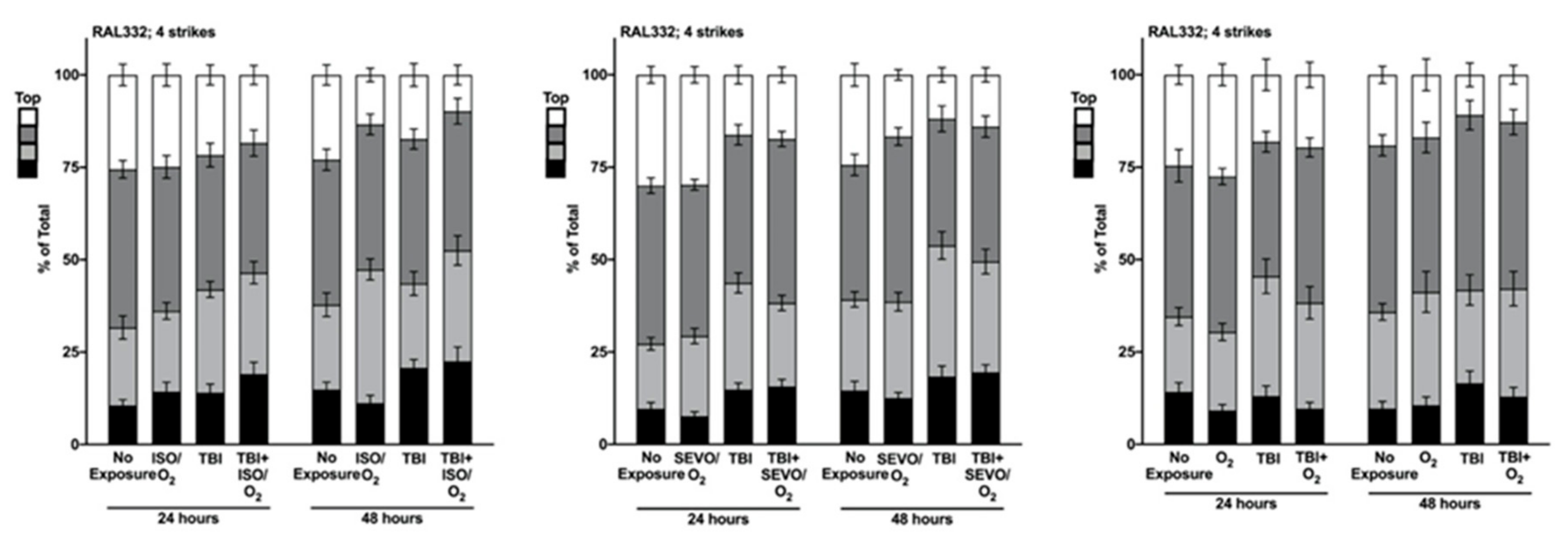
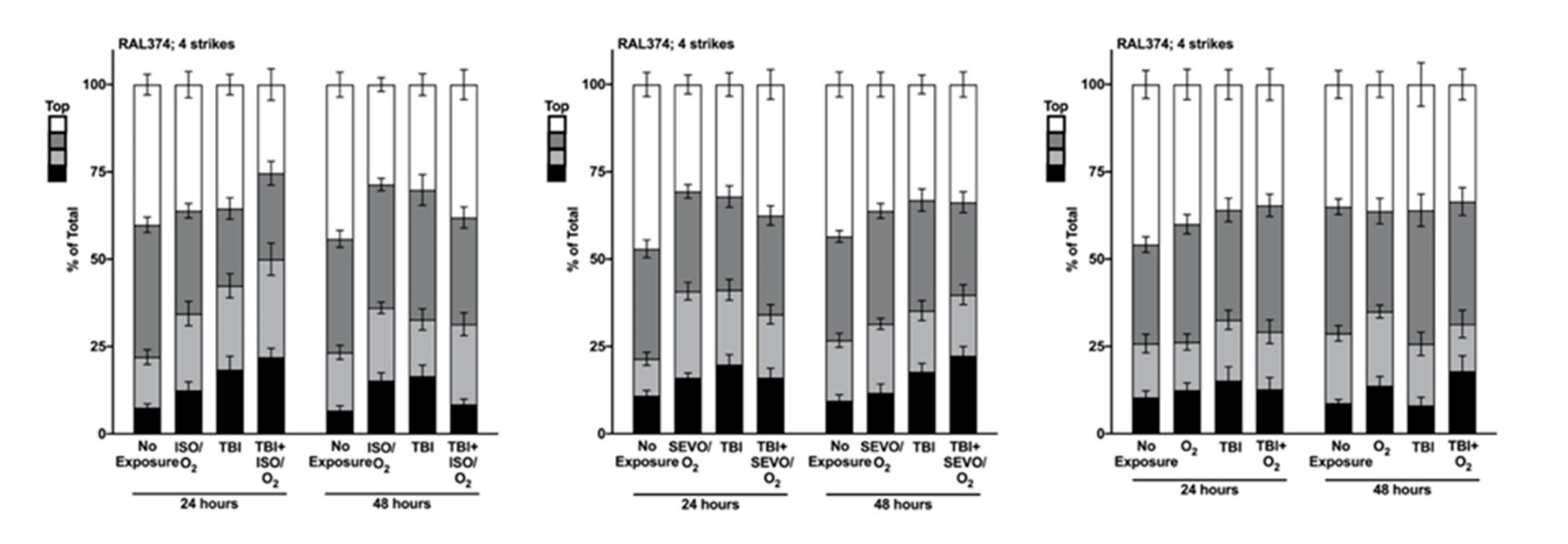
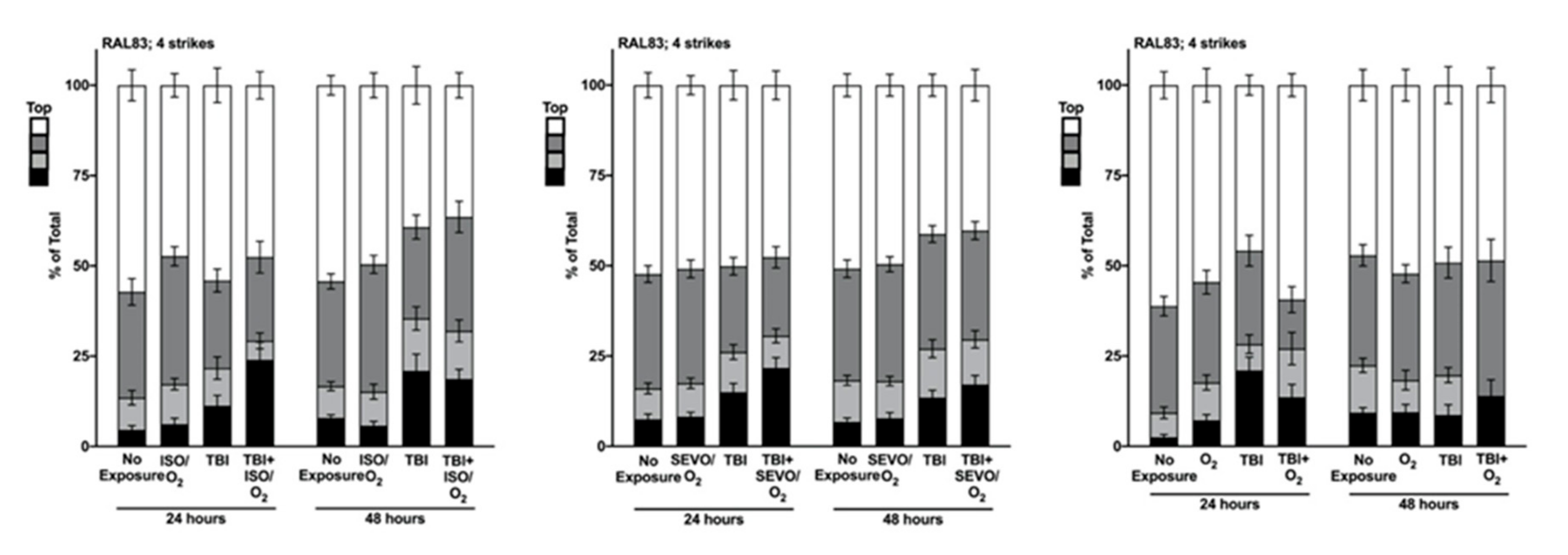
2.5. Genetic Background Modulates the Effect of Anesthetic Exposure on Locomotion Following TBI
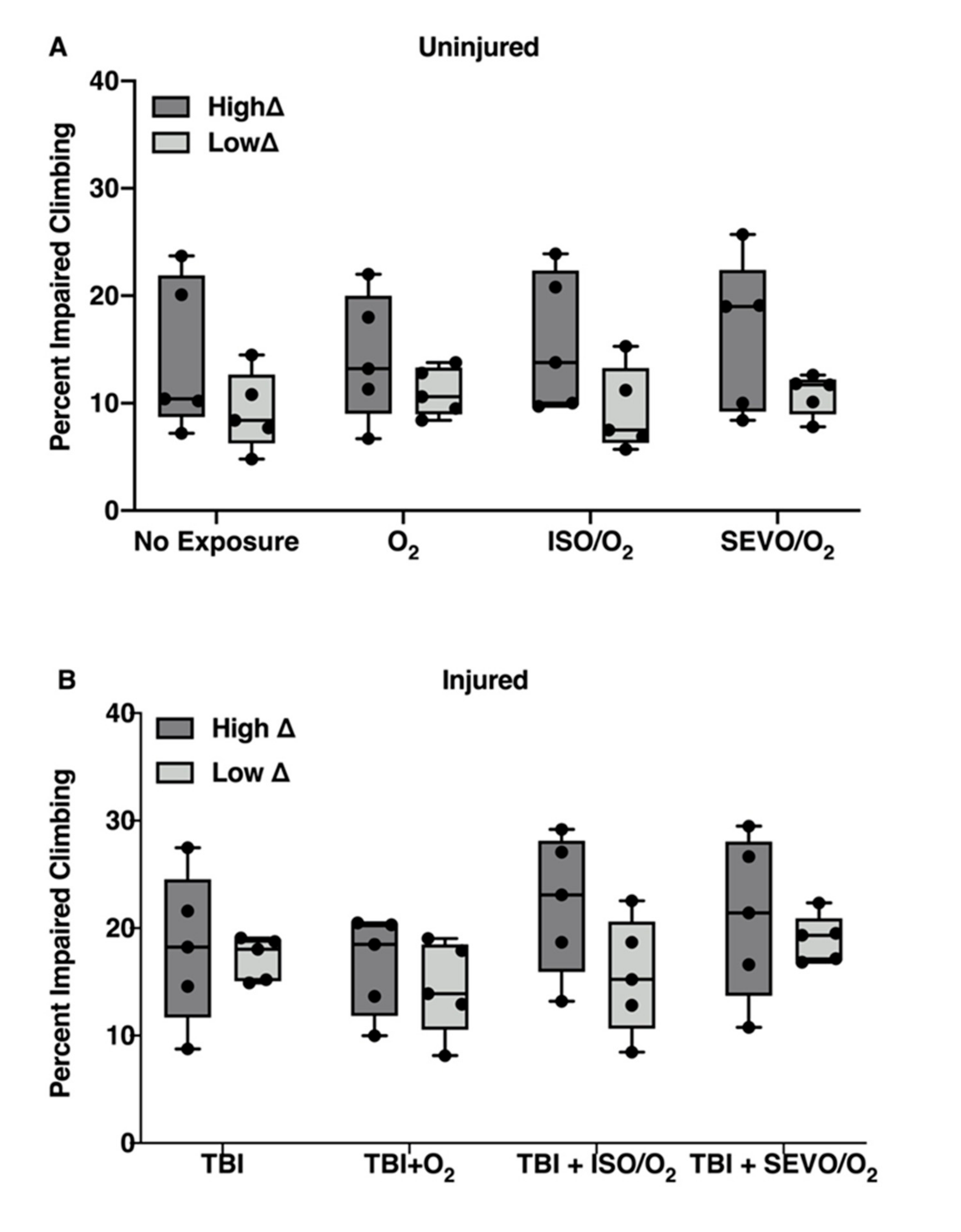
2.6. ISO/O2 and SEVO/O2 Exposure Impairs Climbing of the Low ∆ Group Following TBI
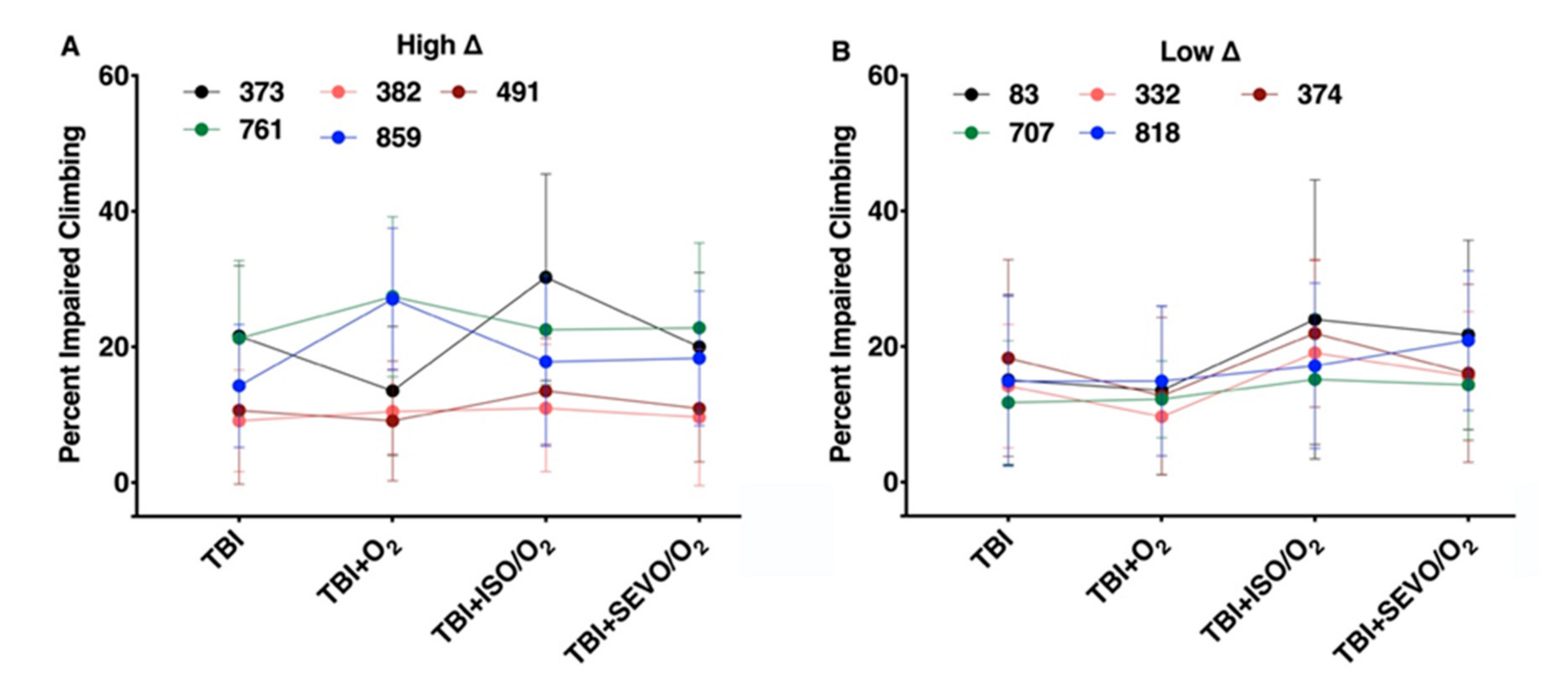
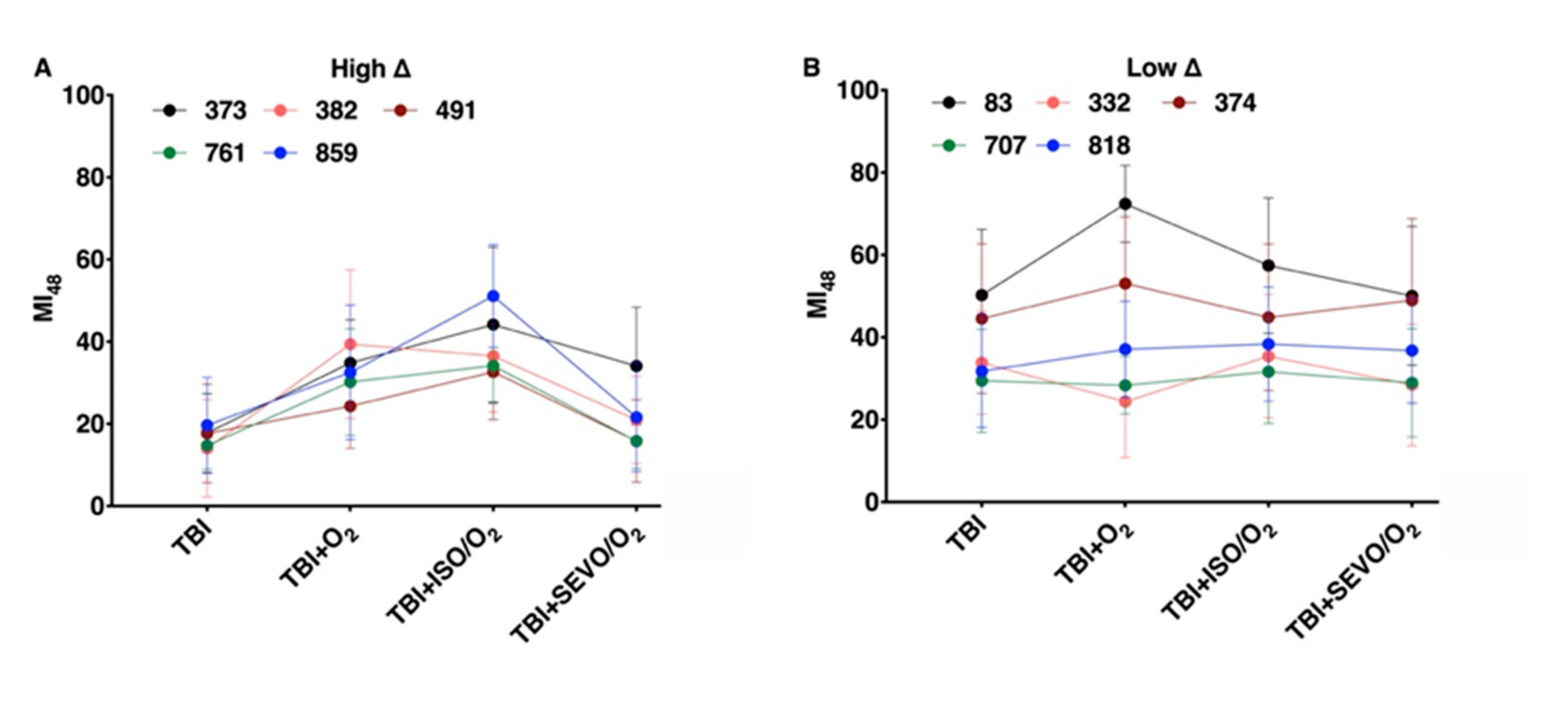
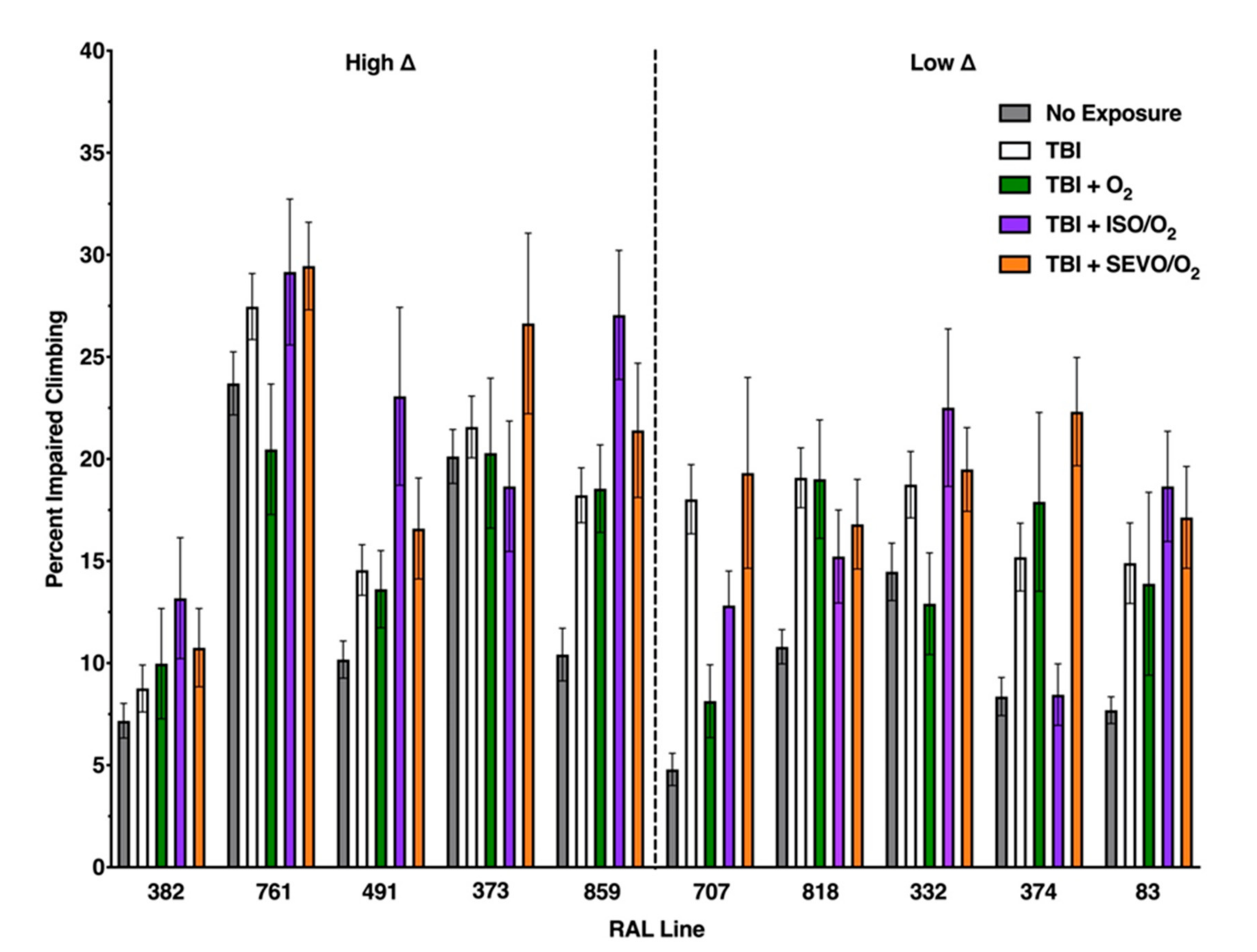
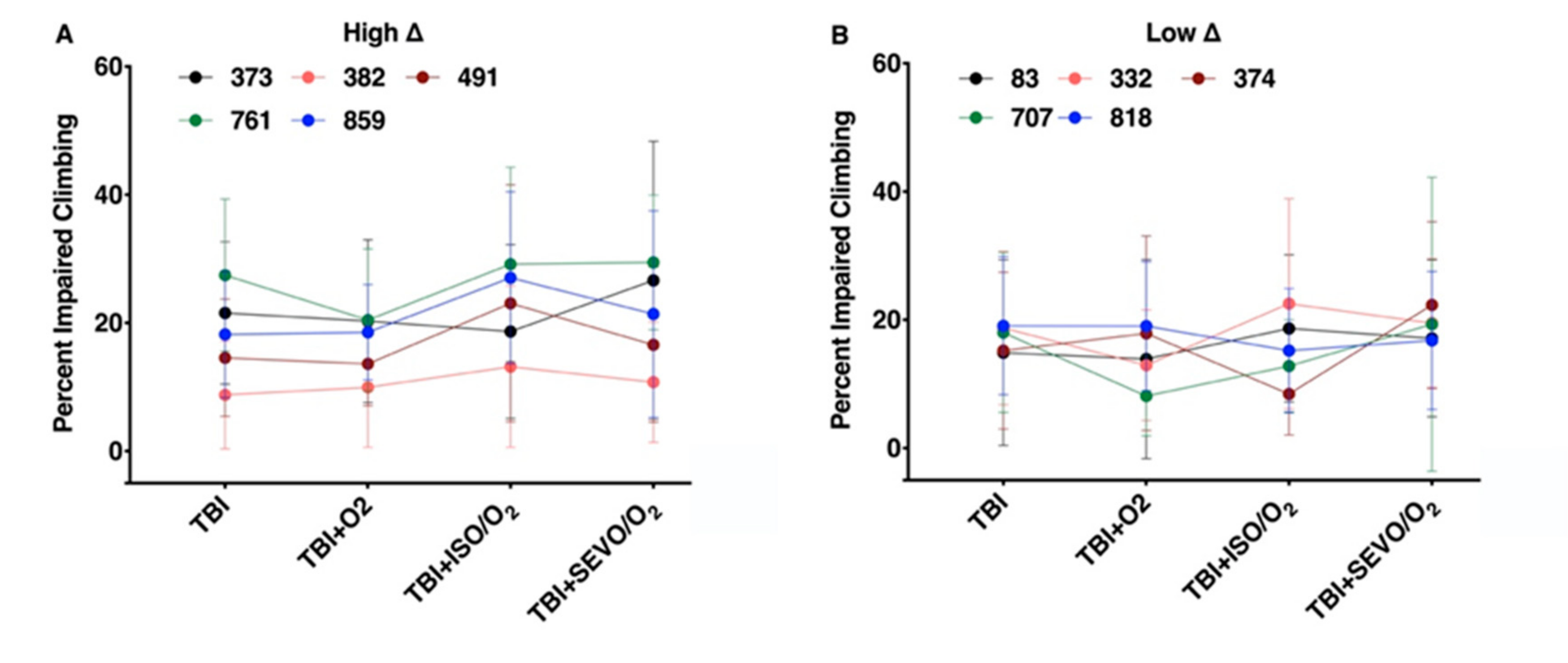
2.7. Percent Impaired Climbing of Individual Lines within the High ∆ Group but Not Low ∆ Groups Varies Following O2, ISO/O2, or SEVO/O2 Exposure after TBI
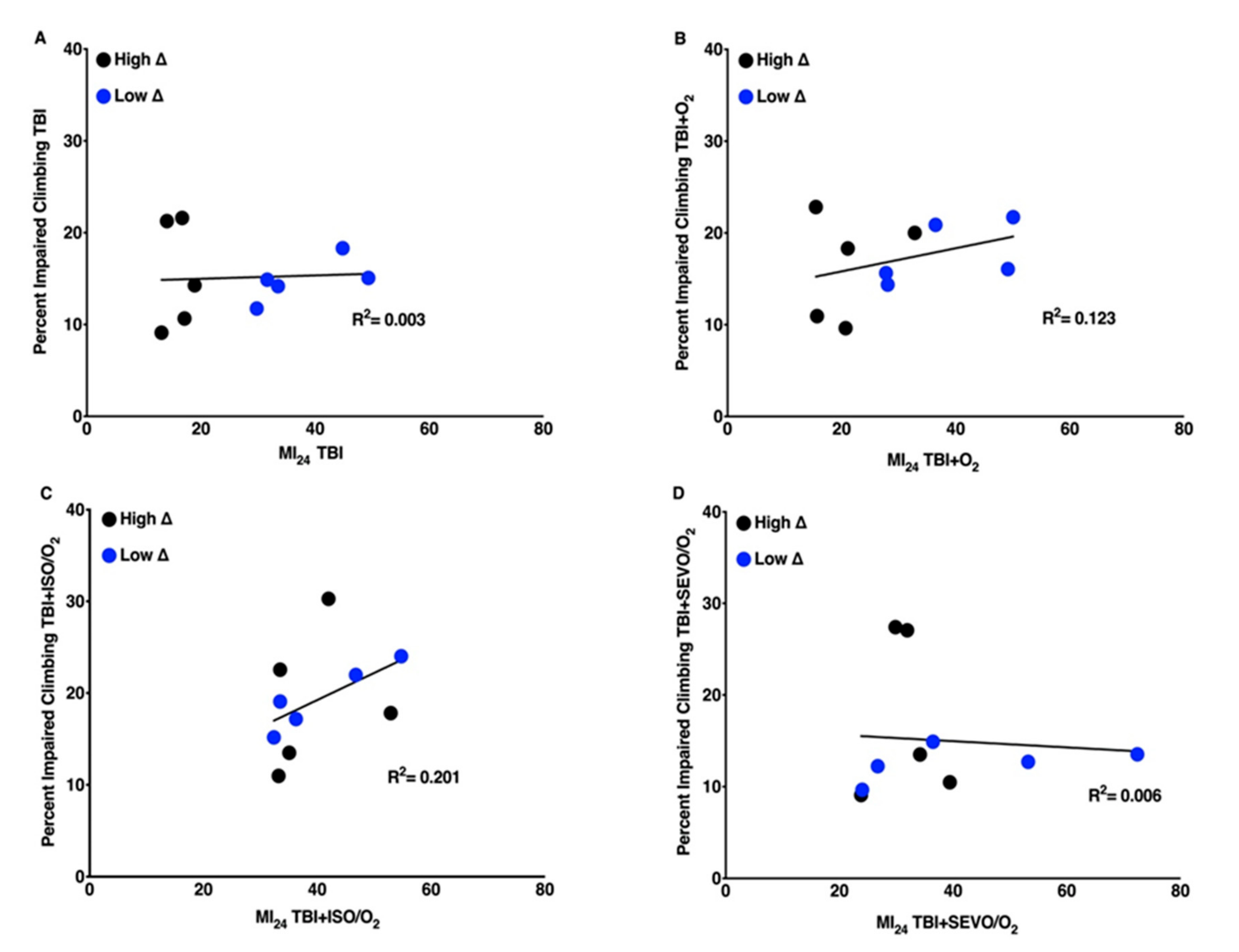
2.8. The Weak Correlation between the MI24 and Locomotion Impairment Is Lost after Interventions
3. Discussion
3.1. Resilience to TBI-Induced Mortality Correlates Inversely with Susceptibility to AiN
3.2. VGA-Toxicity Is Distinguishable from O2-Toxicity
3.3. Are Mortality and Impaired Locomotion Mechanistically Unrelated Phenotypes?
3.4. The Modifying Effects of ISO/O2 on Mortality and Locomotion Following TBI in Different Genetic Backgrounds Does Not Predict the Modifying Effects of O2 or SEVO/O2
3.5. Climbing Activity as Predictor of Resilience to TBI
3.6. Genetic Background Confers Discrete Sensitivity/Resilience to TBI, VGAs, and O2
3.7. Summary
4. Materials and Methods
4.1. TBI
4.2. Exposure to VGAs and O2
4.3. Genetic Background
4.4. High and Low ∆ Groups
4.5. Mortality
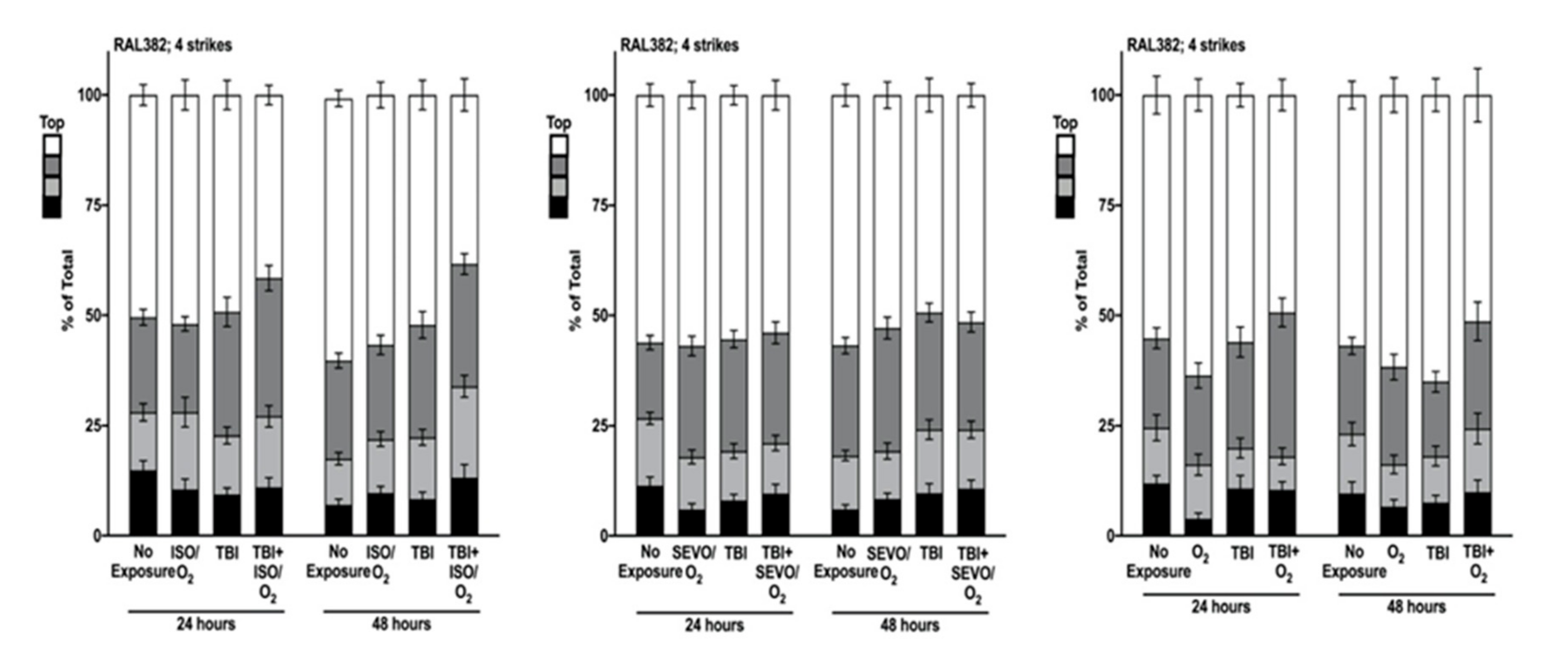
4.6. Impairment of Locomotion
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TBI | Traumatic brain injury |
| VGA | Volatile general anesthetic |
| ISO | Isoflurane |
| SEVO High Δ Low Δ | Sevoflurane High excess mortality Low excess mortality |
Appendix A
















References
- Dutton, R.P.; Stansbury, L.G.; Leone, S.; Kramer, E.; Hess, J.R.; Scalea, T.M. Trauma mortality in mature trauma systems: Are we doing better? An analysis of trauma mortality patterns, 1997-2008. J. Trauma 2010, 69, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Mahmood, A.; Chopp, M. Animal models of traumatic brain injury. Nat. Rev. Neurosci. 2013, 14, 128–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, S.M.; Conley, Y.P.; Wagner, A.K.; Jha, R.M.; Clark, R.S.; Poloyac, S.M.; Kochanek, P.M.; Empey, P.E. The pharmacogenomics of severe traumatic brain injury. Pharmacogenomics 2017, 18, 1413–1425. [Google Scholar] [CrossRef] [PubMed]
- Ganetzky, B.; Wassarman, D.A. Non-mammalian Animal Models Offer New Perspectives on the Treatment of TBI. Curr. Phys. Med. Rehabil. Rep. 2016, 4, 1–4. [Google Scholar] [CrossRef]
- Katzenberger, R.J.; Ganetzky, B.; Wassarman, D.A. Age and Diet Affect Genetically Separable Secondary Injuries that Cause Acute Mortality Following Traumatic Brain Injury in Drosophila. G3 2016, 6, 4151–4166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzenberger, R.J.; Loewen, C.A.; Bockstruck, R.T.; Woods, M.A.; Ganetzky, B.; Wassarman, D.A. A Method to Inflict Closed Head Traumatic Brain Injury in Drosophila. J. Vis. Exp. 2015, 100, e52905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzenberger, R.J.; Loewen, C.A.; Wassarman, D.R.; Petersen, A.J.; Ganetzky, B.; Wassarman, D.A. A Drosophila model of closed head traumatic brain injury. Proc. Natl. Acad. Sci. USA 2013, 110, E4152–E4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olufs, Z.P.G.; Loewen, C.A.; Ganetzky, B.; Wassarman, D.A.; Perouansky, M. Genetic variability affects absolute and relative potencies and kinetics of the anesthetics isoflurane and sevoflurane in Drosophila melanogaster. Sci. Rep. 2018, 8, 2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffman, H.J.; Olufs, Z.P.G.; Lasarev, M.R.; Wassarman, D.A.; Perouansky, M. Ageing and genetic background influence anaesthetic effects in a D. melanogaster model of blunt trauma with brain injury(dagger). Br. J. Anaesth. 2020, 125, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Vutskits, L.; Xie, Z. Lasting impact of general anaesthesia on the brain: Mechanisms and relevance. Nat. Rev. Neurosci. 2016, 17, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Mackay, T.F.; Richards, S.; Stone, E.A.; Barbadilla, A.; Ayroles, J.F.; Zhu, D.; Casillas, S.; Han, Y.; Magwire, M.M.; Cridland, J.M.; et al. The Drosophila melanogaster Genetic Reference Panel. Nature 2012, 482, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rincon, F.; Kang, J.; Maltenfort, M.; Vibbert, M.; Urtecho, J.; Athar, M.K.; Jallo, J.; Pineda, C.C.; Tzeng, D.; McBride, W.; et al. Association between hyperoxia and mortality after stroke: A multicenter cohort study. Crit. Care Med. 2014, 42, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Rincon, F.; Kang, J.; Vibbert, M.; Urtecho, J.; Athar, M.K.; Jallo, J. Significance of arterial hyperoxia and relationship with case fatality in traumatic brain injury: A multicentre cohort study. J. Neurol. Neurosurg. Psychiatry 2014, 85, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Page, D.; Ablordeppey, E.; Wessman, B.T.; Mohr, N.M.; Trzeciak, S.; Kollef, M.H.; Roberts, B.W.; Fuller, B.M. Emergency department hyperoxia is associated with increased mortality in mechanically ventilated patients: A cohort study. Crit. Care 2018, 22, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiani, E.; Adrario, E.; Girardis, M.; Romano, R.; Pelaia, P.; Singer, M.; Donati, A. Arterial hyperoxia and mortality in critically ill patients: A systematic review and meta-analysis. Crit. Care 2014, 18, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rand, M.D. Drosophotoxicology: The growing potential for Drosophila in neurotoxicology. Neurotoxicol. Teratol. 2010, 32, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.A.; Olufs, Z.P.G.; Katzenberger, R.J.; Wassarman, D.A.; Perouansky, M. Anesthetics Influence Mortality in a Drosophila Model of Blunt Trauma with Traumatic Brain Injury. Anesth. Analg. 2018, 126, 1979–1986. [Google Scholar] [CrossRef] [PubMed]
- Fougere, B.; Daumas, M.; Lilamand, M.; Sourdet, S.; Delrieu, J.; Vellas, B.; Abellan van Kan, G. Association Between Frailty and Cognitive Impairment: Cross-Sectional Data From Toulouse Frailty Day Hospital. J. Am. Med. Dir. Assoc. 2017, 18, 990.e1–990.e5. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yolitz, J.; Wang, C.; Spangler, E.; Zhan, M.; Zou, S. Aging studies in Drosophila melanogaster. Methods Mol. Biol. 2013, 1048, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Petersen, A.J.; Rimkus, S.A.; Wassarman, D.A. ATM kinase inhibition in glial cells activates the innate immune response and causes neurodegeneration in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, E656–E664. [Google Scholar] [CrossRef] [PubMed] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scharenbrock, A.R.; Schiffman, H.J.; Olufs, Z.P.G.; Wassarman, D.A.; Perouansky, M. Interactions among Genetic Background, Anesthetic Agent, and Oxygen Concentration Shape Blunt Traumatic Brain Injury Outcomes in Drosophila melanogaster. Int. J. Mol. Sci. 2020, 21, 6926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186926
Scharenbrock AR, Schiffman HJ, Olufs ZPG, Wassarman DA, Perouansky M. Interactions among Genetic Background, Anesthetic Agent, and Oxygen Concentration Shape Blunt Traumatic Brain Injury Outcomes in Drosophila melanogaster. International Journal of Molecular Sciences. 2020; 21(18):6926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186926
Chicago/Turabian StyleScharenbrock, Amanda R., Hannah J. Schiffman, Zachariah P. G. Olufs, David A. Wassarman, and Misha Perouansky. 2020. "Interactions among Genetic Background, Anesthetic Agent, and Oxygen Concentration Shape Blunt Traumatic Brain Injury Outcomes in Drosophila melanogaster" International Journal of Molecular Sciences 21, no. 18: 6926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186926