4-Acetylantroquinonol B Inhibits Osteoclastogenesis by Inhibiting the Autophagy Pathway in a Simulated Microgravity Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
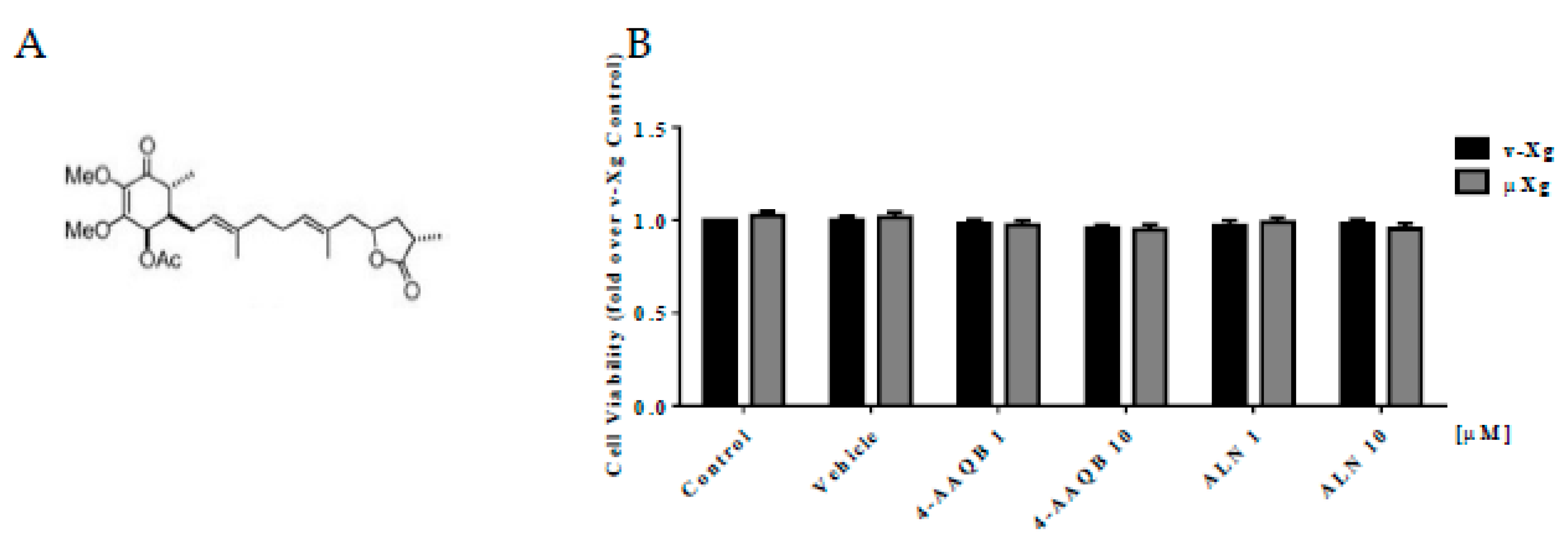
2.1. 4-AAQB
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Rotary Cell Culture System (RCCS)
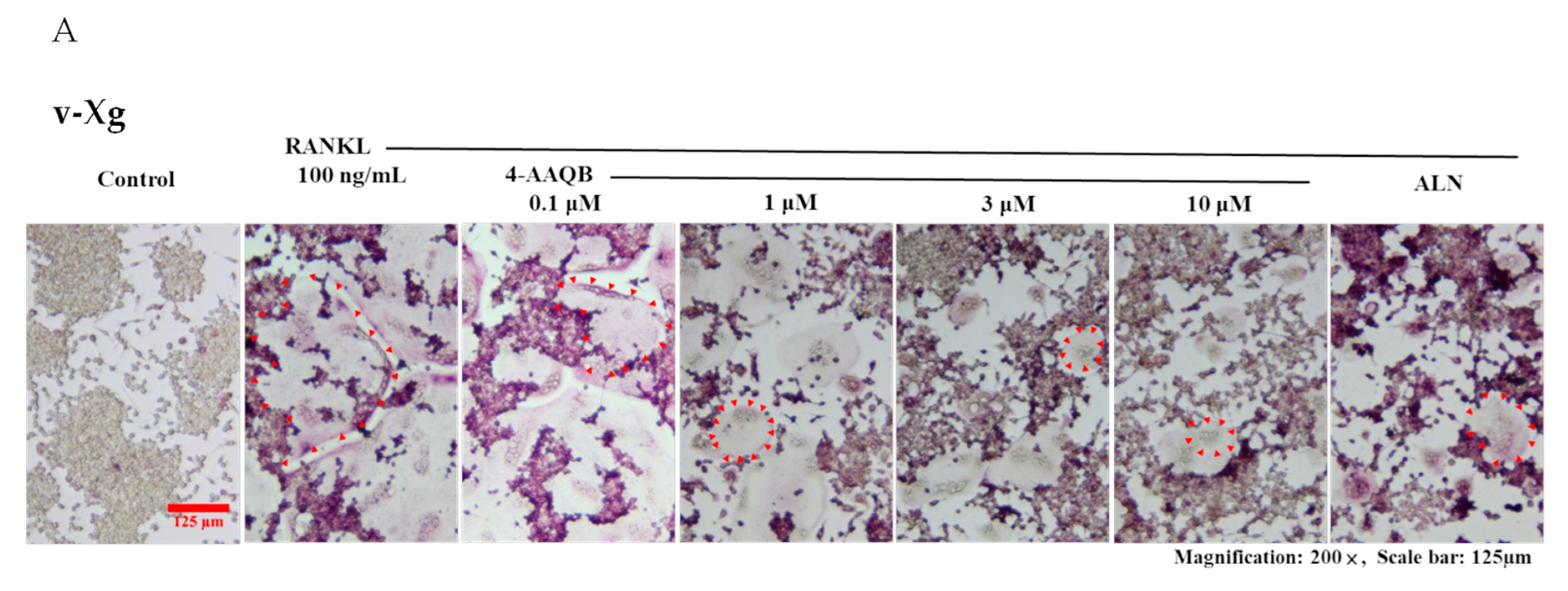
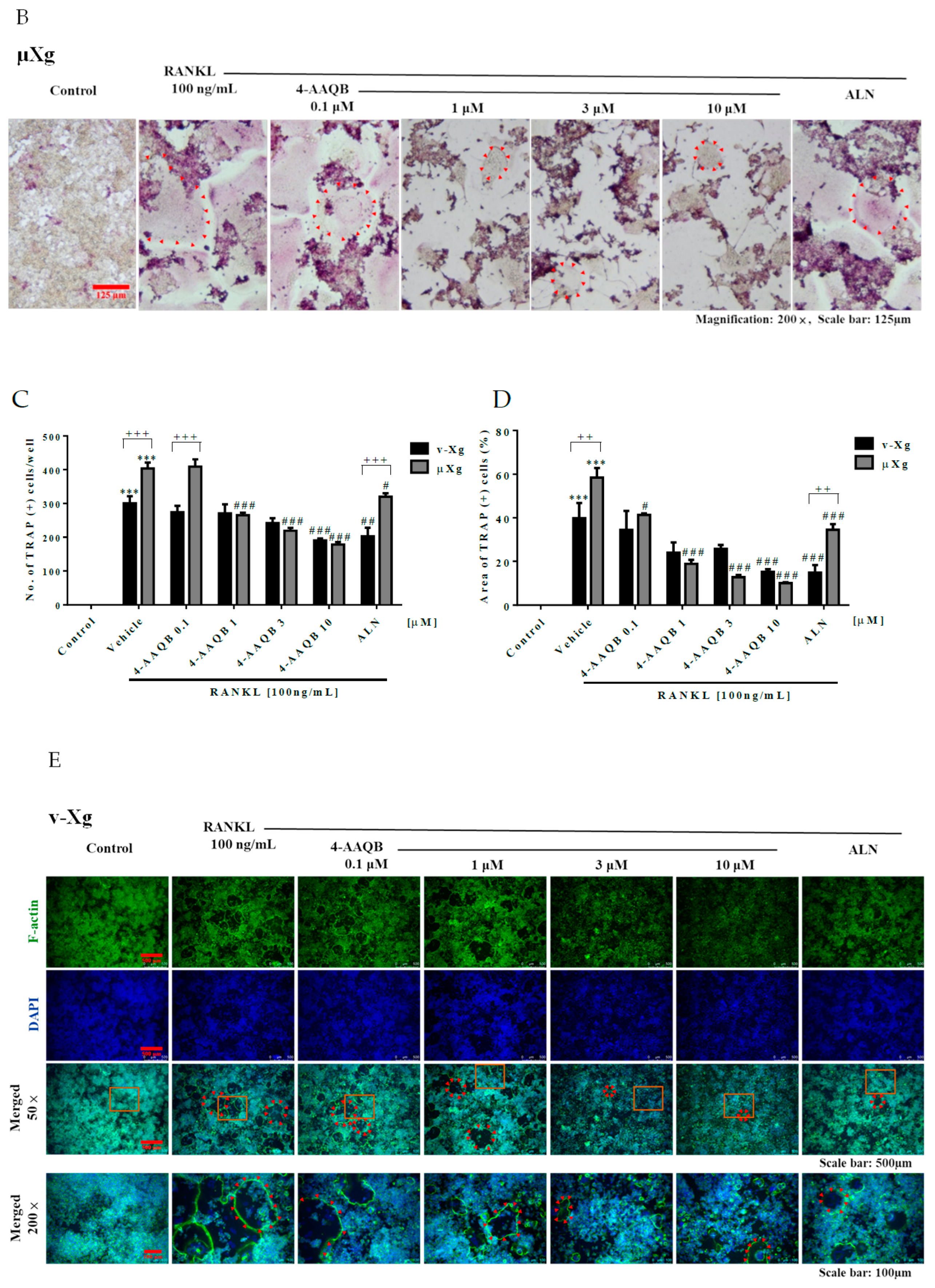
2.5. Tartrate-Resistant Acid Phosphatase (Trap) Staining
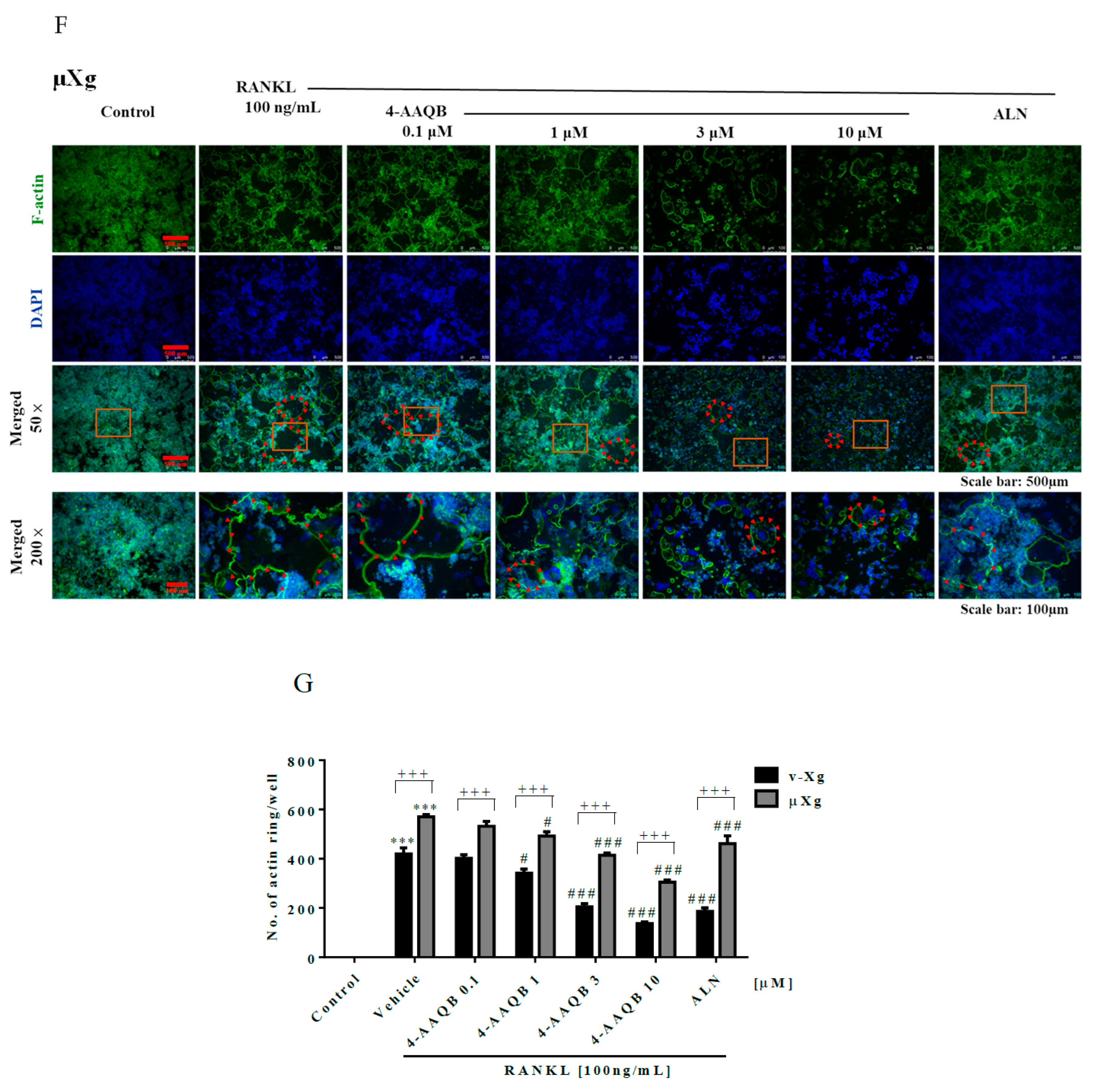
2.6. Actin Ring Immunofluorescence
2.7. Osteo Assay
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. 4-AAQB Cytotoxicity under Both v-Xg And µXg Conditions
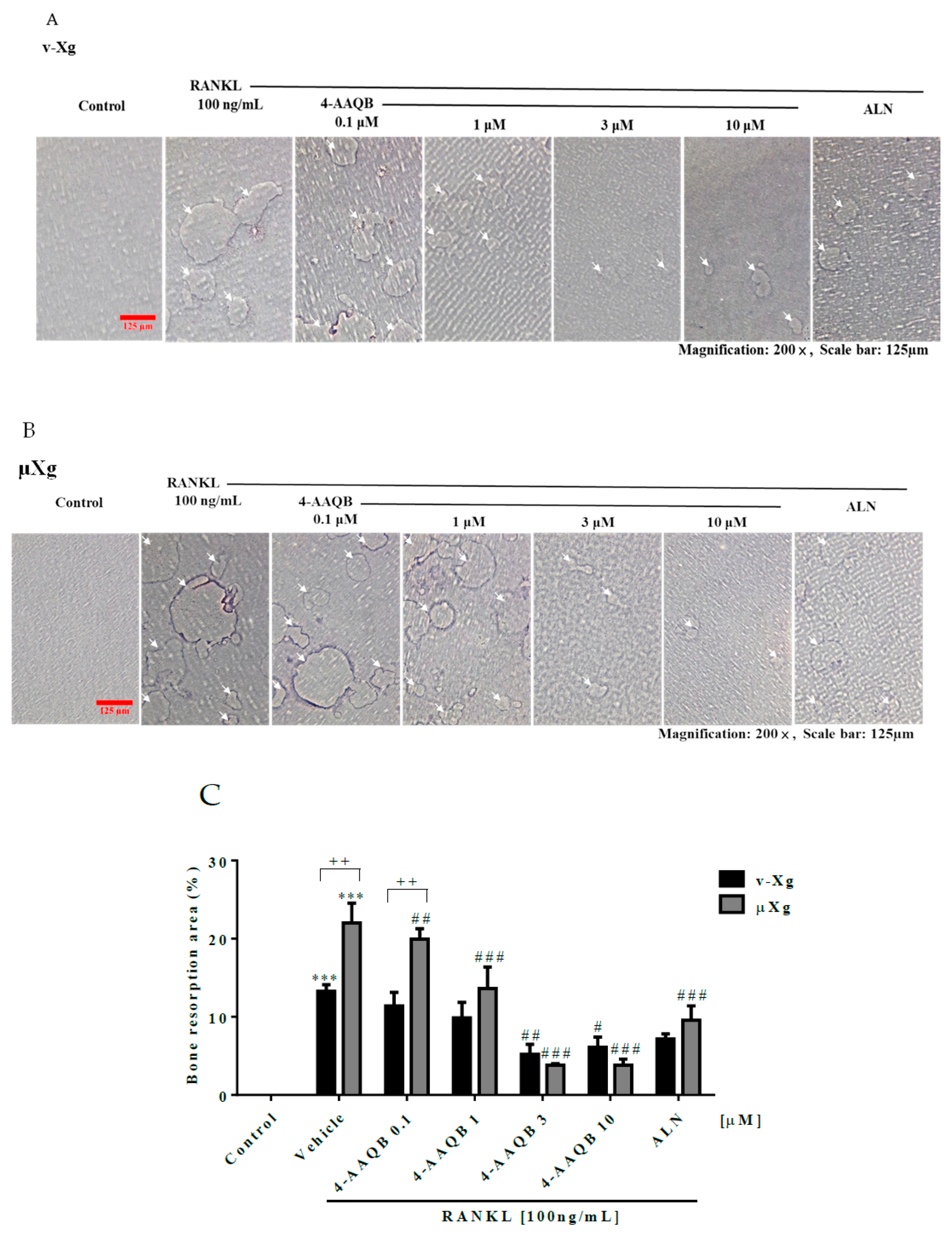
3.2. 4-AAQB Effects on RANKL-Induced Osteoclast Formation and Resorption Ability in Both v-Xg and µXg Conditions
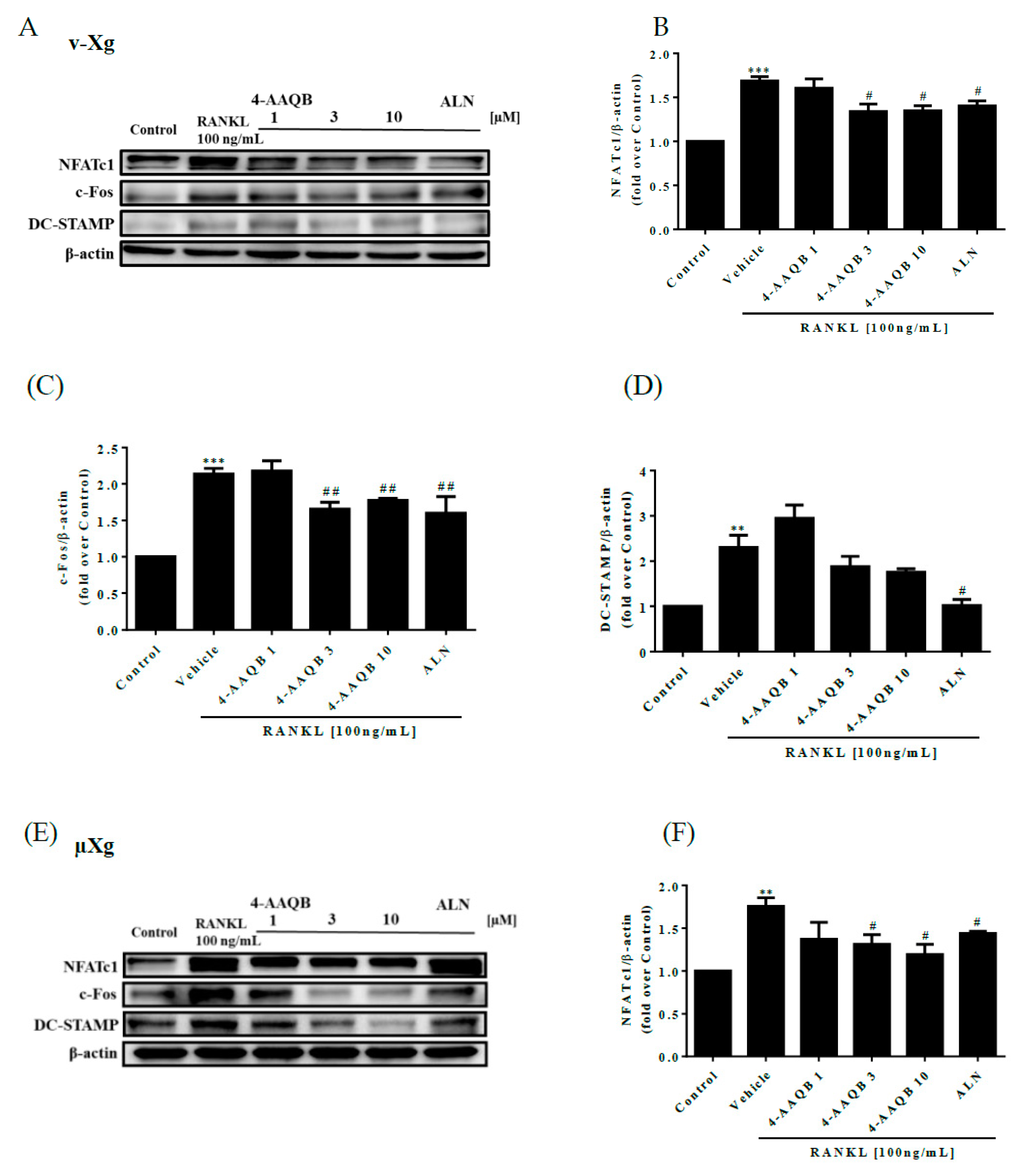
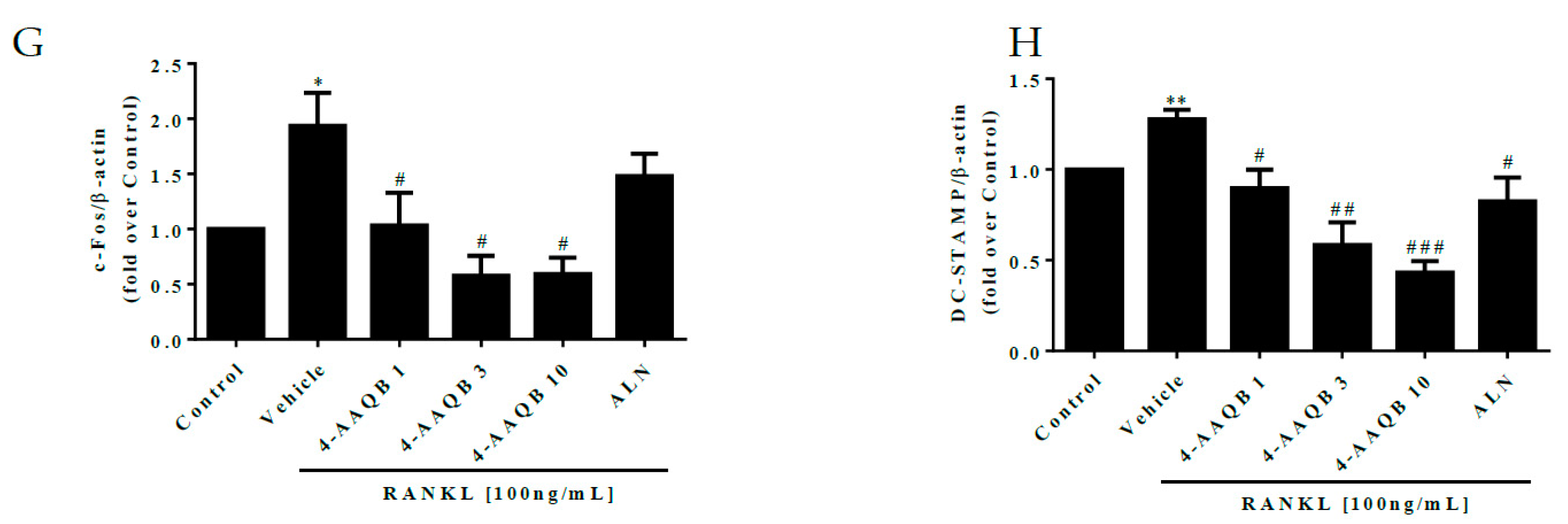
3.3. Inhibition of Essential Osteoclast Differentiation Pathways by 4-AAQB, Especially in µXg Conditions
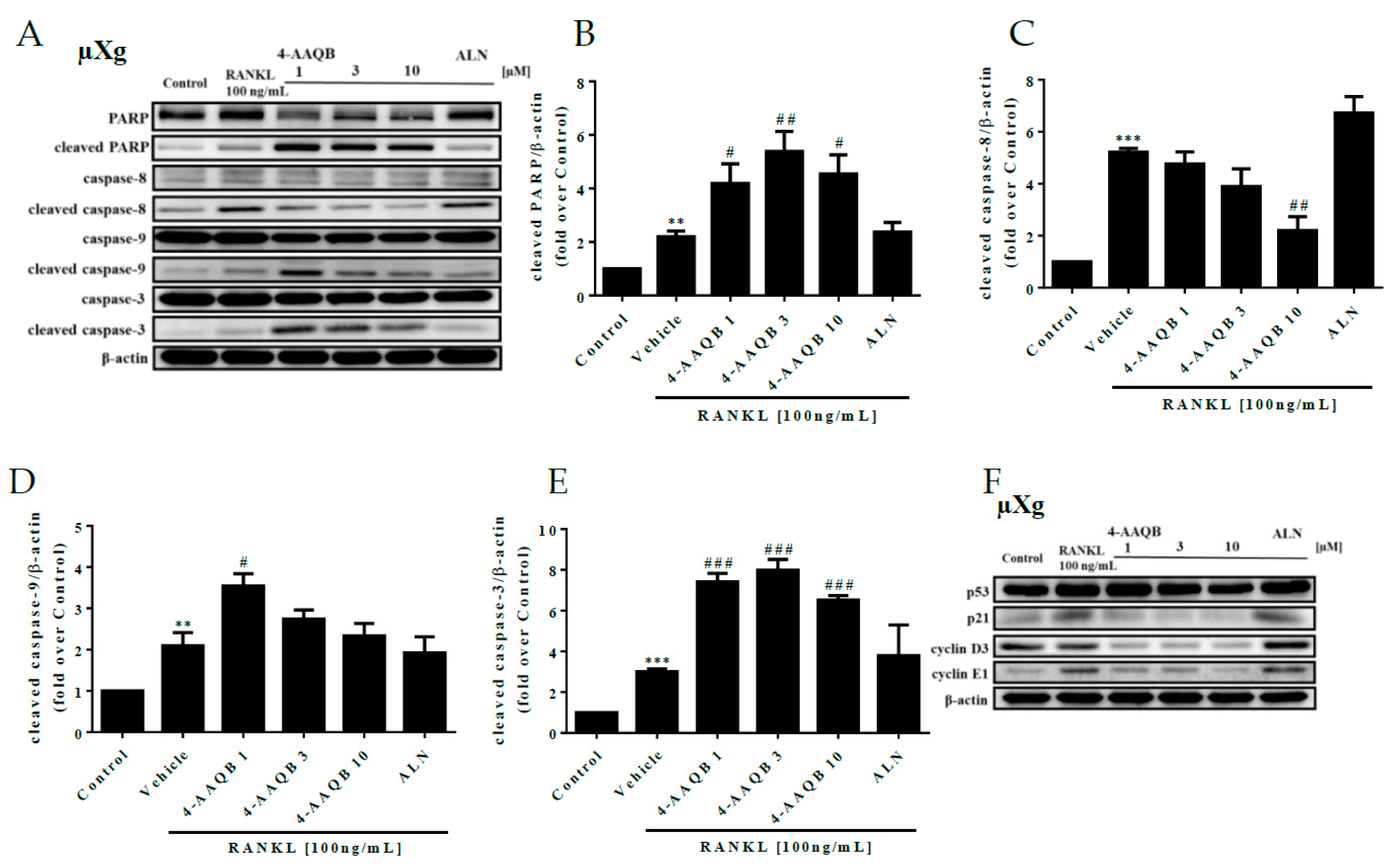
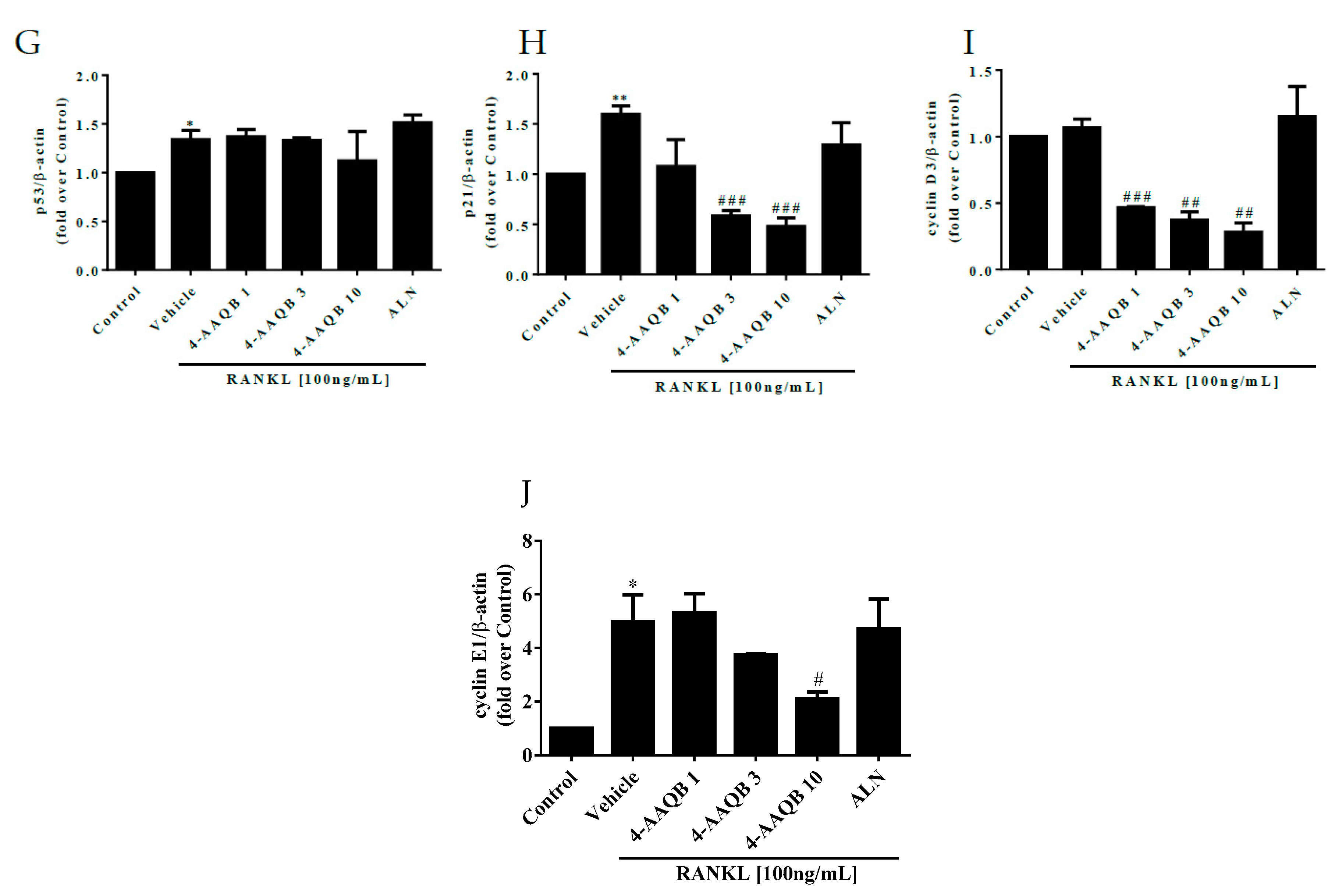
3.4. 4-AAQB Effects on Osteoclast Apoptosis and Cell Cycle Arrest under μXg Conditions
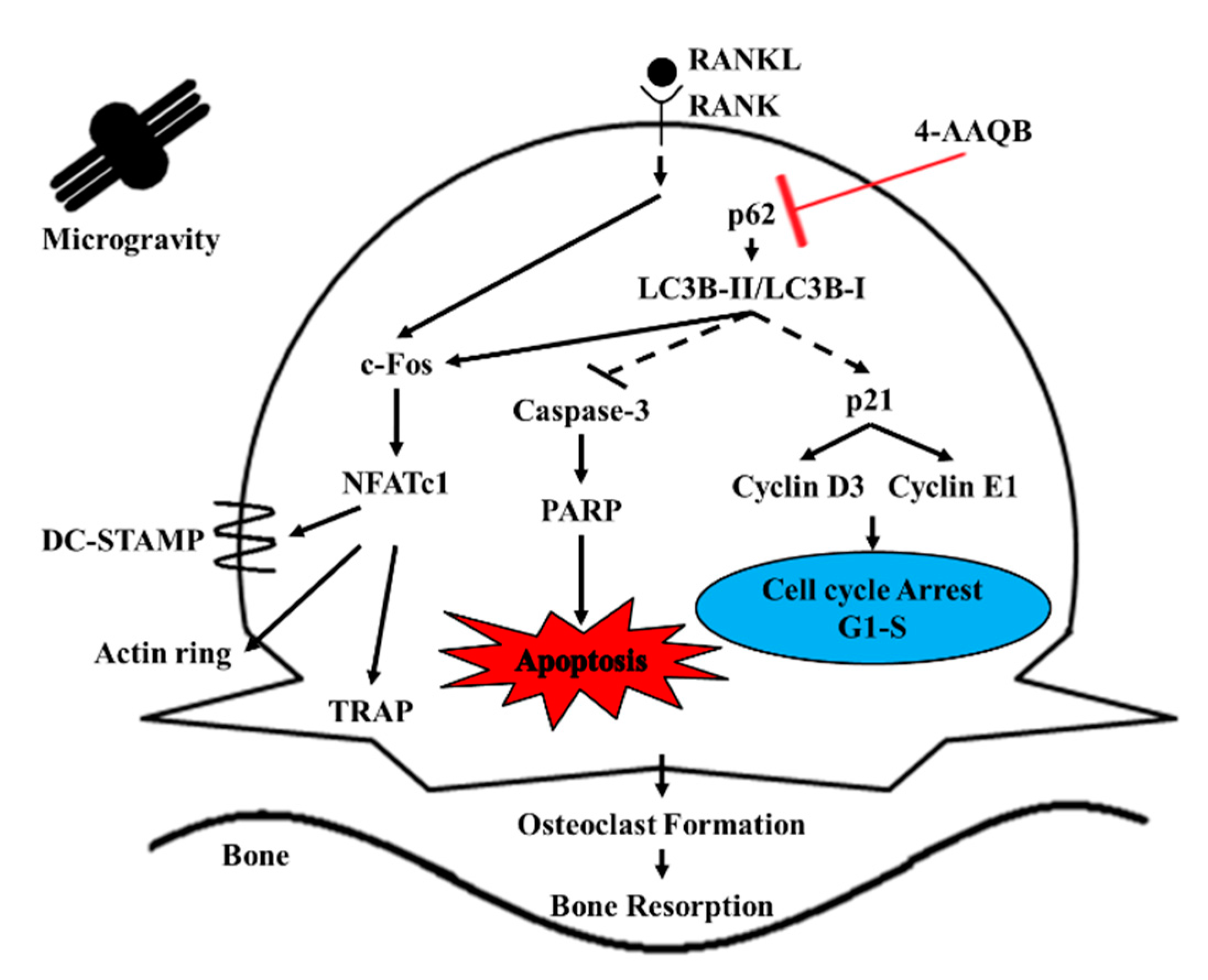
3.5. 4-AAQB Inhibition of RANKL-induced Osteoclastogenesis through an Autophagy-Dependent Pathway Under μXg Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blaber, E.; Marcal, H.; Burns, B.P. Bioastronautics: The influence of microgravity on astronaut health. Astrobiology 2010, 10, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Charles, C.; Christophe, D.; Sandrine, S.; Pierre, M.; Lapiere, C.M.; Betty, M. Gravitational and Space Biology Bulletin; American Society for Gravitational and Space Biology: New Orleans, LA, USA, 2003. [Google Scholar]
- Vernikos, J.; Schneider, V.S. Space, gravity and the physiology of aging: Parallel or convergent disciplines? A mini-review. Gerontology 2010, 56, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Barger, L.K.; Greenleaf, J.E.; Baldini, F.; Huff, D. Effects of space missions on the human immune system: A meta-analysis. Sports Med. Train. Rehabil. 1995, 5, 293–310. [Google Scholar] [CrossRef]
- Dole, N.S. Genetic Determinants of Skeletal Diseases: Role of microRNAs. Ph.D. Thesis, University of Connecticut, Mansfield, CT, USA, 2015. [Google Scholar]
- Lambrecht, G.; Petersen, N.; Weerts, G.; Pruett, C.; Evetts, S.; Stokes, M.; Hides, J. The role of physiotherapy in the European Space Agency strategy for preparation and reconditioning of astronauts before and after long duration space flight. Musculoskelet. Sci. Pract. 2017, 27 (Suppl. 1), S15–S22. [Google Scholar] [CrossRef] [Green Version]
- Ontiveros, C.; McCabe, L.R. Simulated microgravity suppresses osteoblast phenotype, Runx2 levels and AP-1 transactivation. J. Cell. Biochem. 2003, 88, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Ethiraj, P.; Link, J.R.; Sinkway, J.M.; Brown, G.D.; Parler, W.A.; Reddy, S.V. Microgravity modulation of syncytin-A expression enhance osteoclast formation. J. Cell Biochem. 2018, 119, 5696–5703. [Google Scholar] [CrossRef]
- Shanmugarajan, S.; Zhang, Y.; Moreno-Villanueva, M.; Clanton, R.; Rohde, L.H.; Ramesh, G.T.; Sibonga, J.D.; Wu, H. Combined Effects of Simulated Microgravity and Radiation Exposure on Osteoclast Cell Fusion. Int. J. Mol. Sci. 2017, 18, 2443. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.M.; He, L.; Lee, S.; Park, C.; Kim, D.H.; Han, H.J.; Han, J.; Hwang, J.; Cha-Molstad, H.; Lee, K.H.; et al. Inhibition of osteoclasts differentiation by CDC2-induced NFATc1 phosphorylation. Bone 2020, 131, 115153. [Google Scholar] [CrossRef]
- Pang, M.; Rodriguez-Gonzalez, M.; Hernandez, M.; Recinos, C.C.; Seldeen, K.L.; Troen, B.R. AP-1 and Mitf interact with NFATc1 to stimulate cathepsin K promoter activity in osteoclast precursors. J. Cell Biochem. 2019, 120, 12382–12392. [Google Scholar] [CrossRef]
- Choi, Y.; Yoo, J.H.; Lee, J.H.; Lee, Y.; Bae, M.K.; Kim, Y.D.; Kim, H.J. Connective tissue growth factor (CTGF) regulates the fusion of osteoclast precursors by inhibiting Bcl6 in periodontitis. Int. J. Med. Sci. 2020, 17, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.L.; Liu, Y.; Zhou, Y.L.; Li, J.L.; Sun, Q.Y.; Wu, L.X.; Wen, L.; Lai, Y.Y.; Liu, Y.R.; Chang, Y.J.; et al. Osteoclast stimulatory transmembrane protein (OC-STAMP) is a promising molecular prognostic indicator for multiple myeloma. Eur. J. Haematol. 2020, 105, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Nakashima, T.; Hiroshi, N.; Penninger, J.M. RANKL-RANK signaling in osteoclastogenesis and bone disease. Trends Mol. Med. 2006, 12, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Chen, X.; Gao, S.; Yu, X.; Xiao, J.; Zhang, B.; Liu, X.; Dai, M. Key Triggers of Osteoclast-Related Diseases and Available Strategies for Targeted Therapies: A Review. Front. Med. 2017, 4, 234. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, C.; Yi, X.; Liu, H.; Wang, Y. Effects of sodium alendronate on osteoporosis and apoptosis-related factors Cyt C, Apaf-1 and caspase-9. Biomed. Res. 2018, 29, 416–420. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Yoshimura, Y.; Deyama, Y.; Kikuiri, T.; Hasegawa, T.; Tei, K.; Shinoda, H.; Suzuki, K.; Kitagawa, Y. Effects of bisphosphonates on osteoclastogenesis in RAW264.7 cells. Int. J. Mol. Med. 2012, 29, 1007–1015. [Google Scholar] [PubMed] [Green Version]
- Xue, Y.; Liang, Z.; Fu, X.; Wang, T.; Xie, Q.; Ke, D. IL-17A modulates osteoclast precursors’ apoptosis through autophagy-TRAF3 signaling during osteoclastogenesis. Biochem. Biophys. Res. Commun. 2019, 508, 1088–1092. [Google Scholar] [CrossRef]
- Xiu, Y.; Xu, H.; Zhao, C.; Li, J.; Morita, Y.; Yao, Z.; Xing, L.; Boyce, B.F. Chloroquine reduces osteoclastogenesis in murine osteoporosis by preventing TRAF3 degradation. J. Clin. Investig. 2014, 124, 297–310. [Google Scholar] [CrossRef]
- Cavanagh, P.R.; Licata, A.A.; Rice, A.J. Exercise And Pharmacological Countermeasures For Bone Loss During Longduration Space Flight. Gravit. Space Biol. 2005, 18, 39–58. [Google Scholar]
- LeBlanc, A.; Schneider, V. Countermeasures Against Space Flight Related Bone Loss. Acta Astronaut. 1992, 27, 89–92. [Google Scholar] [CrossRef]
- Ikegame, M.; Hattori, A.; Tabata, M.J.; Kitamura, K.I.; Tabuchi, Y.; Furusawa, Y.; Maruyama, Y.; Yamamoto, T.; Sekiguchi, T.; Matsuoka, R.; et al. Melatonin is a potential drug for the prevention of bone loss during space flight. J. Pineal Res. 2019, 67, e12594. [Google Scholar] [CrossRef]
- Chang, C.H.; Hsu, C.C.; Lee, A.S.; Wang, S.W.; Lin, K.T.; Chang, W.L.; Peng, H.C.; Huang, W.C.; Chung, C.H. 4-Acetylantroquinonol B inhibits lipopolysaccharide-induced cytokine release and alleviates sepsis through of MAPK and NFkappaB suppression. BMC Complement. Altern. Med. 2018, 18, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.-Y.; Chiang, B.-H. 4-Acetylantroquinonol B from antrodia cinnamomea enhances immune function of dendritic cells against liver cancer stem cells. Biomed. Pharm. 2019, 109, 2262–2269. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Lin, W.C.; Wang, S.Y.; Lin, L.J.; Yu, B.; Lee, T.T. Evaluation of potential antioxidant and anti-inflammatory effects of Antrodia cinnamomea powder and the underlying molecular mechanisms via Nrf2- and NF-kappaB-dominated pathways in broiler chickens. Poult Sci. 2018, 97, 2419–2434. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Pan, J.H.; Liu, R.H.; Kuo, Y.H.; Sheen, L.Y.; Chiang, B.H. The 4-acetylantroquinonol B isolated from mycelium of Antrodia cinnamomea inhibits proliferation of hepatoma cells. J. Sci. Food Agric. 2010, 90, 1739–1744. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Bamodu, O.A.; Huang, W.C.; Zucha, M.A.; Lin, Y.K.; Wu, A.T.H.; Huang, C.C.; Lee, W.H.; Yuan, C.C.; Hsiao, M.; et al. 4-Acetylantroquinonol B suppresses autophagic flux and improves cisplatin sensitivity in highly aggressive epithelial cancer through the PI3K/Akt/mTOR/p70S6K signaling pathway. Toxicol. Appl. Pharm. 2017, 325, 48–60. [Google Scholar] [CrossRef]
- Zhu, W.; Yin, Z.; Zhang, Q.; Guo, S.; Shen, Y.; Liu, T.; Liu, B.; Wan, L.; Li, S.; Chen, X.; et al. Proanthocyanidins inhibit osteoclast formation and function by inhibiting the NF-kappaB and JNK signaling pathways during osteoporosis treatment. Biochem. Biophys. Res. Commun. 2019, 509, 294–300. [Google Scholar] [CrossRef]
- Yen, I.C.; Lee, S.Y.; Lin, K.T.; Lai, F.Y.; Kuo, M.T.; Chang, W.L. In Vitro Anticancer Activity and Structural Characterization of Ubiquinones from Antrodia cinnamomea Mycelium. Molecules 2017, 22, 747. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.S.; Wang, G.J.; Wang, S.Y.; Lin, Y.Y.; Kuo, Y.H.; Lee, T.H. New constituents with iNOS inhibitory activity from mycelium of Antrodia camphorata. Planta Med. 2009, 75, 512–516. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Lee, K.Y.; Park, B. Icariin abrogates osteoclast formation through the regulation of the RANKL-mediated TRAF6/NF-kappaB/ERK signaling pathway in Raw264.7 cells. Phytomed. Int. J. Phytother. Phytopharm. 2018, 51, 181–190. [Google Scholar]
- Kim, C.J.; Shin, S.H.; Kim, B.J.; Kim, C.H.; Kim, J.H.; Kang, H.M.; Park, B.S.; Kim, I.R. The Effects of Kaempferol-Inhibited Autophagy on Osteoclast Formation. Int. J. Mol. Sci. 2018, 19, 125. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.H.; Shi, L.S.; Chung, M.C.; Chang, T.C.; Lee, S.Y. Antcamphin M Inhibits TLR4-Mediated Inflammatory Responses by Upregulating the Nrf2/HO-1 Pathway and Suppressing the NLRP3 Inflammasome Pathway in Macrophages. Am. J. Chin. Med. 2019, 47, 1611–1626. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Jiang, J.; Lu, X.; Zhang, T.; Zhao, K.; Han, W.; Yang, W.; Qian, Y. Garcinol suppresses RANKL-induced osteoclastogenesis and its underlying mechanism. J. Cell Physiol. 2019, 234, 7498–7509. [Google Scholar] [CrossRef] [PubMed]
- Chaugule, S.; Sureshbabu, S.K.; Dakave, S.; Krishna, C.M.; Chaudhari, P.; Indap, M.; Chiplunkar, S. Hexane Fraction of Turbo brunneus Inhibits Intermediates of RANK-RANKL Signaling Pathway and Prevent Ovariectomy Induced Bone Loss. Front. Med. 2019, 10, 608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, H.; Tan, J.; Faruqi, F.; Beltzer, J. Available online: https://www.corning.com/catalog/cls/documents/application-notes/snappshots_CLS_AN_144_osteo_assay.pdf (accessed on 22 September 2020).
- Yen, I.C.; Shi, L.S.; Chung, M.C.; Ahmetaj-Shala, B.; Chang, T.C.; Lee, S.Y. Antrolone, a Novel Benzoid Derived from Antrodia cinnamomea, Inhibits the LPS-Induced Inflammatory Response in RAW264.7 Macrophage Cells by Balancing the NF-[Formula: See text]B and Nrf2 Pathways. Am. J. Chin. Med. 2018, 46, 1297–1313. [Google Scholar] [CrossRef]
- Chatani, M.; Mantoku, A.; Takeyama, K.; Abduweli, D.; Sugamori, Y.; Aoki, K.; Ohya, K.; Suzuki, H.; Uchida, S.; Sakimura, T.; et al. Microgravity promotes osteoclast activity in medaka fish reared at the international space station. Sci. Rep. 2015, 5, 14172. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, N.; Khandani, A.; Camirand, A.; Harrison, R.E. Effects of microgravity on osteoclast bone resorption and osteoblast cytoskeletal organization and adhesion. Bone 2011, 49, 965–974. [Google Scholar] [CrossRef]
- Chou, K.C.; Yang, S.H.; Wu, H.L.; Lin, P.Y.; Chang, T.L.; Sheu, F.; Chen, K.H.; Chiang, B.H. Biosynthesis of Antroquinonol and 4-Acetylantroquinonol B via a Polyketide Pathway Using Orsellinic Acid as a Ring Precursor in Antrodia cinnamomea. J. Agric. Food Chem. 2017, 65, 74–86. [Google Scholar] [CrossRef]
- Moon, H.J.; Ko, W.K.; Jung, M.S.; Kim, J.H.; Lee, W.J.; Park, K.S.; Heo, J.K.; Bang, J.B.; Kwon, I.K. Coenzyme q10 regulates osteoclast and osteoblast differentiation. J. Food Sci. 2013, 78, H785–H891. [Google Scholar] [CrossRef]
- Zheng, D.; Cui, C.; Yu, M.; Li, X.; Wang, L.; Chen, X.; Lin, Y. Coenzyme Q10 promotes osteoblast proliferation and differentiation and protects against ovariectomy-induced osteoporosis. Mol. Med. Rep. 2018, 17, 400–407. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M. Bones protected by promotion of osteoclast apoptosis. Lancet 1996, 348, 9033. [Google Scholar] [CrossRef]
- Liu, W.; Xu, C.; Zhao, H.; Xia, P.; Song, R.; Gu, J.; Liu, X.; Bian, J.; Yuan, Y.; Liu, Z. Osteoprotegerin Induces Apoptosis of Osteoclasts and Osteoclast Precursor Cells via the Fas/Fas Ligand Pathway. PLoS ONE 2015, 10, e0142519. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.W.; Elledge, S.J.; Keyomarsi, K.; Dynlacht, B.; Tsai, L.H.; Zhang, P.; Dobrowolski, S.; Bai, C.; Connell-Crowley, L.; Swindell, E. Inhibition of Cyclin-dependent Kinases by p21. Mol. Biol. Cell. 1995, 6, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossy, E.; Talmant, M.; Peyrin, F.; Akrout, L.; Cloetens, P.; Laugier, P. An in vitro study of the ultrasonic axial transmission technique at the radius: 1-MHz velocity measurements are sensitive to both mineralization and intracortical porosity. J. Bone Miner. Res. 2004, 19, 1548–1556. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Liu, C.; Yao, L.; Bai, Y.; Cheng, P.; Li, Z.; Luo, K.; Mei, T.; Li, J.; Xing, J.; et al. Alendronate induces osteoclast precursor apoptosis via peroxisomal dysfunction mediated ER stress. J. Cell Physiol. 2018, 233, 7415–7423. [Google Scholar] [CrossRef] [PubMed]
- Kusamori, K.; Katsumi, H.; Abe, M.; Ueda, A.; Sakai, R.; Hayashi, R.; Hirai, Y.; Quan, Y.S.; Kamiyama, F.; Sakane, T.; et al. Development of a novel transdermal patch of alendronate, a nitrogen-containing bisphosphonate, for the treatment of osteoporosis. J. Bone Miner. Res. 2010, 25, 2582–2591. [Google Scholar] [CrossRef]
- Ryu, H.W.; Choi, S.H.; Namkoong, S.; Jang, I.S.; Seo, D.H.; Choi, I.; Kim, H.S.; Park, J. Simulated microgravity contributes to autophagy induction by regulating AMP-activated protein kinase. DNA Cell Biol. 2014, 33, 128–135. [Google Scholar] [CrossRef]
- Shapiro, I.M.; Layfield, R.; Lotz, M.; Settembre, C.; Whitehouse, C. Boning up on autophagy: The role of autophagy in skeletal biology. Autophagy 2014, 10, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Zhang, A.; Cai, J.; Yuan, N.; Lin, W.; Liu, S.; Xu, F.; Song, L.; Li, X.; Fang, Y.; et al. Autophagy regulates the cell cycle of murine HSPCs in a nutrient-dependent manner. Exp. Hematol. 2015, 43, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.P.; Chien, J.Y.; Tsai, R.K. Ethambutol induces impaired autophagic flux and apoptosis in the rat retina. Dis Model. Mech. 2015, 8, 977–987. [Google Scholar] [CrossRef] [Green Version]
- Zheng, K.; He, Z.; Kitazato, K.; Wang, Y. Selective Autophagy Regulates Cell Cycle in Cancer Therapy. Theranostics 2019, 9, 104–125. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Kim, J.M.; Lee, K.; Park, S.Y.; Lim, H.S.; Kim, T.; Jeong, D. Synchronized Cell Cycle Arrest Promotes Osteoclast Differentiation. Int J. Mol. Sci. 2016, 17, 1292. [Google Scholar] [CrossRef] [PubMed]












© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.-H.; Ou, C.-H.; Yen, I.-C.; Lee, S.-Y. 4-Acetylantroquinonol B Inhibits Osteoclastogenesis by Inhibiting the Autophagy Pathway in a Simulated Microgravity Model. Int. J. Mol. Sci. 2020, 21, 6971. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186971
Wu C-H, Ou C-H, Yen I-C, Lee S-Y. 4-Acetylantroquinonol B Inhibits Osteoclastogenesis by Inhibiting the Autophagy Pathway in a Simulated Microgravity Model. International Journal of Molecular Sciences. 2020; 21(18):6971. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186971
Chicago/Turabian StyleWu, Chia-Hsin, Ching-Huei Ou, I-Chuan Yen, and Shih-Yu Lee. 2020. "4-Acetylantroquinonol B Inhibits Osteoclastogenesis by Inhibiting the Autophagy Pathway in a Simulated Microgravity Model" International Journal of Molecular Sciences 21, no. 18: 6971. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186971