SOX9 Knockout Induces Polyploidy and Changes Sensitivity to Tumor Treatment Strategies in a Chondrosarcoma Cell Line

 ,
, 
Abstract
:1. Introduction
2. Results
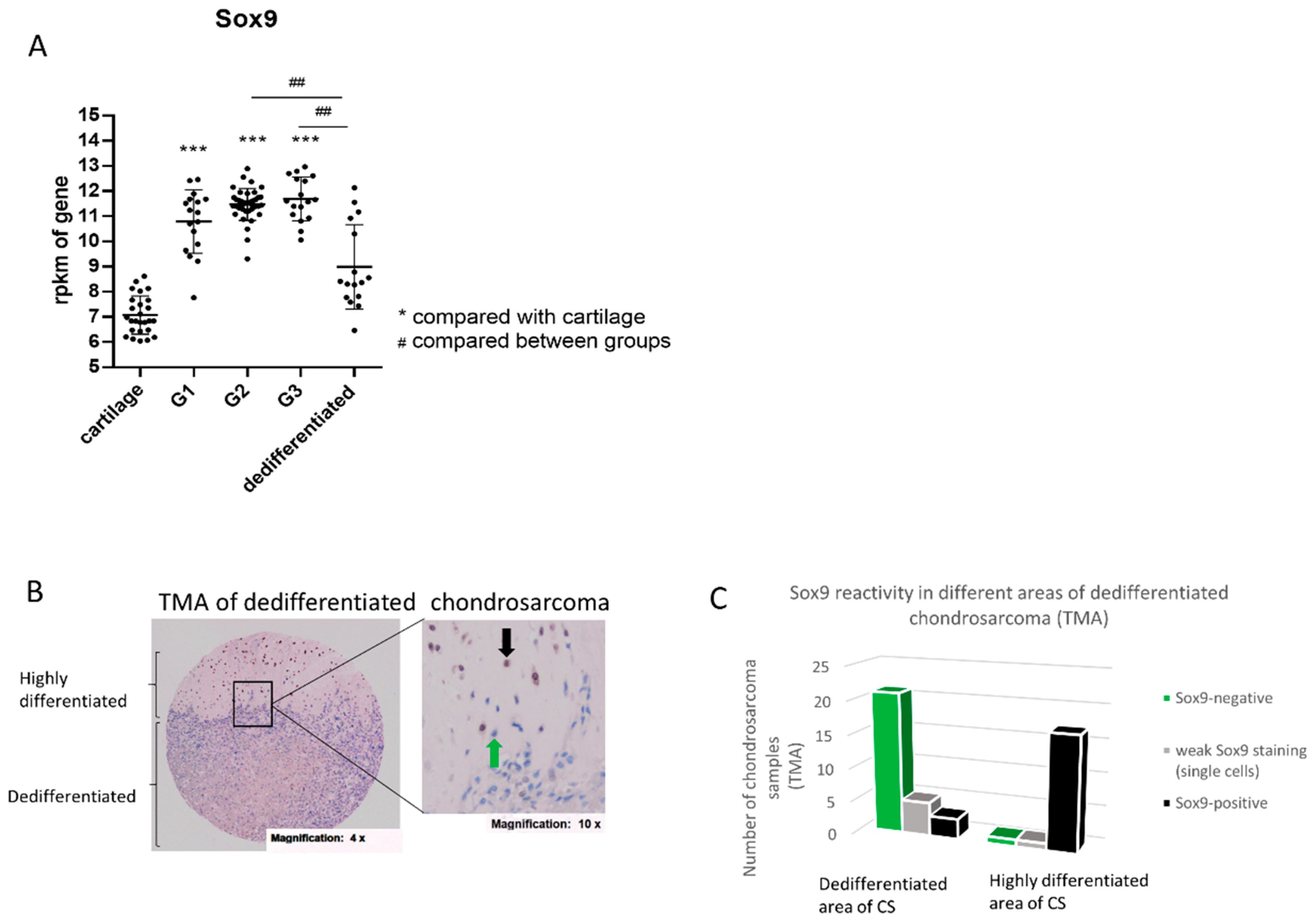
2.1. Increased SOX9 Gene Expression in G1, G2 and G3 Chondrosarcoma, but Not in Dedifferentiated Chondrosarcoma
2.2. SOX9 Protein Expression in Dedifferentiated Chondrosarcoma
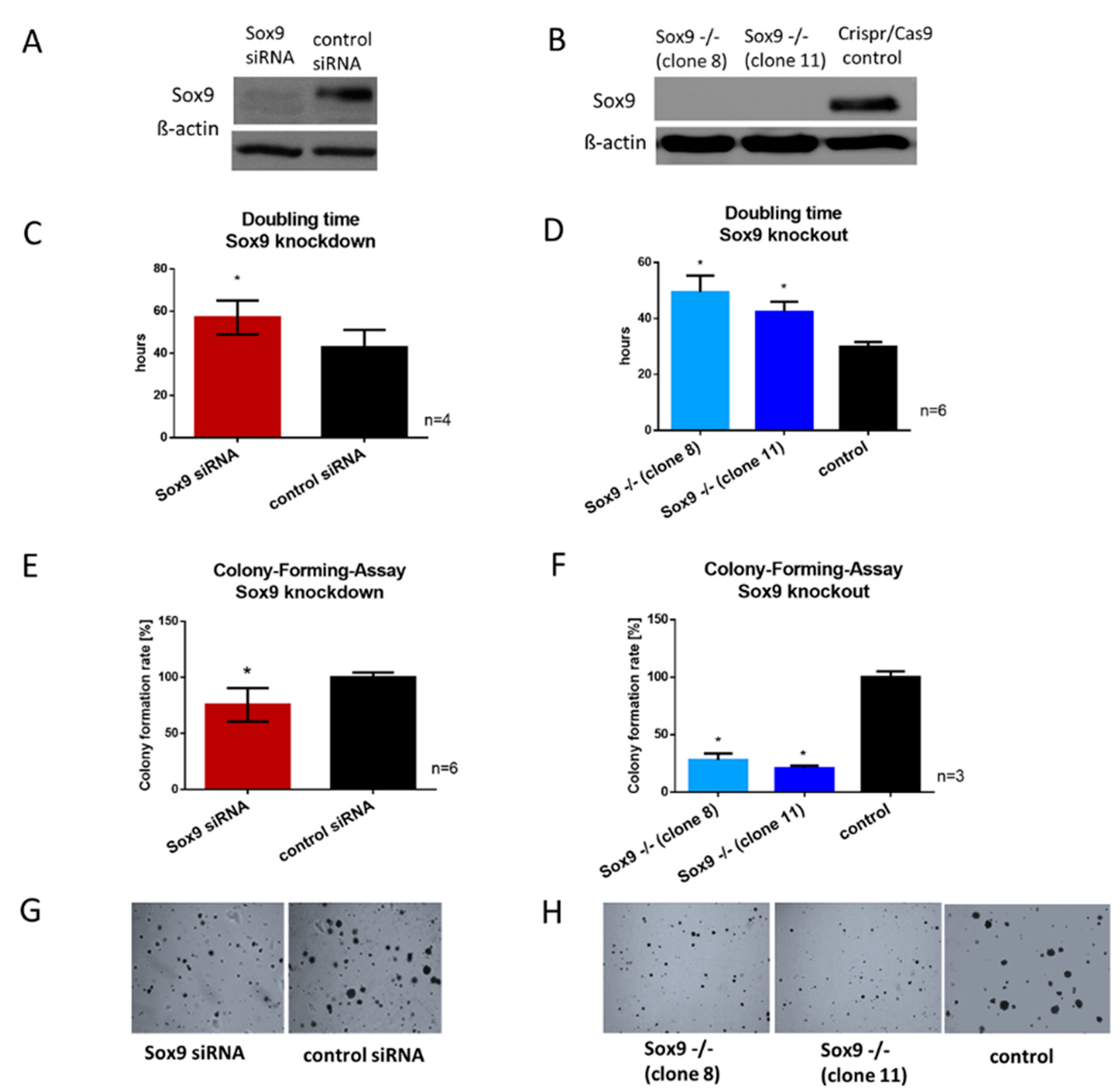
2.3. Generation of siRNA-Mediated SOX9 Knockdown and Crispr/Cas9-Mediated SOX9 Knockout Clones
2.4. SOX9 Knockdown and Knockout Impair Proliferation and the Ability to Form Colonies
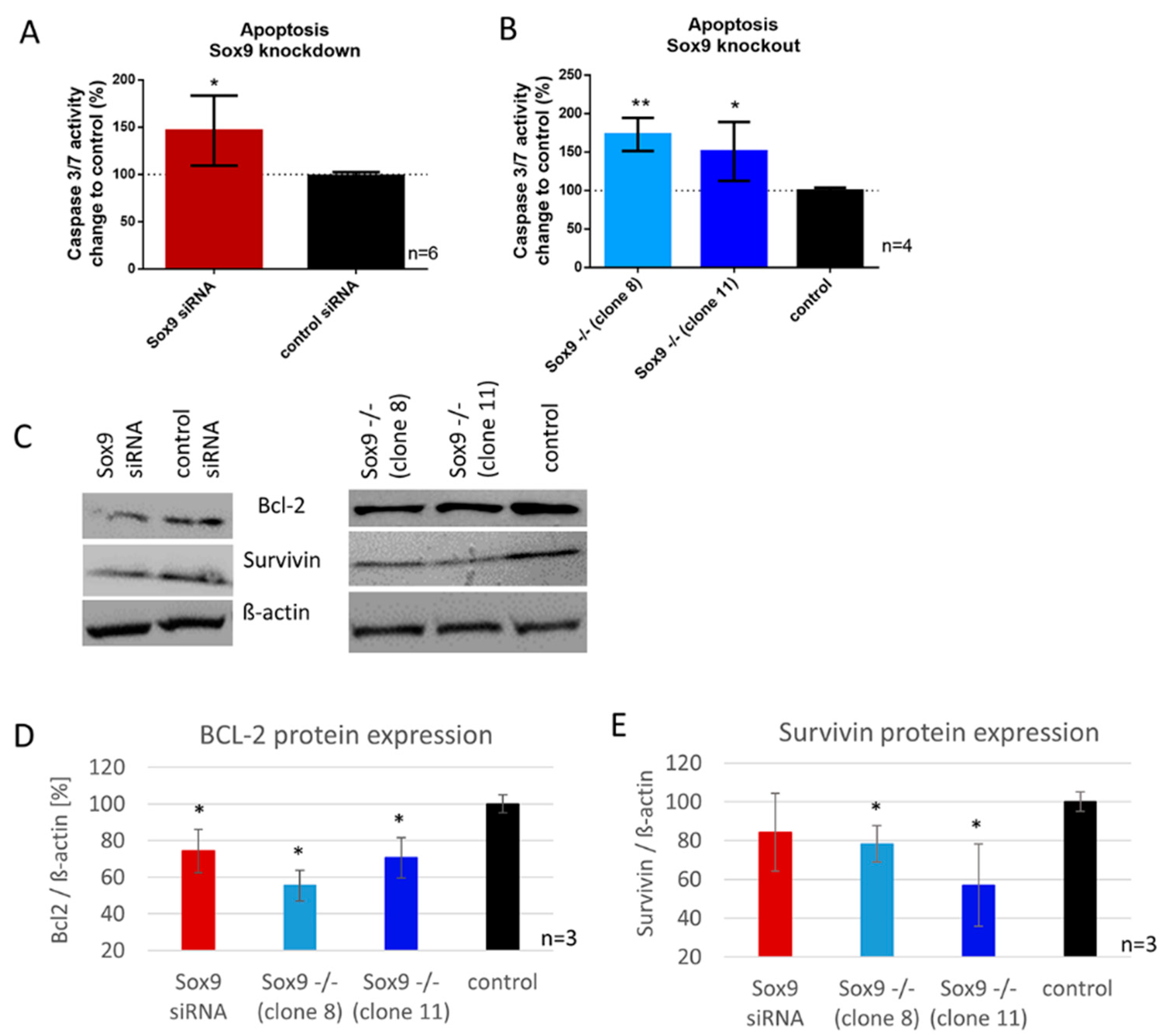
2.5. SOX9 Knockdown and Knockout Increases Apoptosis in Chondrosarcoma Cells Involving BCL-2 and Survivin Expression
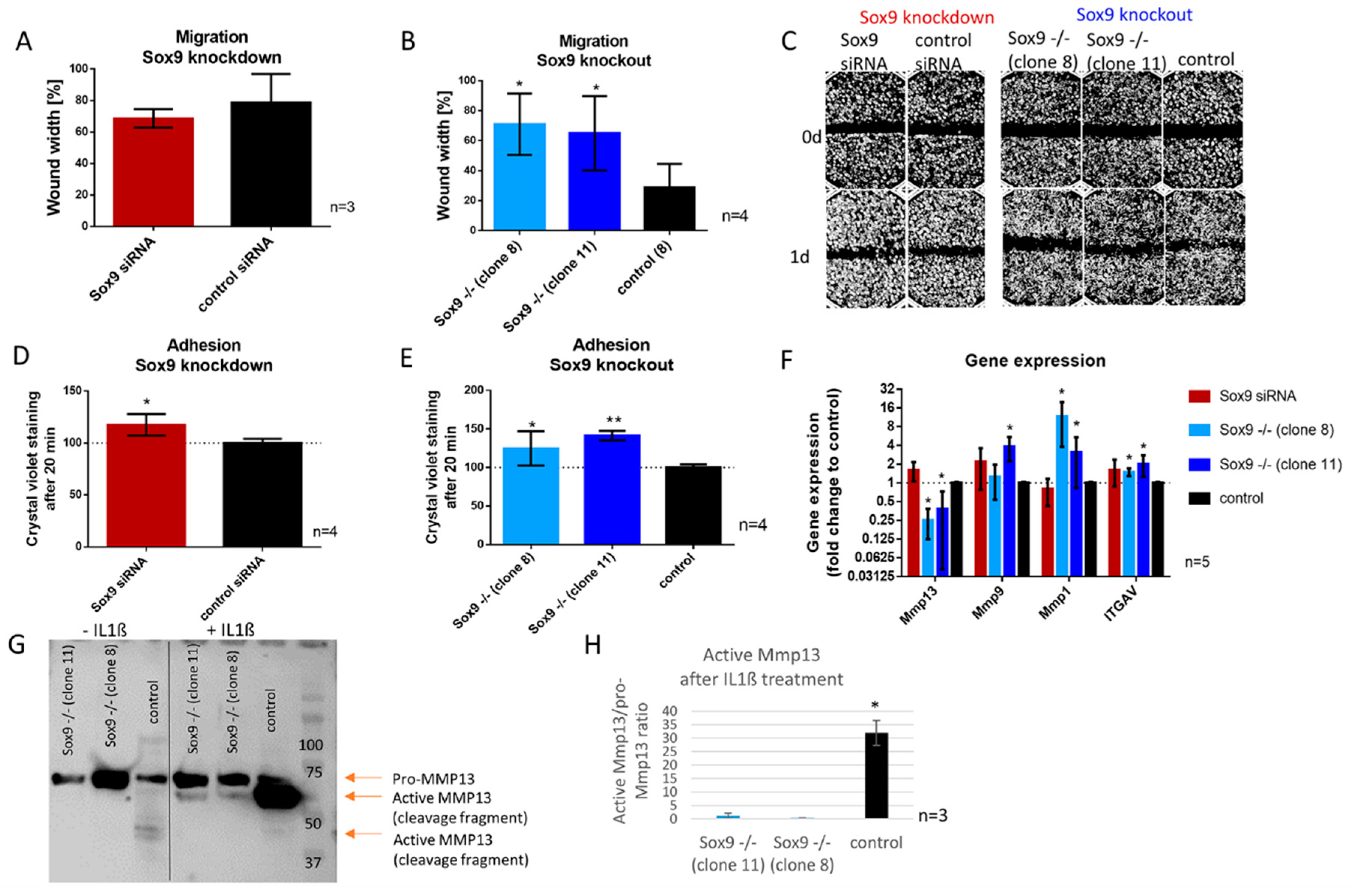
2.6. SOX9 Knockout Increases Cell Adhesion and Decreases Cell Migration Accompanied by the Inability to Activate MMP13
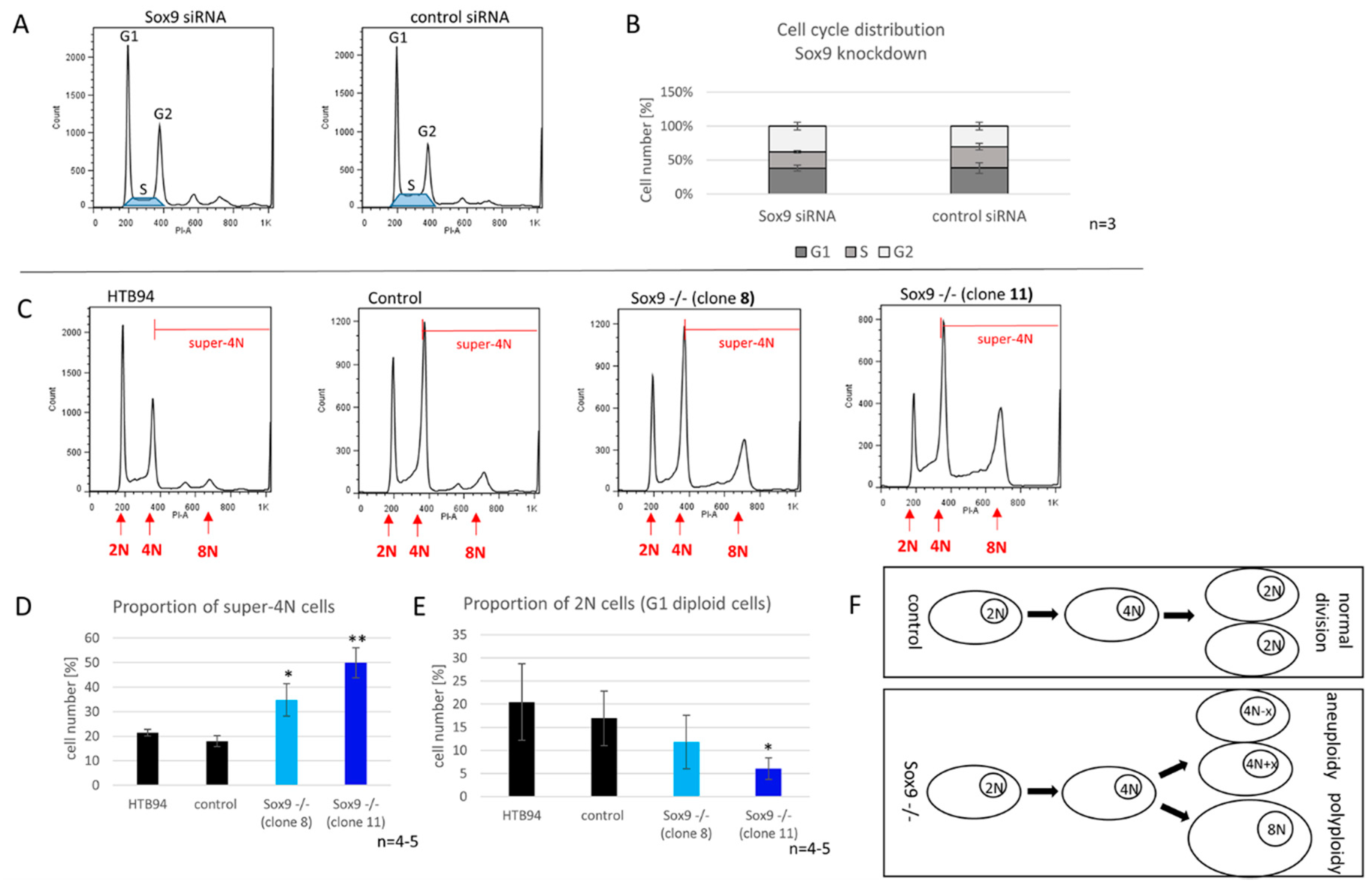
2.7. SOX9 Depletion Induces Polyploidy
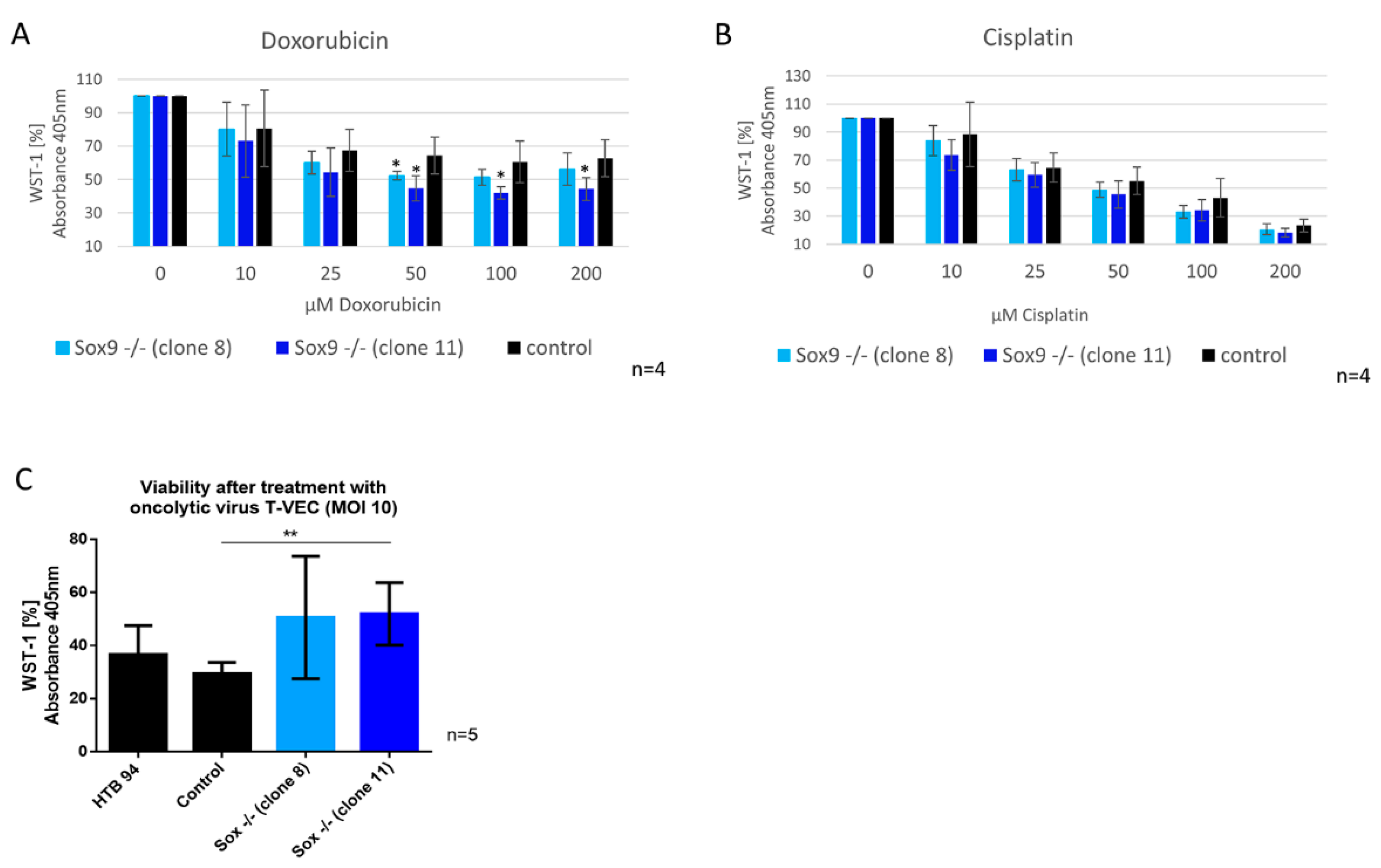
2.8. SOX9 Depletion Alters Sensitivity against Doxorubicin and against Infection with Oncolytic Virus (T-VEC)
3. Discussion
4. Material and Methods
4.1. Meta-Analysis of SOX9 Gene Expression Level
4.2. Cell Culture
4.3. Immunohistochemistry
4.4. TMA Generation
4.5. SOX9 siRNA Transfection
4.6. Generation of SOX9−/− Knockout Cells
4.7. Protein Extraction and Western Blot Analysis
4.8. Flow Cytometry
4.9. RNA Isolation from Human Tissue Samples (Chondrosarcoma Biopsies)
4.10. RNA Isolation and Real-Time RT-PCR from HTB94 Cells
4.11. Doubling Time
4.12. Caspase-3/7 Assay
4.13. Wound Healing
4.14. Colony-Forming Assay
4.15. WST-Assay
4.16. Oncolytic Virus (T-VEC) Infection
4.17. Statistics
4.18. Ethical Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Evola, F.R.; Costarella, L.; Pavone, V.; Caff, G.; Cannavo, L.; Sessa, A.; Avondo, S.; Sessa, G. Biomarkers of Osteosarcoma, Chondrosarcoma, and Ewing Sarcoma. Front. Pharmacol. 2017, 8, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesolowski, R.; Budd, G.T. Use of chemotherapy for patients with bone and soft-tissue sarcomas. Clevel. Clin. J. Med. 2010, 77 (Suppl. 1), S23–S26. [Google Scholar] [CrossRef]
- Onishi, A.C.; Hincker, A.M.; Lee, F.Y. Surmounting chemotherapy and radioresistance in chondrosarcoma: Molecular mechanisms and therapeutic targets. Sarcoma 2011, 2011, 381564. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, H.; Hogendoorn, P.C.; Dijkstra, S.D.; van Rijswijk, C.S.; Krol, A.D.; Taminiau, A.H.; Bovee, J.V. The clinical approach towards chondrosarcoma. Oncology 2008, 13, 320–329. [Google Scholar] [CrossRef]
- Evans, H.L.; Ayala, A.G.; Romsdahl, M.M. Prognostic factors in chondrosarcoma of bone: A clinicopathologic analysis with emphasis on histologic grading. Cancer 1977, 40, 818–831. [Google Scholar] [CrossRef]
- Kreicbergs, A.; Zetterberg, A.; Soderberg, G. The prognostic significance of nuclear DNA content in chondrosarcoma. Anal. Quant. Cytol. 1980, 2, 272–279. [Google Scholar]
- Grimer, R.J.; Gosheger, G.; Taminiau, A.; Biau, D.; Matejovsky, Z.; Kollender, Y.; San-Julian, M.; Gherlinzoni, F.; Ferrari, C. Dedifferentiated chondrosarcoma: Prognostic factors and outcome from a European group. Eur. J. Cancer 2007, 43, 2060–2065. [Google Scholar] [CrossRef]
- Mercuri, M.; Campanacci, L. Dedifferentiated chondrosarcoma. Skelet. Radiol. 1995, 24, 409–416. [Google Scholar] [CrossRef]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; de Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [CrossRef]
- Wang, H.Y.; Lian, P.; Zheng, P.S. SOX9, a potential tumor suppressor in cervical cancer, transactivates p21WAF1/CIP1 and suppresses cervical tumor growth. Oncotarget 2015, 6, 20711–20722. [Google Scholar] [CrossRef] [Green Version]
- Prevostel, C.; Rammah-Bouazza, C.; Trauchessec, H.; Canterel-Thouennon, L.; Busson, M.; Ychou, M.; Blache, P. SOX9 is an atypical intestinal tumor suppressor controlling the oncogenic Wnt/ss-catenin signaling. Oncotarget 2016, 7, 82228–82243. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, M.; Nakatani, F.; Matsunaga, H.; Seki, T.; Endo, M.; Ogawara, Y.; Machida, Y.; Katsumoto, T.; Yamagata, K.; Hattori, A.; et al. Selective inhibition of mutant IDH1 by DS-1001b ameliorates aberrant histone modifications and impairs tumor activity in chondrosarcoma. Oncogene 2019, 38, 6835–6849. [Google Scholar] [CrossRef] [Green Version]
- Jo, A.; Denduluri, S.; Zhang, B.; Wang, Z.; Yin, L.; Yan, Z.; Kang, R.; Shi, L.L.; Mok, J.; Lee, M.J.; et al. The versatile functions of Sox9 in development, stem cells, and human diseases. Genes Dis. 2014, 1, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Matheu, A.; Collado, M.; Wise, C.; Manterola, L.; Cekaite, L.; Tye, A.J.; Canamero, M.; Bujanda, L.; Schedl, A.; Cheah, K.S.; et al. Oncogenicity of the developmental transcription factor Sox9. Cancer Res. 2012, 72, 1301–1315. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Medina, M.; Avendano-Felix, M.; Lizarraga-Verdugo, E.; Bermudez, M.; Romero-Quintana, J.G.; Ramos-Payan, R.; Ruiz-Garcia, E.; Lopez-Camarillo, C. SOX9 Stem-Cell Factor: Clinical and Functional Relevance in Cancer. J. Oncol. 2019, 2019, 6754040. [Google Scholar] [CrossRef] [Green Version]
- Mak, I.W.; Singh, S.; Turcotte, R.; Ghert, M. The epigenetic regulation of SOX9 by miR-145 in human chondrosarcoma. J. Cell. Biochem. 2015, 116, 37–44. [Google Scholar] [CrossRef]
- Tang, X.; Lu, X.; Guo, W.; Ren, T.; Zhao, H.; Zhao, F.; Tang, G. Different expression of Sox9 and Runx2 between chondrosarcoma and dedifferentiated chondrosarcoma cell line. Eur. J. Cancer Prev. Off. J. Eur. Cancer Prev. Organ. 2010, 19, 466–471. [Google Scholar] [CrossRef]
- Wehrli, B.M.; Huang, W.; De Crombrugghe, B.; Ayala, A.G.; Czerniak, B. Sox9, a master regulator of chondrogenesis, distinguishes mesenchymal chondrosarcoma from other small blue round cell tumors. Hum. Pathol. 2003, 34, 263–269. [Google Scholar] [CrossRef]
- Bovee, J.V.M.G.; Cleton-Jansen, A.M.; Rosenberg, C.; Taminiau, A.H.M.; Cornelisse, C.J.; Hogendoorn, P.C.W. Molecular genetic characterization of both components of a dedifferentiated chondrosarcoma, with implications for its histogenesis. J Pathol 1999, 189, 454–462. [Google Scholar] [CrossRef] [Green Version]
- Ropke, M.; Boltze, C.; Neumann, H.W.; Roessner, A.; Schneider-Stock, R. Genetic and epigenetic alterations in tumor progression in a dedifferentiated chondrosarcoma. Pathol Res. Pract. 2003, 199, 437–444. [Google Scholar] [CrossRef]
- Grote, H.J.; Schneider-Stock, R.; Neumann, W.; Roessner, A. Mutation of p53 with loss of heterozygosity in the osteosarcomatous component of a dedifferentiated chondrosarcoma. Virchows Arch. Int. J. Pathol. 2000, 436, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, B.; Feliz, A.; Ishida, T.; Czerniak, B.; Dorfman, H.D. p53 expression and DNA ploidy of cartilage lesions. Hum. Pathol. 1995, 26, 620–624. [Google Scholar] [CrossRef]
- Bovee, J.V.; Cleton-Jansen, A.M.; Kuipers-Dijkshoorn, N.J.; van den Broek, L.J.; Taminiau, A.H.; Cornelisse, C.J.; Hogendoorn, P.C. Loss of heterozygosity and DNA ploidy point to a diverging genetic mechanism in the origin of peripheral and central chondrosarcoma. Genes Chromosomes Cancer 1999, 26, 237–246. [Google Scholar] [CrossRef]
- Papachristou, D.J.; Goodman, M.A.; Cieply, K.; Hunt, J.L.; Rao, U.N. Comparison of allelic losses in chondroblastoma and primary chondrosarcoma of bone and correlation with fluorescence in situ hybridization analysis. Hum. Pathol. 2006, 37, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Minn, A.J.; Boise, L.H.; Thompson, C.B. Expression of Bcl-xL and loss of p53 can cooperate to overcome a cell cycle checkpoint induced by mitotic spindle damage. Genes Dev. 1996, 10, 2621–2631. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, D.W.; Mao, Y.; Sullivan, K.F. Centromeres and kinetochores: From epigenetics to mitotic checkpoint signaling. Cell 2003, 112, 407–421. [Google Scholar] [CrossRef] [Green Version]
- Storchova, Z.; Pellman, D. From polyploidy to aneuploidy, genome instability and cancer. Nat. Rev. Mol. Cell Biol. 2004, 5, 45–54. [Google Scholar] [CrossRef]
- Weaver, B.A.; Cleveland, D.W. Aneuploidy: Instigator and inhibitor of tumorigenesis. Cancer Res. 2007, 67, 10103–10105. [Google Scholar] [CrossRef] [Green Version]
- Mosieniak, G.; Sikora, E. Polyploidy: The link between senescence and cancer. Curr. Pharm. Des. 2010, 16, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Garbutt, C.C.; Spentzos, D.; Choy, E.; Hornicek, F.J.; Duan, Z. Expression and Therapeutic Potential of SOX9 in Chordoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 5176–5186. [Google Scholar] [CrossRef] [Green Version]
- Stockl, S.; Bauer, R.J.; Bosserhoff, A.K.; Gottl, C.; Grifka, J.; Grassel, S. Sox9 modulates cell survival and adipogenic differentiation of multipotent adult rat mesenchymal stem cells. J. Cell Sci. 2013, 126, 2890–2902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evdokimova, V.; Tognon, C.; Ng, T.; Sorensen, P.H. Reduced proliferation and enhanced migration: Two sides of the same coin? Molecular mechanisms of metastatic progression by YB-1. Cell Cycle 2009, 8, 2901–2906. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Huang, H.; Shi, G.; Zhao, L.; Li, T.; Zhang, Z.; Liu, R.; Hu, Y.; Liu, H.; Yu, J.; et al. TGF-beta1-SOX9 axis-inducible COL10A1 promotes invasion and metastasis in gastric cancer via epithelial-to-mesenchymal transition. Cell Death Dis. 2018, 9, 849. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liang, R.; Liu, C.; Liu, J.A.; Cheung, M.P.L.; Liu, X.; Man, O.Y.; Guan, X.Y.; Lung, H.L.; Cheung, M. SOX9 is a dose-dependent metastatic fate determinant in melanoma. J. Exp. Clin. Cancer Res. CR 2019, 38, 17. [Google Scholar] [CrossRef] [PubMed]
- Johansson, N.; Airola, K.; Grenman, R.; Kariniemi, A.L.; Saarialho-Kere, U.; Kahari, V.M. Expression of collagenase-3 (matrix metalloproteinase-13) in squamous cell carcinomas of the head and neck. Am. J. Pathol. 1997, 151, 499–508. [Google Scholar]
- Yuan, X.; Li, J.; Coulouarn, C.; Lin, T.; Sulpice, L.; Bergeat, D.; De La Torre, C.; Liebe, R.; Gretz, N.; Ebert, M.P.A.; et al. SOX9 expression decreases survival of patients with intrahepatic cholangiocarcinoma by conferring chemoresistance. Br. J. Cancer 2018, 119, 1358–1366. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhao, W.; Zhang, L.; Wang, L.; Wang, J.; Wan, Z.; Hong, Y.; Yu, L. MALAT1-miR-101-SOX9 feedback loop modulates the chemo-resistance of lung cancer cell to DDP via Wnt signaling pathway. Oncotarget 2017, 8, 94317–94329. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Ma, F.; Hu, C.; Ma, J.A.; Wang, J.; Zhang, Y.; Wu, F.; Hou, T.; Jiang, S.; Wang, Y.; et al. SOX9/miR-130a/CTR1 axis modulates DDP-resistance of cervical cancer cell. Cell Cycle 2018, 17, 448–458. [Google Scholar] [CrossRef]
- Garros-Regulez, L.; Aldaz, P.; Arrizabalaga, O.; Moncho-Amor, V.; Carrasco-Garcia, E.; Manterola, L.; Moreno-Cugnon, L.; Barrena, C.; Villanua, J.; Ruiz, I.; et al. mTOR inhibition decreases SOX2-SOX9 mediated glioma stem cell activity and temozolomide resistance. Expert Opin. Ther. Targets 2016, 20, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Bian, X.; Shim, H. Downregulation of microRNA-206 promotes invasion and angiogenesis of triple negative breast cancer. Biochem. Biophys. Res. Commun. 2016, 477, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Riemenschnitter, C.; Teleki, I.; Tischler, V.; Guo, W.; Varga, Z. Stability and prognostic value of Slug, Sox9 and Sox10 expression in breast cancers treated with neoadjuvant chemotherapy. SpringerPlus 2013, 2, 695. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xue, X.; Hong, H.; Qin, M.; Zhou, J.; Sun, Q.; Liang, H.; Gao, L. Upregulation of microRNA-524-5p enhances the cisplatin sensitivity of gastric cancer cells by modulating proliferation and metastasis via targeting SOX9. Oncotarget 2017, 8, 574–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, L.K.; Lemoine, N.R.; Kirn, D. Oncolytic biotherapy: A novel therapeutic plafform. Lancet. Oncol. 2002, 3, 17–26. [Google Scholar] [CrossRef]
- Conry, R.M.; Westbrook, B.; McKee, S.; Norwood, T.G. Talimogene laherparepvec: First in class oncolytic virotherapy. Hum. Vaccines Immunother. 2018, 14, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Denard, B.; Lee, C.; Ye, J. Doxorubicin blocks proliferation of cancer cells through proteolytic activation of CREB3L1. eLife 2012, 1, e00090. [Google Scholar] [CrossRef] [PubMed]
- Bommareddy, P.K.; Shettigar, M.; Kaufman, H.L. Integrating oncolytic viruses in combination cancer immunotherapy. Nat. Rev. Immunol. 2018, 18, 498–513. [Google Scholar] [CrossRef]
- Xia, T.; Konno, H.; Barber, G.N. Recurrent Loss of STING Signaling in Melanoma Correlates with Susceptibility to Viral Oncolysis. Cancer Res. 2016, 76, 6747–6759. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gen | Forward | Reverse |
|---|---|---|
| ITGAV | GGCTGCATATTTCGGATTTTCTG | CCATTCAGCTTTGTCGTCTGG |
| MMP13 | GACTGGTAATGGCATCAAGGGA | CACCGGCAAAAGCCACTTTA |
| MMP1 | GCCAGATTTGCCAAGAGCAG | GAGTTGTCCCGATGATCTCCC |
| MMP9 | GTACCACGGCCAACTACGAC | GCCTTGGAAGATGAATGGAA |
| SOX9 | GTACCCGCACTTGCACAAC | TCTCGCTCTCGTTCAGAAGTC |
| TBP | GAACATCATGGATCAGAACAACA | ATAGGGATTCCGGGAGTCAT |
| 18s | CTGGATACCGCAGCTAGGAA | GAATTTCACCTCTAGCGGCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stöckl, S.; Lindner, G.; Li, S.; Schuster, P.; Haferkamp, S.; Wagner, F.; Prodinger, P.M.; Multhoff, G.; Boxberg, M.; Hillmann, A.; et al. SOX9 Knockout Induces Polyploidy and Changes Sensitivity to Tumor Treatment Strategies in a Chondrosarcoma Cell Line. Int. J. Mol. Sci. 2020, 21, 7627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207627
Stöckl S, Lindner G, Li S, Schuster P, Haferkamp S, Wagner F, Prodinger PM, Multhoff G, Boxberg M, Hillmann A, et al. SOX9 Knockout Induces Polyploidy and Changes Sensitivity to Tumor Treatment Strategies in a Chondrosarcoma Cell Line. International Journal of Molecular Sciences. 2020; 21(20):7627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207627
Chicago/Turabian StyleStöckl, Sabine, Georg Lindner, Shushan Li, Philipp Schuster, Sebastian Haferkamp, Ferdinand Wagner, Peter M. Prodinger, Gabriele Multhoff, Melanie Boxberg, Axel Hillmann, and et al. 2020. "SOX9 Knockout Induces Polyploidy and Changes Sensitivity to Tumor Treatment Strategies in a Chondrosarcoma Cell Line" International Journal of Molecular Sciences 21, no. 20: 7627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207627