Four and a Half LIM Domains 2 (FHL2) Contribute to the Epithelial Ovarian Cancer Carcinogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
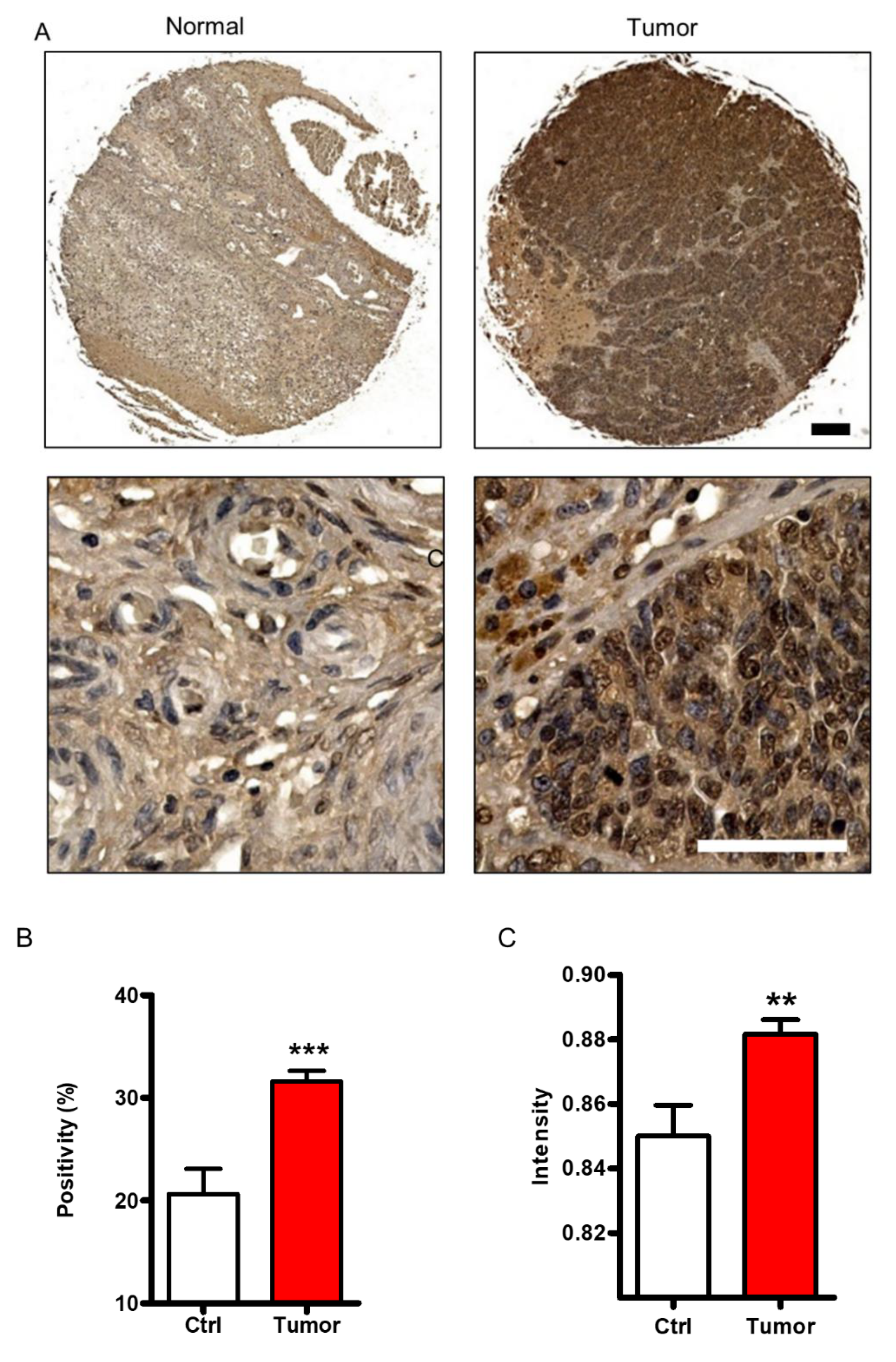
2.1. FHL2 Is Overexpressed in Human EOC Tissues
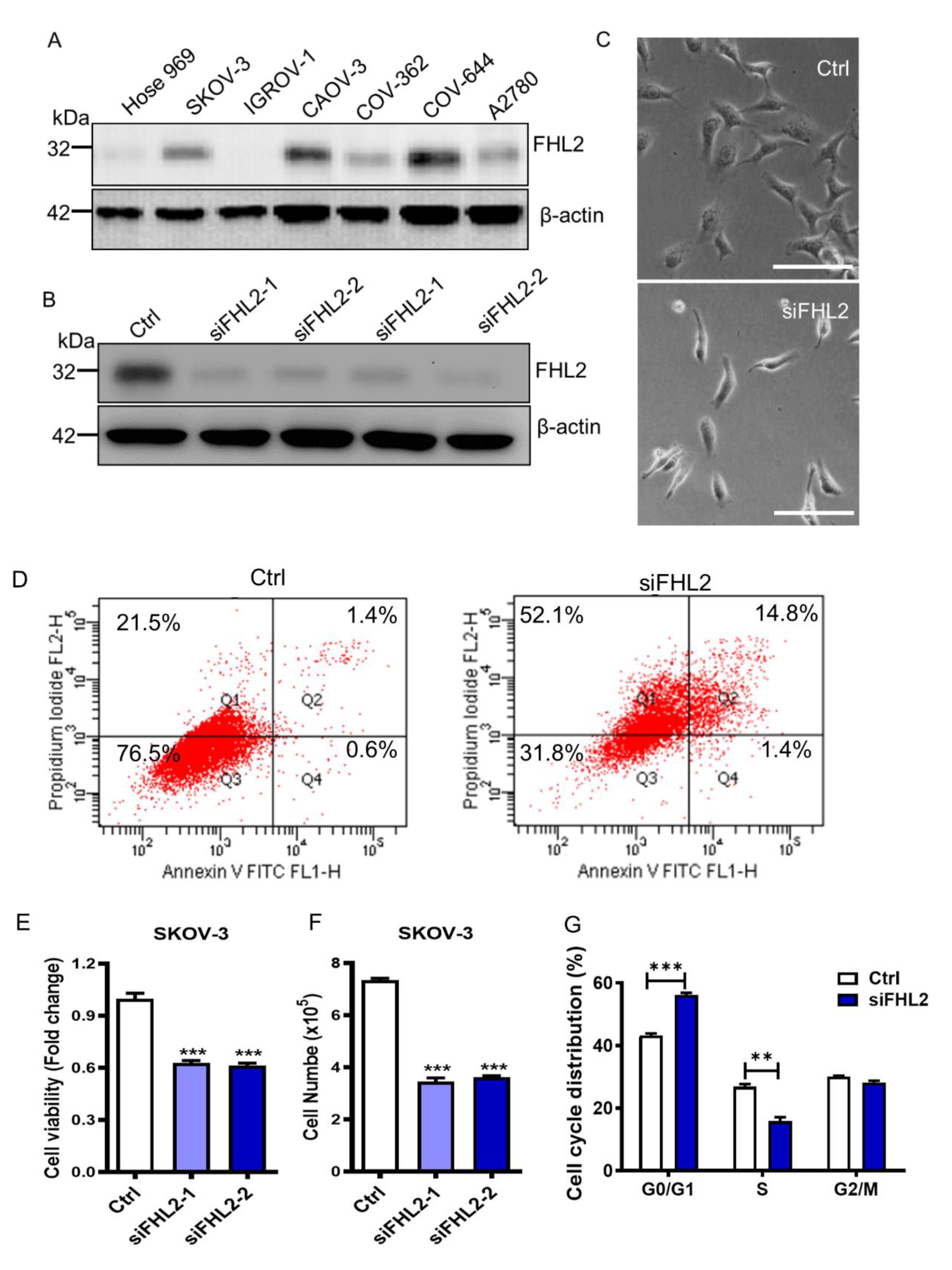
2.2. Knockdown of FHL2 in EOC Cells Inhibited Cell Growth
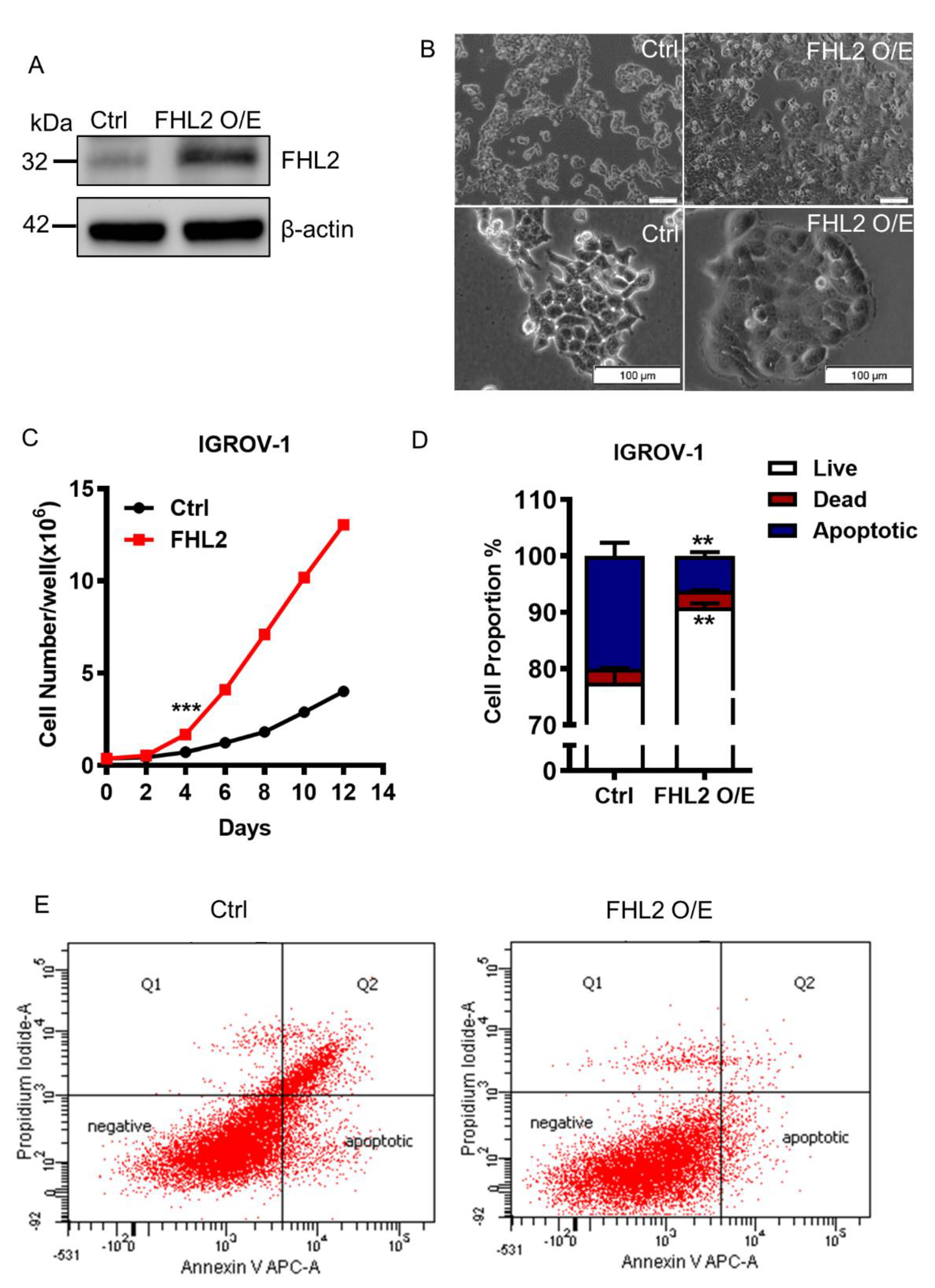
2.3. Ectopic Expression of FHL2 Improve EOC Cell Growth
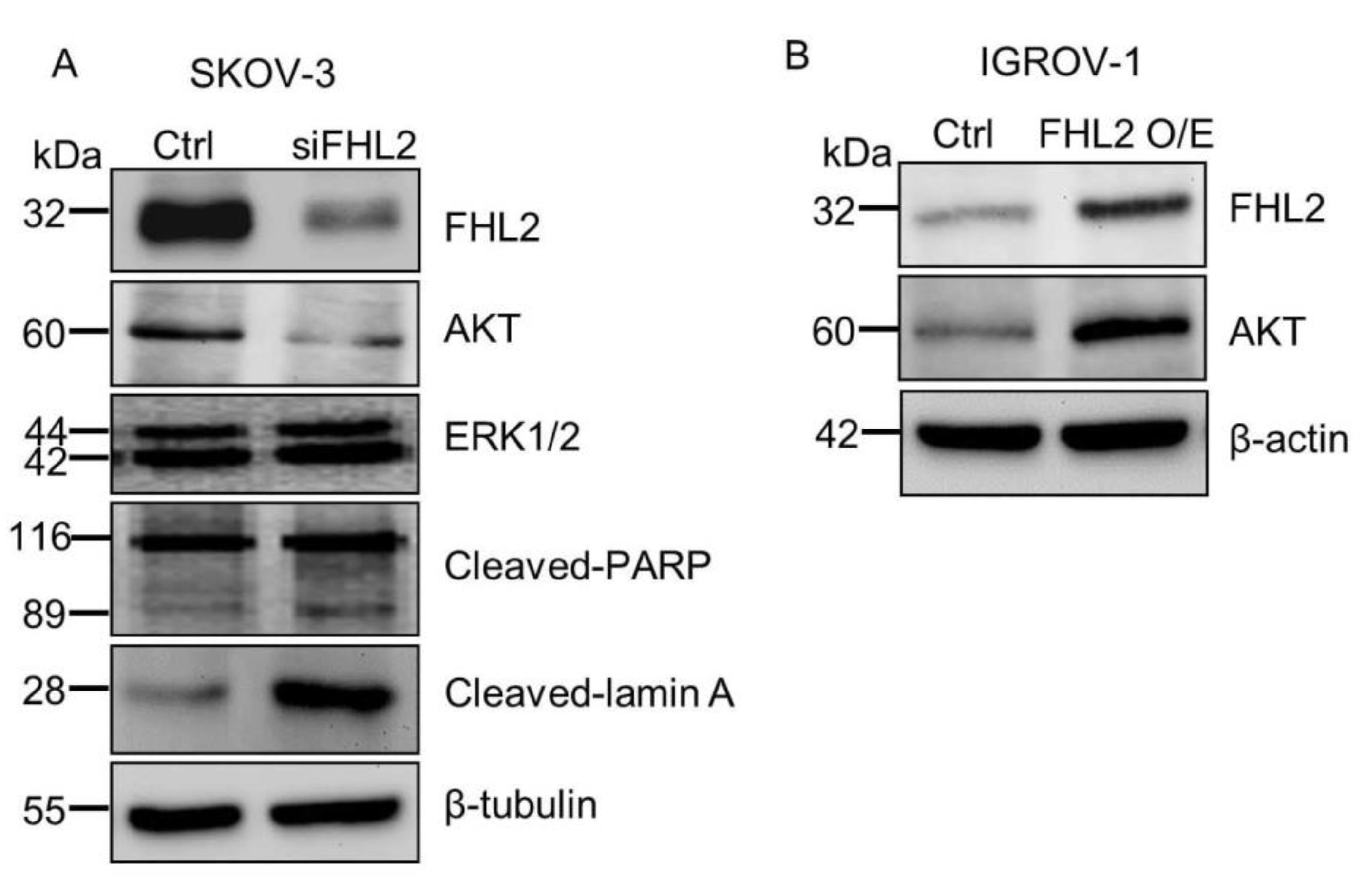
2.4. FHL2 Regulate AKT Expression in EOC Cells
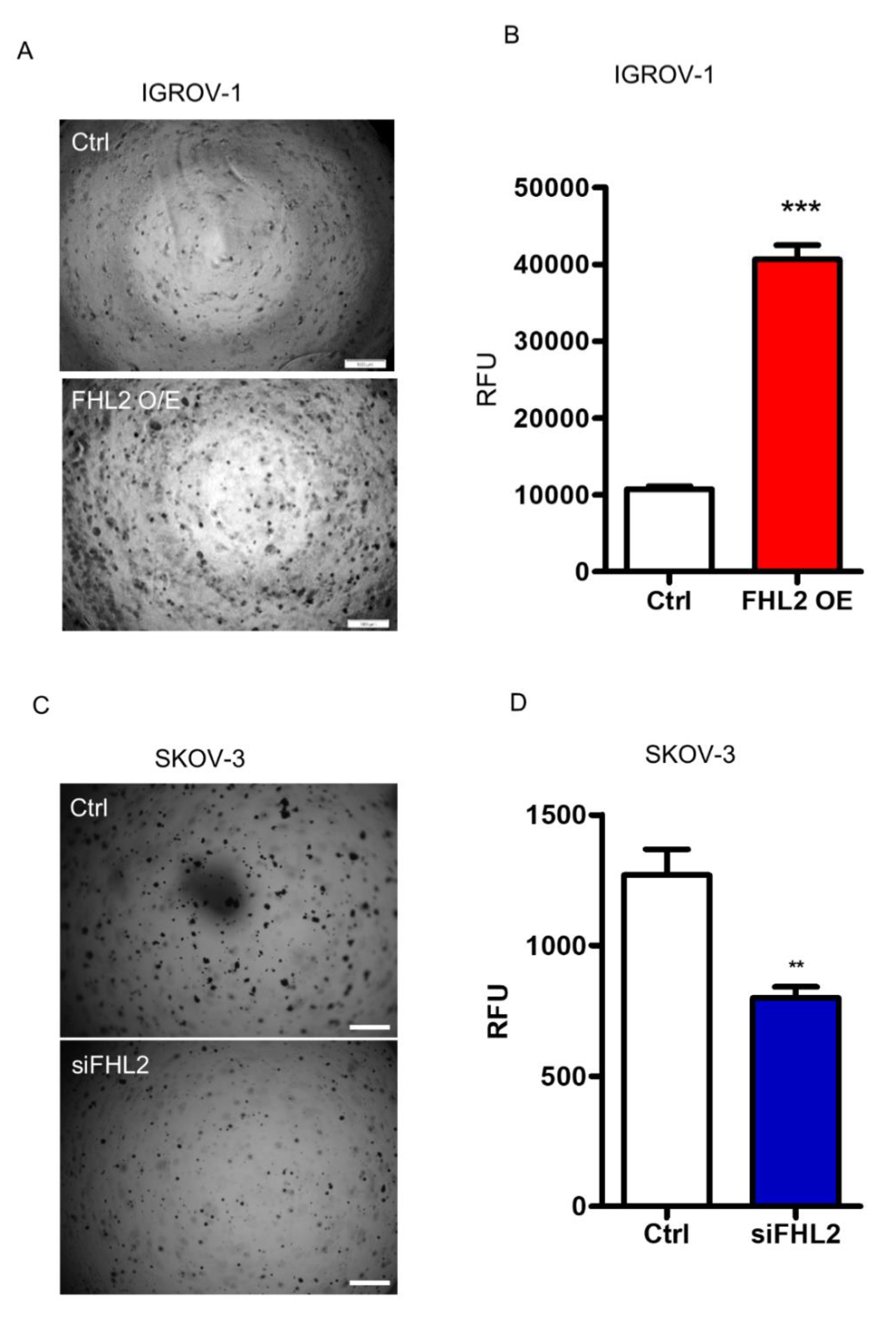
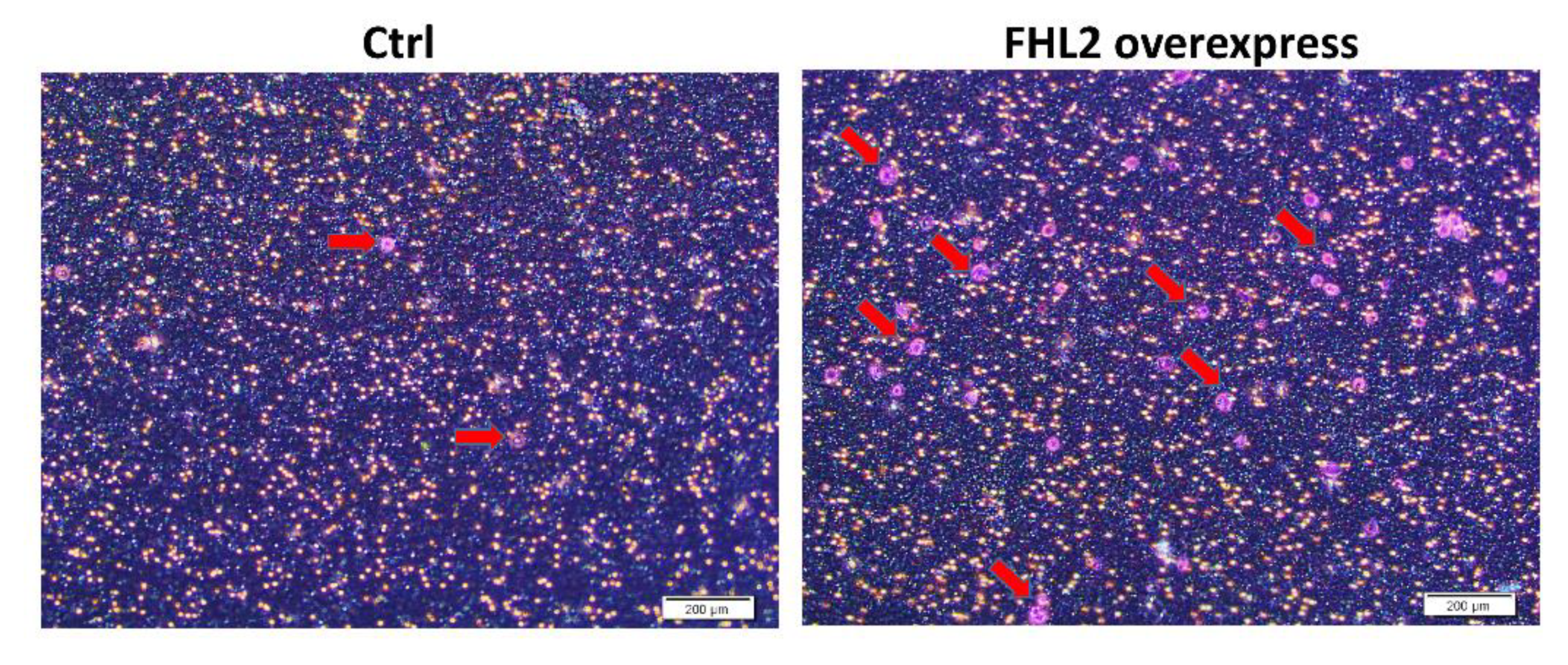
2.5. FHL2 Enhance Anchorage-Independent EOC Cell Growth
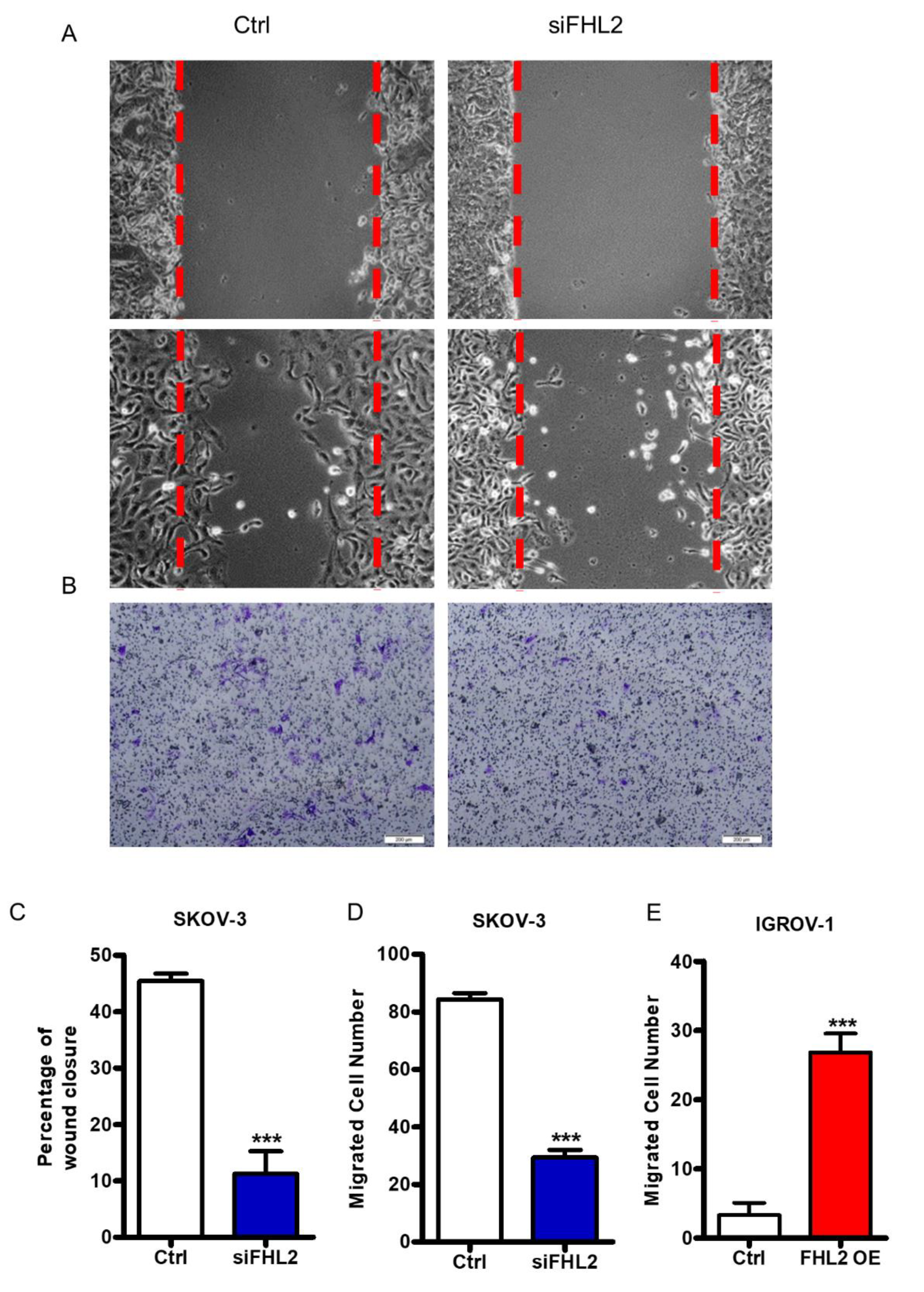
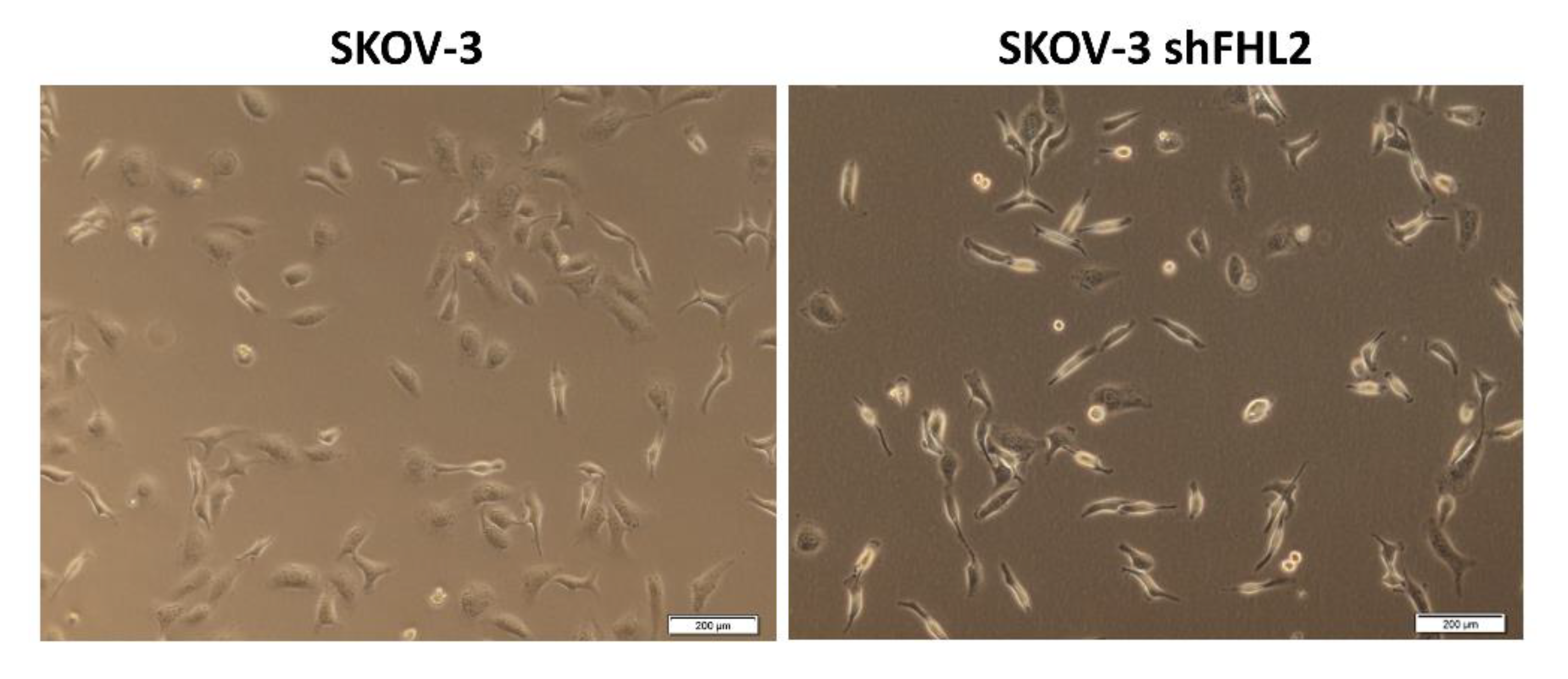
2.6. FHL2 Regulates EOC Cell Migration and Invasion In Vitro
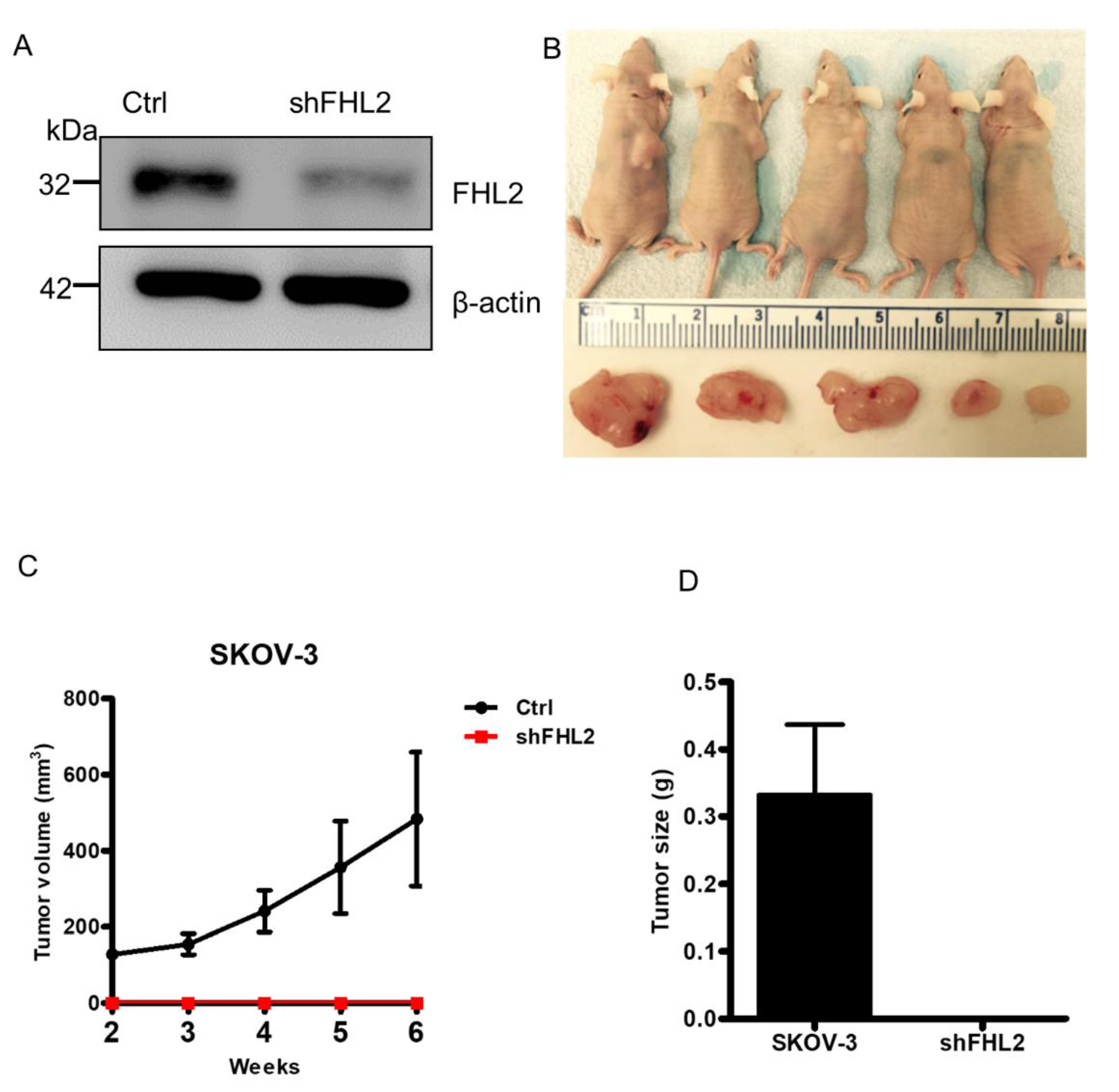
2.7. Knockdown of FHL2 Inhibited Tumorigenesis In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Human EOC Tissue Slides
4.2. Immunohistochemistry Staining
4.3. Western Blot Analysis
4.4. Cell Proliferation Analysis
4.5. Cell Viability Analysis
4.6. Cell Cycle and Apoptosis Analysis
4.7. Cell Migration Assays
4.8. Colony Formation Assay
4.9. Establishment of FHL2 Knockdown and Overexpression Stable Cell Lines
4.10. Tumorigenicity in Nude Mice
4.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Appendix A


References
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Hua, G.; Lv, X.; He, C.; Remmenga, S.W.; Rodabough, K.J.; Dong, J.; Yang, L.; Lele, S.M.; Yang, P.; Zhou, J.; et al. YAP induces high-grade serous carcinoma in fallopian tube secretory epithelial cells. Oncogene 2016, 35, 2247–2265. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Coffey, D.M.; Creighton, C.J.; Yu, Z.; Hawkins, S.M.; Matzuk, M.M. High-grade serous ovarian cancer arises from fallopian tube in a mouse model. Proc. Natl. Acad. Sci. USA 2012, 10, 3921–3926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testa, U.; Petrucci, E.; Pasquini, L.; Castelli, G.; Pelosi, E. Ovarian cancers: Genetic abnormalities, tumor heterogeneity and progression, clonal evolution and cancer stem cells. Medicines 2018, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wentzensen, N.; Poole, E.M.; Trabert, B.; White, E.; Arslan, A.A.; Patel, A.V.; Setiawan, V.W.; Visvanathan, K.; Weiderpass, E.; Adami, H.O.; et al. Ovarian cancer risk factors by histologic subtype: An analysis from the ovarian cancer cohort consortium. J. Clin. Oncol. 2016, 34, 2888. [Google Scholar] [CrossRef]
- Jones, M.R.; Kamara, D.; Karlan, B.Y.; Pharoah, P.D.; Gayther, S.A. Genetic epidemiology of ovarian cancer and prospects for polygenic risk prediction. Gynecol. Oncol. 2017, 147, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Manderson, E.N.; Presneau, N.; Provencher, D.; Mes-Masson, A.M.; Tonin, P.N. Comparative analysis of loss of heterozygosity of specific chromosome 3, 13, 17, and X loci and TP53 mutations in human epithelial ovarian cancer. Mol. Carcinog. Publ. Coop. Univ. Tex. MD Anderson Cancer Center 2002, 34, 78–90. [Google Scholar]
- Alsop, K.; Fereday, S.; Meldrum, C.; DeFazio, A.; Emmanuel, C.; George, J.; Dobrovic, A.; Birrer, M.J.; Webb, P.M.; Stewart, C.; et al. BRCA mutation frequency and patterns of treatment response in BRCA mutation–positive women with ovarian cancer: A report from the Australian Ovarian Cancer Study Group. J. Clin. Oncol. 2012, 30, 2654. [Google Scholar] [CrossRef] [Green Version]
- Samartzis, E.P.; Noske, A.; Dedes, K.J.; Fink, D.; Imesch, P. ARID1A mutations and PI3K/AKT pathway alterations in endometriosis and endometriosis-associated ovarian carcinomas. Int. J. Mol. Sci. 2013, 14, 18824–18849. [Google Scholar] [CrossRef] [Green Version]
- Martins, F.C.; de Santiago, I.; Trinh, A.; Xian, J.; Guo, A.; Sayal, K.; Jimenez-Linan, M.; Deen, S.; Driver, K.; Mack, M.; et al. Combined image and genomic analysis of high-grade serous ovarian cancer reveals PTEN loss as a common driver event and prognostic classifier. Genome Biol. 2014, 15, 526. [Google Scholar] [CrossRef] [Green Version]
- Giles, R.H.; van Es, J.H.; Clevers, H. Caught up in a Wnt storm: Wnt signaling in cancer. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2003, 1653, 1–24. [Google Scholar] [CrossRef]
- He, C.; Lv, X.; Hua, G.; Lele, S.M.; Remmenga, S.; Dong, J.; Davis, J.S.; Wang, C. YAP forms autocrine loops with the ERBB pathway to regulate ovarian cancer initiation and progression. Oncogene 2015, 34, 6040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Dai, X.; Gao, Y.; Shen, F.; Ding, J.; Chen, Q. The positivity of estrogen receptor and progesterone receptor may not be associated with metastasis and recurrence in epithelial ovarian cancer. Sci. Rep. 2017, 7, 16922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Dai, X.; Gao, Y.; Shen, F.; Ding, J.; Chen, Q. FHL2: A scaffold protein of carcinogenesis, tumour-stroma interactions and treatment response. Histol. Histopathol. 2016, 31, 469–478. [Google Scholar]
- Ng, C.F.; Ng, P.K.; Lui, V.W.; Li, J.; Chan, J.Y.; Fung, K.P.; Ng, Y.K.; Bo-San Lai, P.; Tsui, S.K. FHL2 exhibits anti-proliferative and anti-apoptotic activities in liver cancer cells. Cancer Lett. 2011, 304, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.H.; Yeh, L.K.; Lin, H.C.; Jung, S.M.; Ma, D.H.; Wang, I.J.; Wu, H.H.; Shiu, T.F.; Chen, J. Deletion of the FHL2 gene attenuating neovascularization after corneal injury. Invest. Ophthalmol. Vis. Sci. 2008, 49, 5314–5318. [Google Scholar]
- Xue, Y.L.; Meng, X.Q.; Ma, L.J.; Yuan, Z. Plumbagin exhibits an anti-proliferative effect in human osteosarcoma cells by downregulating FHL2 and interfering with Wnt/β-catenin signalling. Oncol. Lett. 2016, 12, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- Hua, G.; He, C.; Lv, X.; Fan, L.; Wang, C.; Remmenga, S.W.; Rodabaugh, K.J.; Yang, L.; Lele, S.M.; Yang, P.; et al. The four and a half LIM domains 2 (FHL2) regulates ovarian granulosa cell tumor progression via controlling AKT1 transcription. Cell Death Dis. 2016, 7, e2297. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, B.; Mildenberger, S.; Weisser, C.W.; Metzger, E.; Gitsch, G.; Schüle, R.; Müller, J.M. Focal adhesion kinase interacts with the transcriptional coactivator FHL2 and both are overexpressed in epithelial ovarian cancer. Anticancer Res. 2004, 24, 921–928. [Google Scholar] [PubMed]
- Hernandez, L.; Kim, M.K.; Lyle, L.T.; Bunch, K.P.; House, C.D.; Ning, F.; Noonan, A.M.; Annunziata, C.M. Characterization of ovarian cancer cell lines as in vivo models for preclinical studies. Gynecol. Oncol. 2016, 142, 332–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, B.D.; Toker, A. AKT/PKB signaling: Navigating the network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Paul, C.D.; Mistriotis, P.; Konstantopoulos, K. Cancer cell motility: Lessons from migration in confined spaces. Nat. Rev. Cancer 2017, 17, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zienert, E.; Eke, I.; Aust, D.; Cordes, N. LIM-only protein FHL2 critically determines survival and radioresistance of pancreatic cancer cells. Cancer Lett. 2015, 364, 17–24. [Google Scholar] [CrossRef]
- Martin, B.T.; Kleiber, K.; Wixler, V.; Raab, M.; Zimmer, B.; Kaufmann, M.; Strebhardt, K. FHL2 regulates cell cycle-dependent and doxorubicin-induced p21Cip1/Waf1 expression in breast cancer cells. Cell Cycle 2007, 6, 1779–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Wang, J.; Tang, W.; Zhan, X.; Li, Y.; Peng, Y.; Huang, X.; Bai, Y.; Zhao, J.; Li, A.; et al. FOXK1 interaction with FHL2 promotes proliferation, invasion and metastasis in colorectal cancer. Oncogenesis 2016, 5, e271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labalette, C.; Nouët, Y.; Sobczak-Thepot, J.; Armengol, C.; Levillayer, F.; Gendron, M.C.; Renard, C.A.; Regnault, B.; Chen, J.; Buendia, M.A.; et al. The LIM-only protein FHL2 regulates cyclin D1 expression and cell proliferation. J. Biol. Chem. 2008, 283, 15201–15208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves-Perez, A.; Mack, B.; Maetzel, D.; Kremling, H.; Eggert, C.; Harreus, U.; Gires, O. EpCAM regulates cell cycle progression via control of cyclin D1 expression. Oncogene 2013, 32, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, H.; Wyckoff, J.; Condeelis, J. Cell migration in tumors. Curr. Opin. Cell Biol. 2005, 17, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Boateng, L.R.; Bennin, D.; De Oliveira, S.; Huttenlocher, A. Mammalian Actin-binding Protein-1/Hip-55 Interacts with FHL2 and Negatively Regulates Cell Invasion. J. Biol. Chem. 2016, 291, 13987–13998. [Google Scholar] [CrossRef] [Green Version]
- Canault, M.; Tellier, E.; Bonardo, B.; Mas, E.; Aumailley, M.; Juhan-Vague, I.; Nalbone, G.; Peiretti, F. FHL2 interacts with both ADAM-17 and the cytoskeleton and regulates ADAM-17 localization and activity. J. Cell. Physiol. 2006, 208, 363–372. [Google Scholar] [CrossRef]
- Jason, S.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar]
- Follo, M.Y.; Manzoli, L.; Poli, A.; McCubrey, J.A.; Cocco, L. PLC and PI3K/Akt/mTOR signalling in disease and cancer. Adv. Biol. Regul. 2015, 57, 10–16. [Google Scholar] [CrossRef]
- Abeyrathna, P.; Su, Y. The critical role of Akt in cardiovascular function. Vascul. Pharmacol. 2015, 74, 38–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Yu, S.; Xu, H.; Zheng, Y.; Lin, J.; Wu, M.; Wang, J.; Wang, A.; Lan, Q.; Furnari, F.; et al. FHL2 interacts with EGFR to promote glioblastoma growth. Oncogene 2018, 37, 1386. [Google Scholar] [CrossRef]
- Chen, L.; Lu, Y.; Wen, J.; Wang, X.; Wu, L.; Wu, D.; Sun, X.; Fu, B.; Yin, Z.; Jiang, H.; et al. Comparative proteomics analysis of mouse Habu nephritis models with and without unilateral nephrectomy. Cell. Physiol. Biochem. 2016, 39, 1761–1776. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lv, X.; Jiang, C.; Cordes, C.M.; Fu, L.; Lele, S.M.; Davis, J.S. Transforming growth factor alpha (TGFα) regulates granulosa cell tumor (GCT) cell proliferation and migration through activation of multiple pathways. PLoS ONE 2012, 7, e48299. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Lv, X.; Hua, G.; He, C.; Dong, J.; Lele, S.M.; Li, D.W.; Zhai, Q.; Davis, J.S.; Wang, C. YAP regulates cell proliferation, migration, and steroidogenesis in adult granulosa cell tumors. Endocr. Relat. Cancer 2014, 21, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Lv, X.; He, C.; Hua, G.; Tsai, M.Y.; Davis, J.S. The G-protein-coupled estrogen receptor agonist G-1 suppresses proliferation of ovarian cancer cells by blocking tubulin polymerization. Cell Death Dis. 2013, 4, e869. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Lv, X.; He, C.; Davis, J.S.; Wang, C.; Hua, G. Four and a Half LIM Domains 2 (FHL2) Contribute to the Epithelial Ovarian Cancer Carcinogenesis. Int. J. Mol. Sci. 2020, 21, 7751. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207751
Wang C, Lv X, He C, Davis JS, Wang C, Hua G. Four and a Half LIM Domains 2 (FHL2) Contribute to the Epithelial Ovarian Cancer Carcinogenesis. International Journal of Molecular Sciences. 2020; 21(20):7751. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207751
Chicago/Turabian StyleWang, Chen, Xiangmin Lv, Chunbo He, John S. Davis, Cheng Wang, and Guohua Hua. 2020. "Four and a Half LIM Domains 2 (FHL2) Contribute to the Epithelial Ovarian Cancer Carcinogenesis" International Journal of Molecular Sciences 21, no. 20: 7751. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207751