Capacity of Retinal Ganglion Cells Derived from Human Induced Pluripotent Stem Cells to Suppress T-Cells

 ,
,
Abstract
:
1. Introduction
2. Results
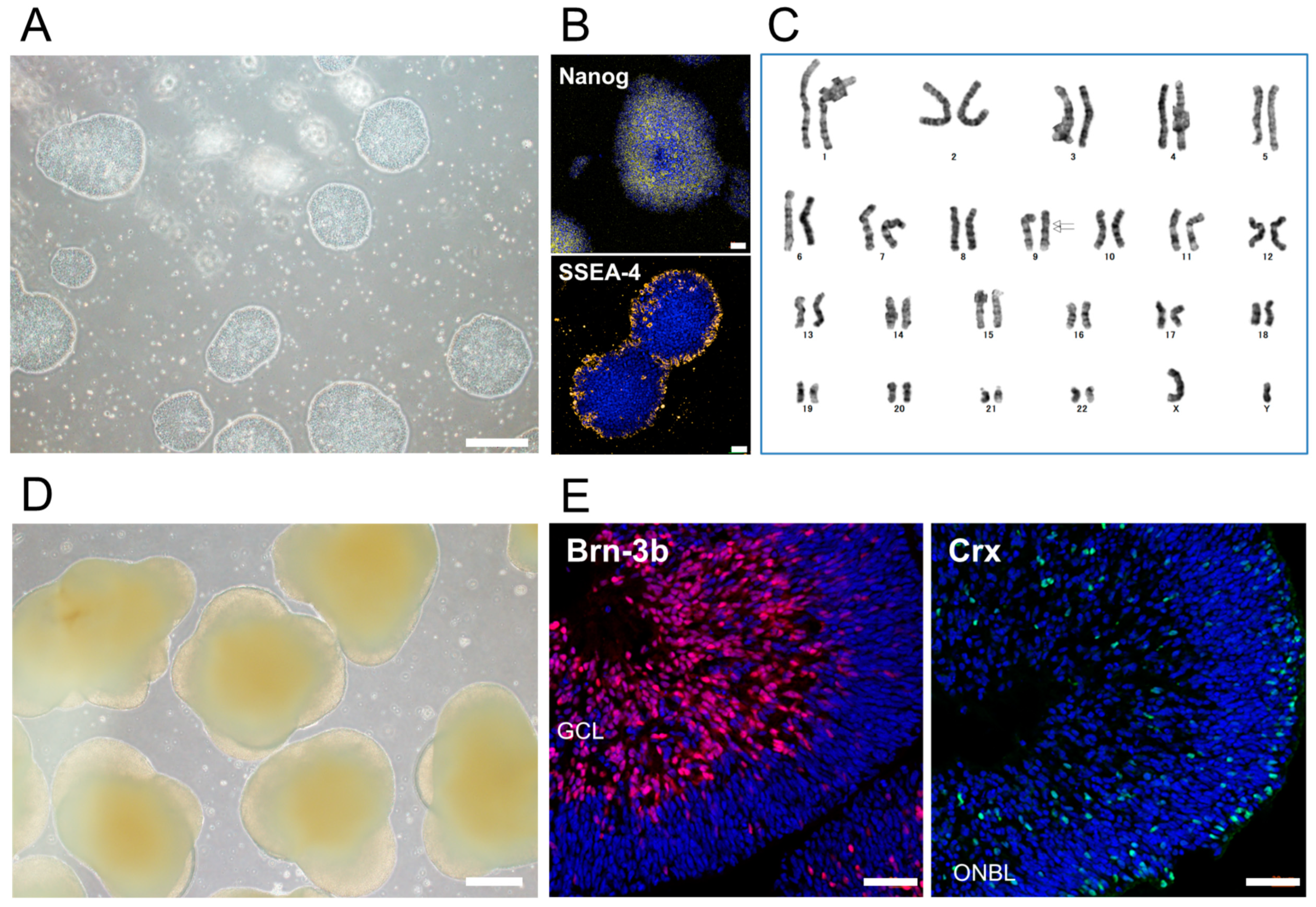
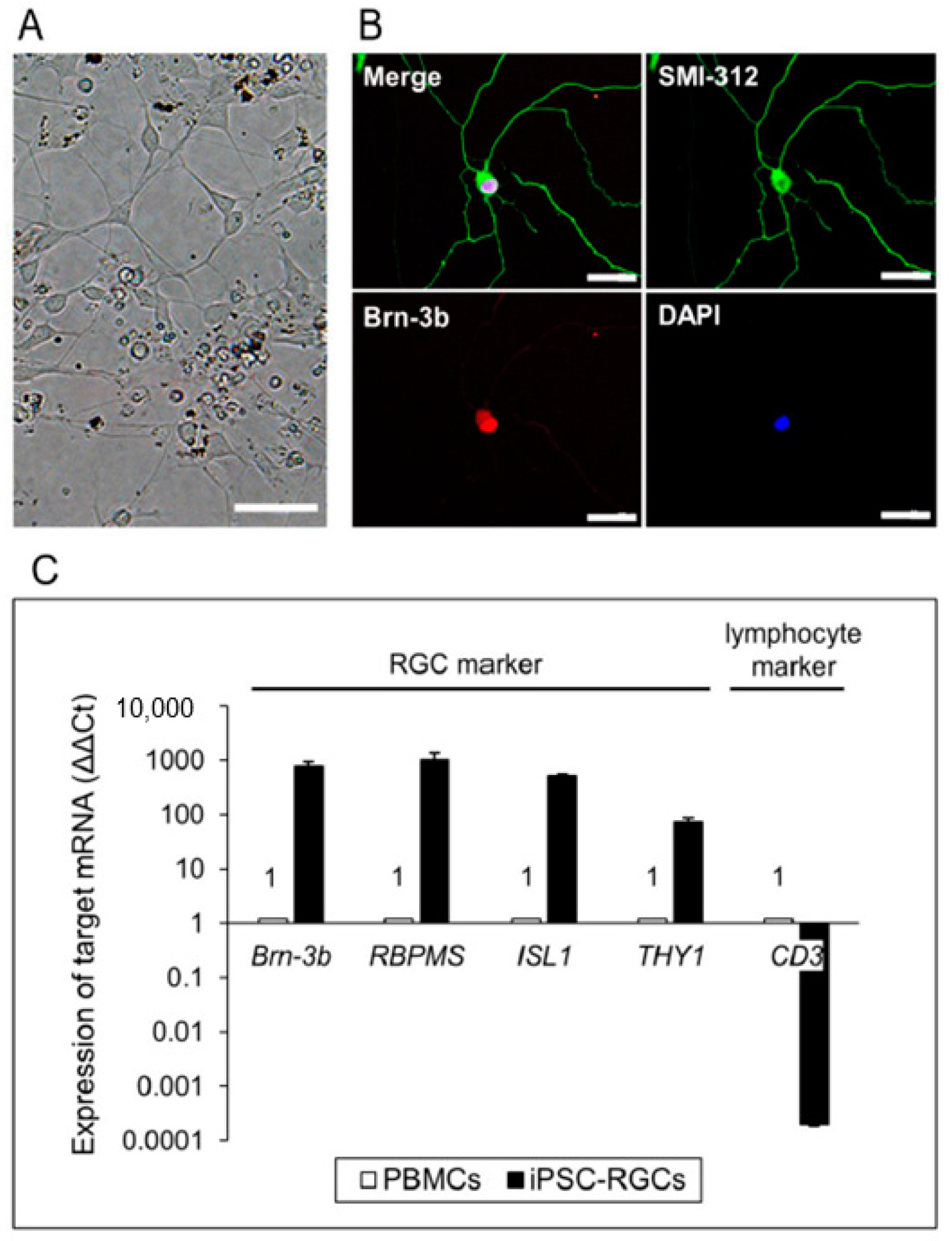
2.1. Preparation of iPSC-RGCs
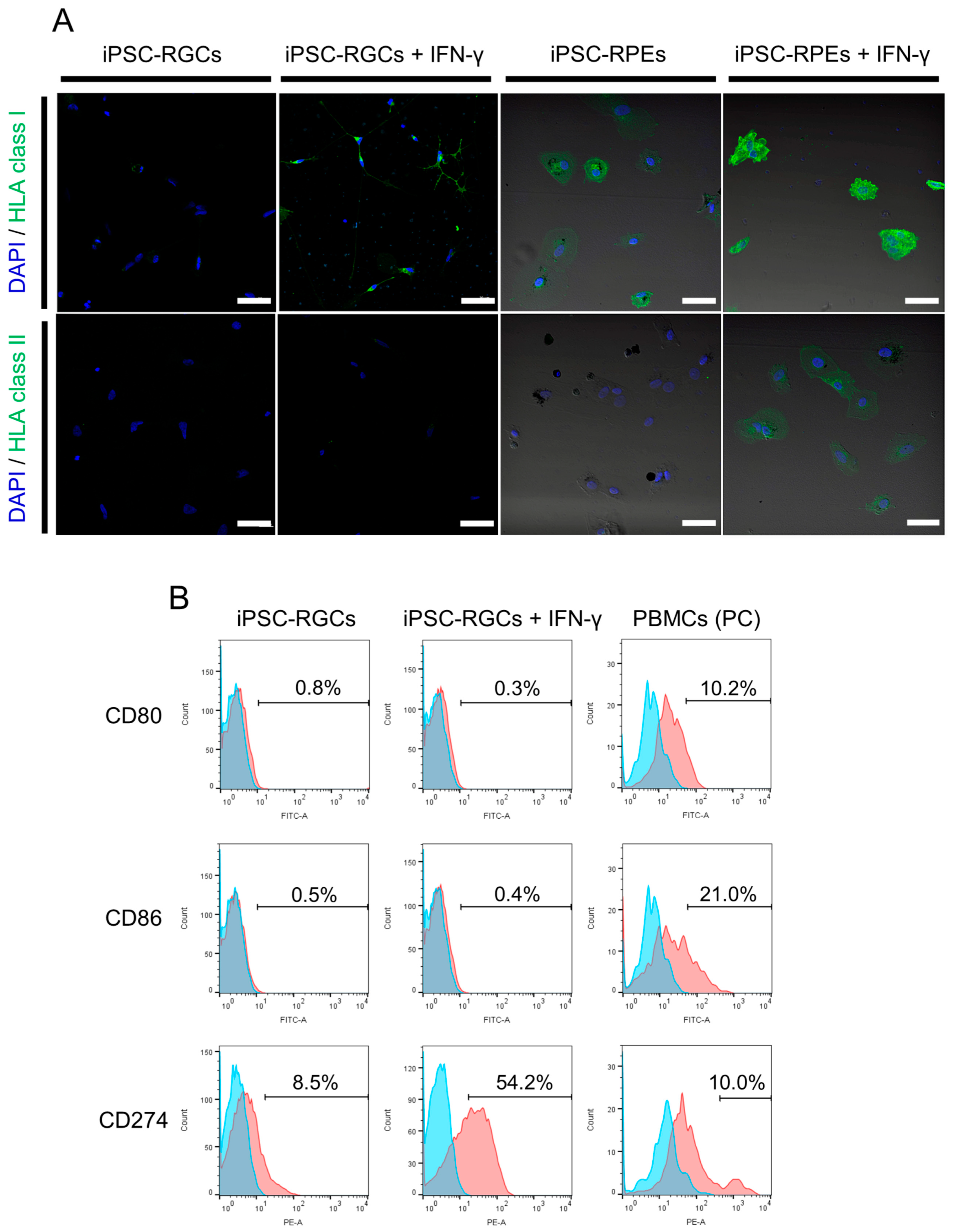
2.2. Immunogenicity of iPSC-RGCs Assessed with HLA Class I, Class II, and Co-Stimulatory Molecules
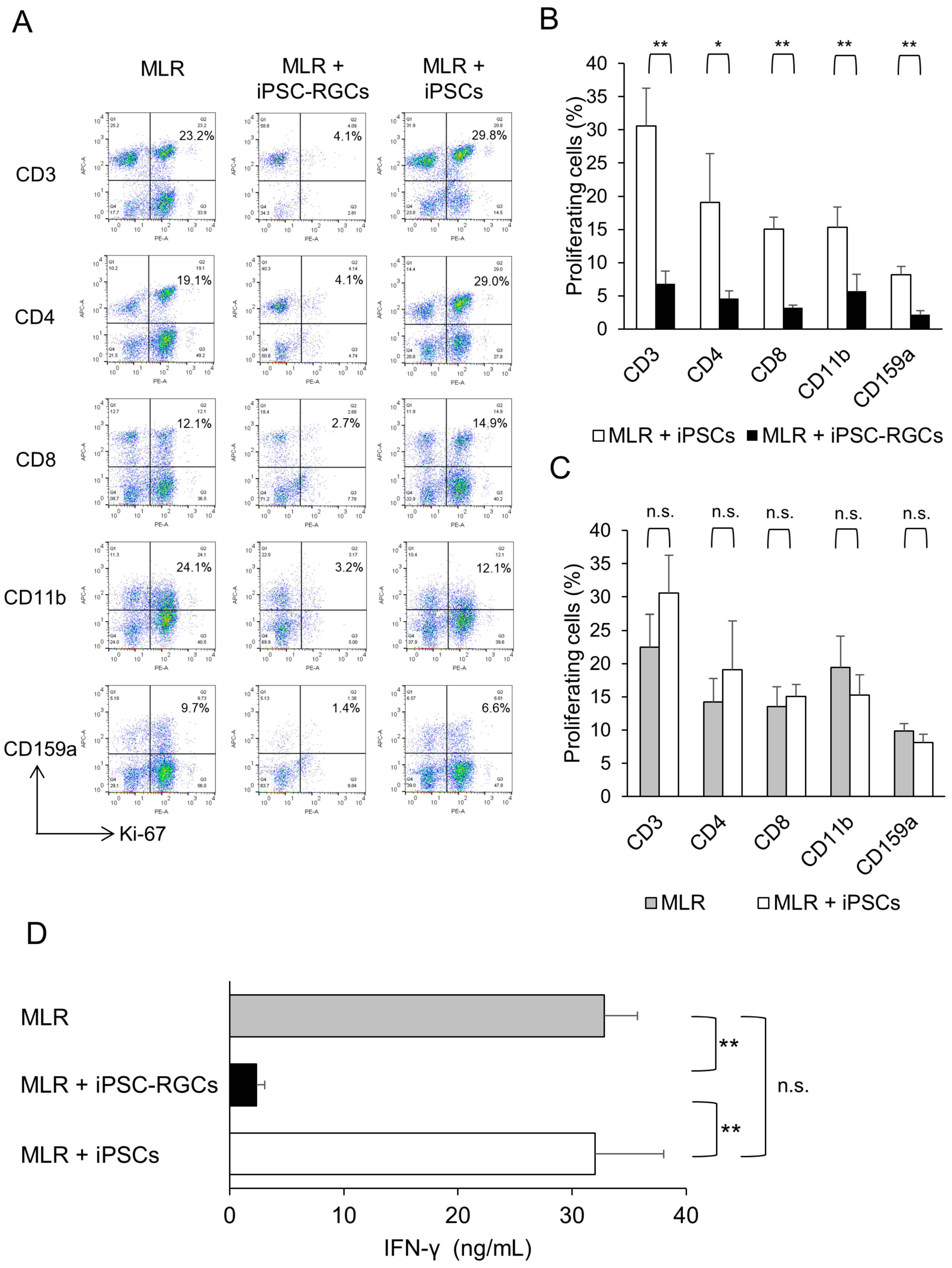
2.3. Human iPSC-RGCs are able to Suppress Immune Cell Activation In Vitro
2.4. Elucidating of the Immunosuppressive Mechanism of iPSC-RGCs
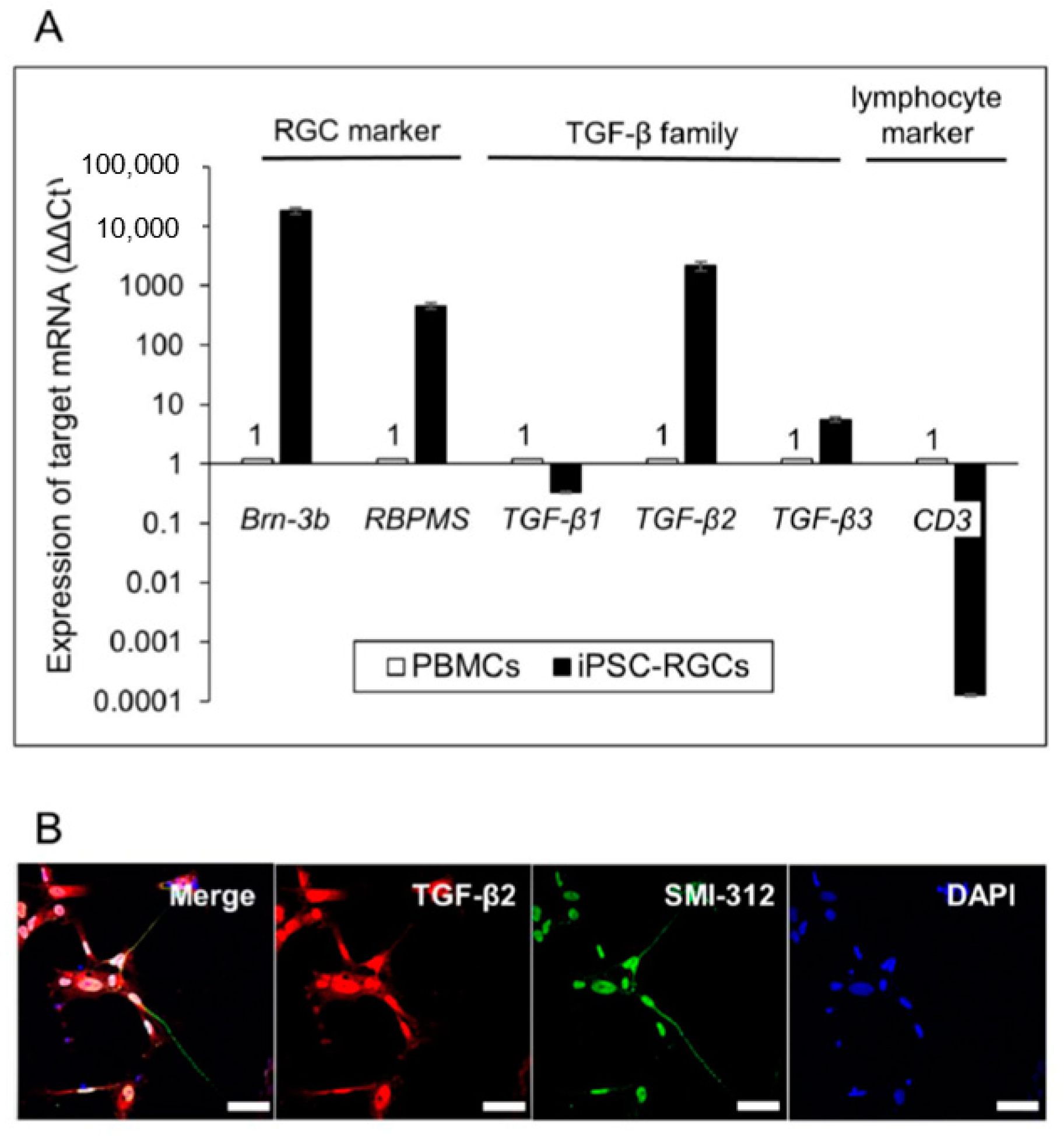
2.5. Detection of TGF-β in iPSC-RGCs
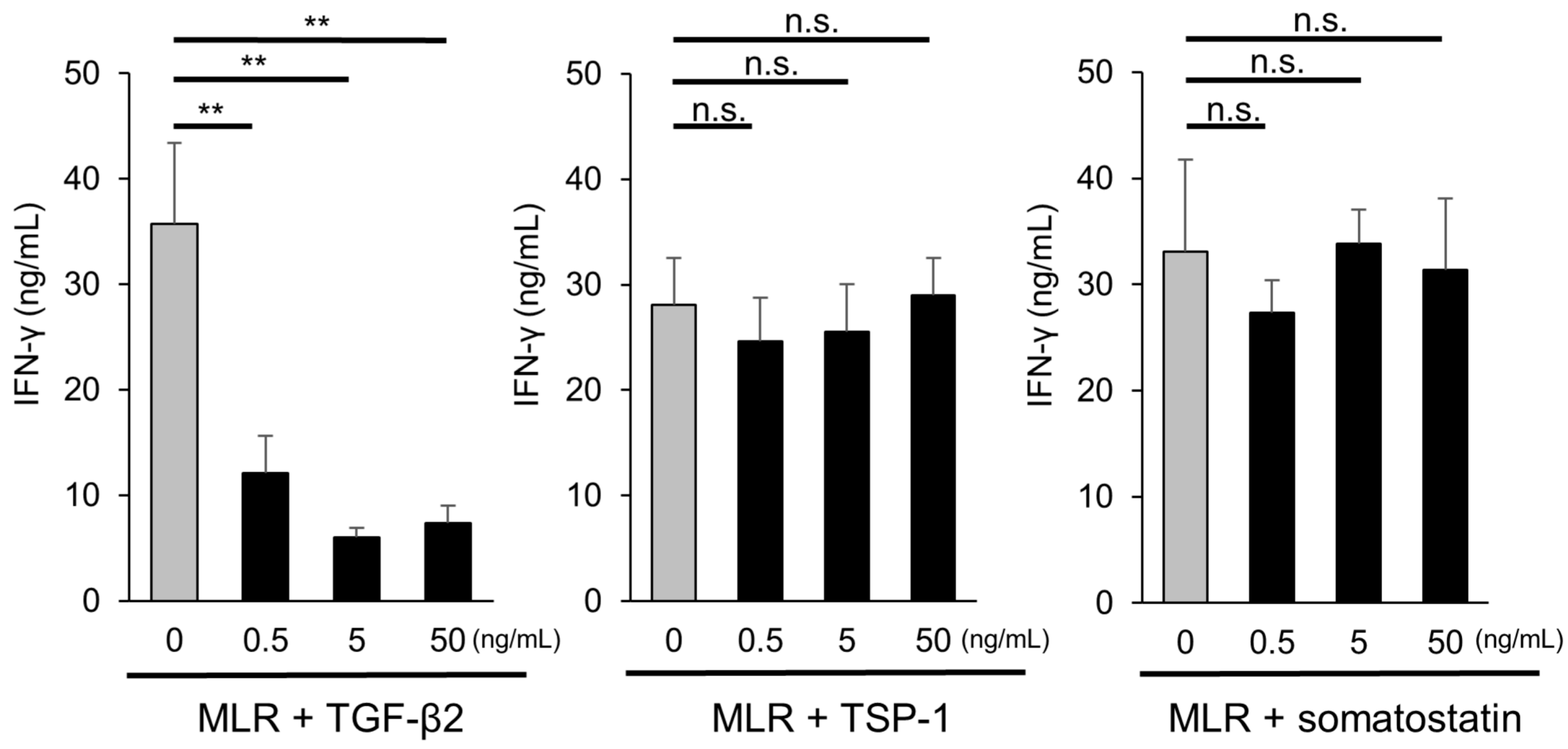
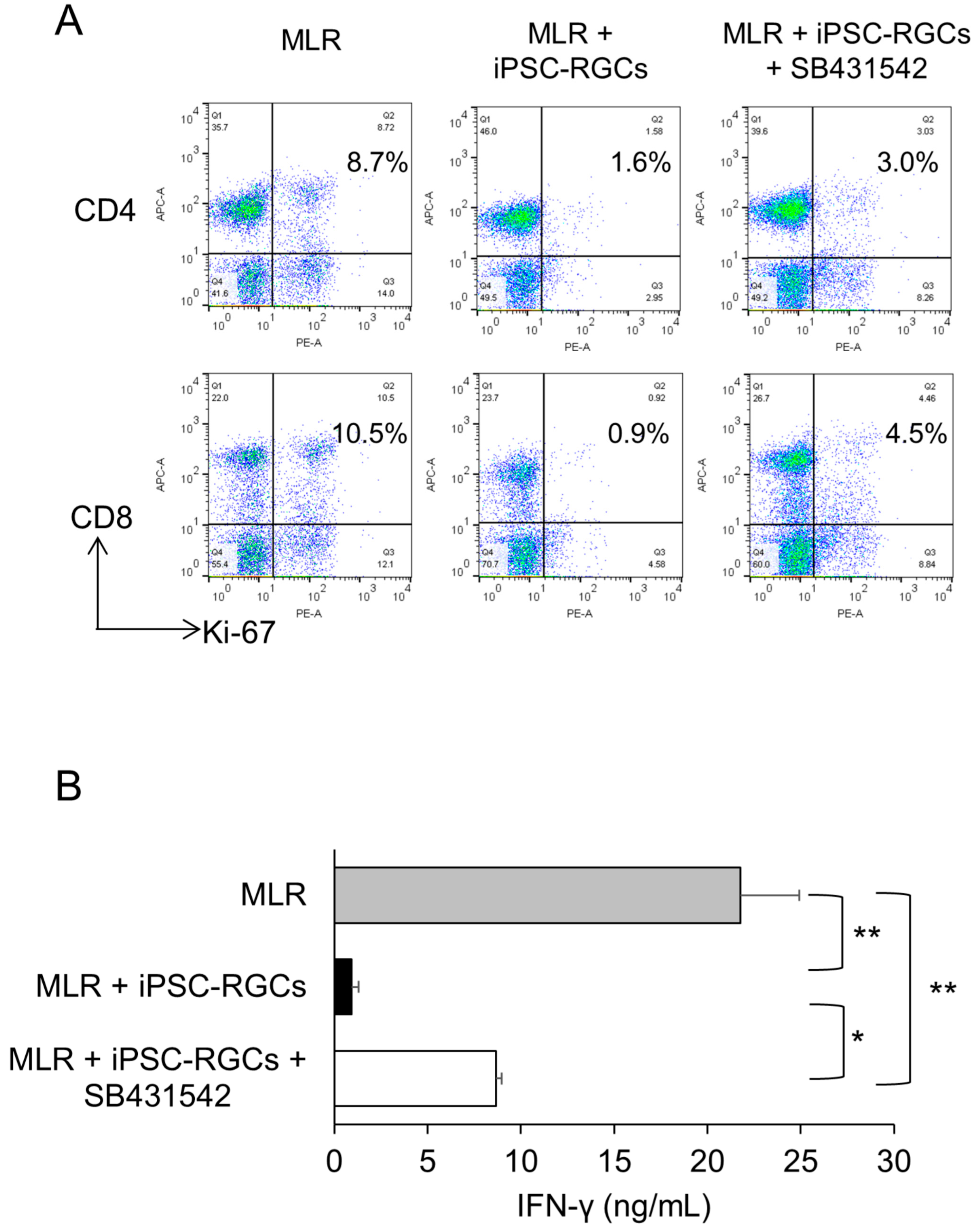
2.6. Role of TGF-β in T-Cell Suppression by iPSC-RGCs
3. Discussion
4. Materials and Methods
4.1. Establishment of Human iPSCs
4.2. Preparation, Isolation, and Culture of iPSC-RGCs
4.3. Flow Cytometry
4.4. Quantitative RT-PCR
4.5. IHC and ICC
4.6. MLR Assay with iPSC-RGCs
4.7. T-cell Proliferation Assay with iPSC-RGCs
4.8. Expression Analysis with Microarray
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FACS | fluorescence-activated cell sorting |
| HLA | human leukocyte antigen |
| ICC | Immunocytochemistry |
| IHC | Immunohistochemistry |
| iPSCs | induced pluripotent stem cells |
| MLR | mixed lymphocyte reaction |
| NK | natural killer |
| PBMCs | peripheral blood mononuclear cells |
| RGCs | retinal ganglion cells |
| RPEs | retinal pigment epithelial cells |
| RT-PCR | reverse transcription-polymerase chain reaction |
References
- Jin, Z.-B.; Gao, M.-L.; Deng, W.-L.; Wu, K.-C.; Sugita, S.; Mandai, M.; Takahashi, M. Stemming retinal regeneration with pluripotent stem cells. Prog. Retin. Eye Res. 2019, 69, 38–56. [Google Scholar] [CrossRef]
- Kæstel, C.G.; Jørgensen, A.; Nielsen, M.; Eriksen, K.W.; Ødum, N.; Holst Nissen, M.; Röpke, C. Human Retinal Pigment Epithelial Cells Inhibit Proliferation and IL2R Expression of Activated T Cells. Exp. Eye Res. 2002, 74, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Wallace, C.A.; Moir, G.; Malone, D.F.G.; Duncan, L.; Devarajan, G.; Crane, I.J. Regulation of T-Lymphocyte CCL3 and CCL4 Production by Retinal Pigment Epithelial Cells. Investig. Opthalmology Vis. Sci. 2013, 54, 722–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugita, S.; Kamao, H.; Iwasaki, Y.; Okamoto, S.; Hashiguchi, T.; Iseki, K.; Hayashi, N.; Mandai, M.; Takahashi, M. Inhibition of T-Cell Activation by Retinal Pigment Epithelial Cells Derived From Induced Pluripotent Stem Cells. Invest. Ophthalmol. Vis. Sci. 2015, 56, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; He, J.; Tang, L.; Kong, Y.; Shi, Y.; Chen, S.; Huang, Z. Transforming growth factor-β2 increases the capacity of retinal pigment epithelial cells to induce the generation of regulatory T cells. Mol. Med. Report. 2016, 13, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Idelson, M.; Alper, R.; Obolensky, A.; Yachimovich-Cohen, N.; Rachmilewitz, J.; Ejzenberg, A.; Beider, E.; Banin, E.; Reubinoff, B. Immunological Properties of Human Embryonic Stem Cell-Derived Retinal Pigment Epithelial Cells. Stem Cell Rep. 2018, 11, 681–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xu, H.W.; Wang, L.; Li, S.Y.; Zhao, C.J.; Hao, J.; Li, Q.Y.; Zhao, T.T.; Wu, W.; Wang, Y.; et al. Human embryonic stem cell-derived retinal pigment epithelium transplants as a potential treatment for wet age-related macular degeneration. Cell Discov. 2018, 4, 50. [Google Scholar] [CrossRef]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; et al. Autologous Induced Stem-Cell–Derived Retinal Cells for Macular Degeneration. N. Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef]
- Mehat, M.S.; Sundaram, V.; Ripamonti, C.; Robson, A.G.; Smith, A.J.; Borooah, S.; Robinson, M.; Rosenthal, A.N.; Innes, W.; Weleber, R.G.; et al. Transplantation of Human Embryonic Stem Cell-Derived Retinal Pigment Epithelial Cells in Macular Degeneration. Ophthalmology 2018, 125, 1765–1775. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Mandai, M.; Hirami, Y.; Takagi, S.; Maeda, T.; Fujihara, M.; Matsuzaki, M.; Yamamoto, M.; Iseki, K.; Hayashi, N.; et al. HLA-Matched Allogeneic iPS Cells-Derived RPE Transplantation for Macular Degeneration. J. Clin. Med. 2020, 9, 2217. [Google Scholar] [CrossRef]
- Toosy, A.T.; Mason, D.F.; Miller, D.H. Optic neuritis. Lancet Neurol. 2014, 13, 83–99. [Google Scholar] [CrossRef]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The Pathophysiology and Treatment of Glaucoma. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Miltner, A.M.; La Torre, A. Retinal Ganglion Cell Replacement: Current Status and Challenges Ahead. Dev. Dyn. 2018, 248, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Quigley, H.A. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Choi, S.H.; Kim, Y.B.; Jun, I.; Sung, J.J.; Lee, D.R.; Kim, Y.I.; Cho, M.S.; Byeon, S.H.; Kim, D.S.; et al. Defined Conditions for Differentiation of Functional Retinal Ganglion Cells From Human Pluripotent Stem Cells. Invest. Ophthalmol. Vis. Sci. 2018, 59, 3531–3542. [Google Scholar] [CrossRef] [Green Version]
- Venugopalan, P.; Wang, Y.; Nguyen, T.; Huang, A.; Muller, K.J.; Goldberg, J.L. Transplanted neurons integrate into adult retinas and respond to light. Nat. Commun. 2016, 7, 10472. [Google Scholar] [CrossRef]
- Suen, H.C.; Qian, Y.; Liao, J.; Luk, C.S.; Lee, W.T.; Ng, J.K.W.; Chan, T.T.H.; Hou, H.W.; Li, I.; Li, K.; et al. Transplantation of Retinal Ganglion Cells Derived from Male Germline Stem Cell as a Potential Treatment to Glaucoma. Stem Cells Dev. 2019, 28, 1365–1375. [Google Scholar] [CrossRef]
- Hertz, J.; Qu, B.; Hu, Y.; Patel, R.D.; Valenzuela, D.A.; Goldberg, J.L. Survival and Integration of Developing and Progenitor-Derived Retinal Ganglion Cells following Transplantation. Cell Transplant. 2014, 23, 855–872. [Google Scholar] [CrossRef]
- Cen, L.-P.; Ng, T.K.; Liang, J.-J.; Zhuang, X.; Yao, X.; Yam, G.H.-F.; Chen, H.; Cheung, H.S.; Zhang, M.; Pang, C.P. Human Periodontal Ligament-Derived Stem Cells Promote Retinal Ganglion Cell Survival and Axon Regeneration After Optic Nerve Injury. Stem Cells 2018, 36, 844–855. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Chang, K.-C.; Nahmou, M.; Goldberg, J.L. Induced Pluripotent Stem Cells Promote Retinal Ganglion Cell Survival After Transplant. Invest. Ophthalmol. Vis. Sci. 2018, 59, 1571–1576. [Google Scholar] [CrossRef]
- Rocha, P.N.; Plumb, T.J.; Crowley, S.D.; Coffman, T.M. Effector mechanisms in transplant rejection. Immunol. Rev. 2003, 196, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Kinnear, G.; Jones, N.D.; Wood, K.J. Costimulation Blockade. Transplantation 2013, 95, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, J.; Paster, J.; Benichou, G. Allorecognition by T Lymphocytes and Allograft Rejection. Front. Immunol. 2016, 7, 582. [Google Scholar] [CrossRef] [Green Version]
- Odeberg, J.; Piao, J.-H.; Samuelsson, E.-B.; Falci, S.; Åkesson, E. Low immunogenicity of in vitro-expanded human neural cells despite high MHC expression. J. Neuroimmunol. 2005, 161, 1–11. [Google Scholar] [CrossRef]
- Johansson, S.; Price, J.; Modo, M. Effect of Inflammatory Cytokines on Major Histocompatibility Complex Expression and Differentiation of Human Neural Stem/Progenitor Cells. Stem Cells 2008, 26, 2444–2454. [Google Scholar] [CrossRef]
- Liu, J.; Götherström, C.; Forsberg, M.; Samuelsson, E.-B.; Wu, J.; Calzarossa, C.; Hovatta, O.; Sundström, E.; Åkesson, E. Human neural stem/progenitor cells derived from embryonic stem cells and fetal nervous system present differences in immunogenicity and immunomodulatory potentials in vitro. Stem Cell Res. 2013, 10, 325–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, S.K.; Sharma, R.K.; Gupta, A.; Naik, S. Increased Interleukin-10 Production Without Expansion of CD4+CD25+ T-Regulatory Cells in Early Stable Renal Transplant Patients on Calcineurin Inhibitors. Transplantation 2009, 88, 435–441. [Google Scholar] [CrossRef]
- Levitsky, J.; Gallon, L.; Miller, J.; Tambur, A.R.; Leventhal, J.; Flaa, C.; Huang, X.; Sarraj, B.; Wang, E.; Mathew, J.M. Allospecific Regulatory Effects of Sirolimus and Tacrolimus in the Human Mixed Lymphocyte Reaction. Transplantation 2011, 91, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Ishiyama, K.; Tanaka, Y.; Ide, K.; Ohira, M.; Tahara, H.; Abe, T.; Hirata, F.; Morimoto, H.; Hashimoto, S.; et al. Potential Benefit of Mixed Lymphocyte Reaction Assay-based Immune Monitoring After Living Donor Liver Transplantation for Recipients With Autoimmune Hepatitis. Transplant. Proc. 2014, 46, 785–789. [Google Scholar] [CrossRef]
- Ozaki, M.; Iwanami, A.; Nagoshi, N.; Kohyama, J.; Itakura, G.; Iwai, H.; Nishimura, S.; Nishiyama, Y.; Kawabata, S.; Sugai, K.; et al. Evaluation of the immunogenicity of human iPS cell-derived neural stem/progenitor cells in vitro. Stem Cell Res. 2017, 19, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Miyamae, J.; Yagi, H.; Sato, K.; Okano, M.; Nishiya, K.; Katakura, F.; Sakai, M.; Nakayama, T.; Moritomo, T.; Shiina, T. Evaluation of alloreactive T cells based on the degree of MHC incompatibility using flow cytometric mixed lymphocyte reaction assay in dogs. Immunogenetics 2019, 71, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Colls, P.; Blanco, J.; Martínez-Pasarell, O.; Vidal, F.; Egozcue, J.; Márquez, C.; Guitart, M.; Templado, C. Chromosome segregation in a man heterozygous for a pericentric inversion, inv(9)(p11q13), analyzed by using sperm karyotyping and two-color fluorescence in situ hybridization on sperm nuclei. Hum. Genet. 1997, 99, 761–765. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Kim, B.Y.; Yu, J.E. De novo pericentric inversion of chromosome 9 in congenital anomaly. Yonsei Med. J. 2010, 51, 775–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barres, B.A.; Silverstein, B.E.; Corey, D.P.; Chun, L.L.Y. Immunological, morphological, and electrophysiological variation among retinal ganglion cells purified by panning. Neuron 1988, 1, 791–803. [Google Scholar] [CrossRef]
- Kobayashi, W.; Onishi, A.; Tu, H.-Y.; Takihara, Y.; Matsumura, M.; Tsujimoto, K.; Inatani, M.; Nakazawa, T.; Takahashi, M. Culture Systems of Dissociated Mouse and Human Pluripotent Stem Cell–Derived Retinal Ganglion Cells Purified by Two-Step Immunopanning. Invest. Ophthalmol. Vis. Sci. 2018, 59, 776–787. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Ferguson, T.A. The role of fas ligand in immune privilege. Nat. Rev. Mol. Cell Biol. 2001, 2, 917–924. [Google Scholar] [CrossRef]
- Sharpe, A.H.; Freeman, G.J. The B7–CD28 superfamily. Nat. Rev. Immunol. 2002, 2, 116–126. [Google Scholar] [CrossRef]
- Schoenborn, J.R.; Wilson, C.B. Regulation of Interferon-γ During Innate and Adaptive Immune Responses. Adv. Immunol. 2007, 96, 41–101. [Google Scholar]
- Fujii, S.; Yoshida, S.; Inagaki, E.; Hatou, S.; Tsubota, K.; Takahashi, M.; Shimmura, S.; Sugita, S. Immunological Properties of Neural Crest Cells Derived from Human Induced Pluripotent Stem Cells. Stem Cells Dev. 2019, 28, 28–43. [Google Scholar] [CrossRef]
- Gao, W.; Demirci, G.; Strom, T.B.; Li, X.C. Stimulating PD-1-negative signals concurrent with blocking CD154 co-stimulation induces long-term islet allograft survival. Transplantation 2003, 76, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Ozkaynak, E.; Wang, L.; Goodearl, A.; McDonald, K.; Qin, S.; O’Keefe, T.; Duong, T.; Smith, T.; Gutierrez-Ramos, J.C.; Rottman, J.B.; et al. Programmed Death-1 Targeting Can Promote Allograft Survival. J. Immunol. 2002, 169, 6546–6553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, S.; Sugita, S.; Futagami, Y.; Kawaguchi, T.; Kamoi, K.; Shirato, S.; Mochizuki, M. Human iris pigment epithelium suppresses activation of bystander T cells via TGFβ–TGFβ receptor interaction. Exp. Eye Res. 2009, 88, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Sugita, S.; Horie, S.; Yamagami, S.; Mochizuki, M. Mechanisms of Immune Suppression for CD8+T Cells by Human Corneal Endothelial Cells via Membrane-Bound TGFβ. Invest. Ophthalmol. Vis. Sci. 2010, 51, 2548–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheibani, N.; Sorenson, C.M.; Cornelius, L.A.; Frazier, W.A. Thrombospondin-1, a Natural Inhibitor of Angiogenesis, Is Present in Vitreous and Aqueous Humor and Is Modulated by Hyperglycemia. Biochem. Biophys. Res. Commun. 2000, 267, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Lawler, J. The functions of thrombospondin-1 and-2. Curr. Opin. Cell Biol. 2000, 12, 634–640. [Google Scholar] [CrossRef]
- Taylor, A.W.; Yee, D.G. Somatostatin Is an Immunosuppressive Factor in Aqueous Humor. Invest. Ophthalmol. Vis. Sci. 2003, 44, 2644–2649. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.W. A review of the influence of aqueous humor on immunity. Ocul. Immunol. Inflamm. 2003, 11, 231–241. [Google Scholar] [CrossRef]
- Streilein, J.W. Ocular immune privilege: Therapeutic opportunities from an experiment of nature. Nat. Rev. Immunol. 2003, 3, 879–889. [Google Scholar] [CrossRef]
- Stein-Streilein, J. Mechanisms of immune privilege in the posterior eye. Int. Rev. Immunol. 2013, 32, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Murphy-Ullrich, J.E.; Suto, M.J. Thrombospondin-1 regulation of latent TGF-β activation: A therapeutic target for fibrotic disease. Matrix Biol. 2018, 68–69, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Wenkel, H.; Streilein, J.W. Analysis of immune deviation elicited by antigens injected into the subretinal space. Invest. Ophthalmol. Vis. Sci. 1998, 39, 1823–1834. [Google Scholar]
- Sonoda, K.-H.; Sakamoto, T.; Qiao, H.; Hisatomi, T.; Oshima, T.; Tsutsumi-Miyahara, C.; Exley, M.; Balk, S.P.; Taniguchi, M.; Ishibashi, T. The analysis of systemic tolerance elicited by antigen inoculation into the vitreous cavity: Vitreous cavity-associated immune deviation. Immunology 2005, 116, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; Makabe, K.; Fujii, S.; Iwasaki, Y.; Kamao, H.; Shiina, T.; Ogasawara, K.; Takahashi, M. Detection of Retinal Pigment Epithelium-Specific Antibody in iPSC-Derived Retinal Pigment Epithelium Transplantation Models. Stem Cell Rep. 2017, 9, 1501–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGill, T.J.; Stoddard, J.; Renner, L.M.; Messaoudi, I.; Bharti, K.; Mitalipov, S.; Lauer, A.; Wilson, D.J.; Neuringer, M. Allogeneic iPSC-Derived RPE Cell Graft Failure Following Transplantation Into the Subretinal Space in Nonhuman Primates. Invest. Ophthalmol. Vis. Sci. 2018, 59, 1374–1383. [Google Scholar] [CrossRef] [Green Version]
- Fujii, S.; Sugita, S.; Futatsugi, Y.; Ishida, M.; Edo, A.; Makabe, K.; Kamao, H.; Iwasaki, Y.; Sakaguchi, H.; Hirami, Y.; et al. A Strategy for Personalized Treatment of iPS-Retinal Immune Rejections Assessed in Cynomolgus Monkey Models. Int. J. Mol. Sci. 2020, 21, 3077. [Google Scholar] [CrossRef]
- Sugita, S.; Iwasaki, Y.; Makabe, K.; Kamao, H.; Mandai, M.; Shiina, T.; Ogasawara, K.; Hirami, Y.; Kurimoto, Y.; Takahashi, M. Successful Transplantation of Retinal Pigment Epithelial Cells from MHC Homozygote iPSCs in MHC-Matched Models. Stem Cell Rep. 2016, 7, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Okita, K.; Yamakawa, T.; Matsumura, Y.; Sato, Y.; Amano, N.; Watanabe, A.; Goshima, N.; Yamanaka, S. An Efficient Nonviral Method to Generate Integration-Free Human-Induced Pluripotent Stem Cells from Cord Blood and Peripheral Blood Cells. Stem Cells 2013, 31, 458–466. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, M.; Taniguchi, Y.; Senda, S.; Takizawa, N.; Ichisaka, T.; Asano, K.; Morizane, A.; Doi, D.; Takahashi, J.; Nishizawa, M.; et al. A novel efficient feeder-free culture system for the derivation of human induced pluripotent stem cells. Sci. Rep. 2014, 4, 3594. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Gene Description | Abbreviations | Signal Log2 Ratio | ||
|---|---|---|---|---|---|
| Signal in iPSC-RGCs | vs. iPSCs | vs. PBMCs | |||
| MHC Class I and Class II | |||||
| NM_001242758 | major histocompatibility complex, class I, A | HLA-A | 56.74 | −2.60 | −7.91 |
| ENST00000412585 | major histocompatibility complex, class I, B | HLA-B | 130.96 | −2.00 | −8.48 |
| NM_001243042 | major histocompatibility complex, class I, C | HLA-C | 134.83 | −3.20 | −8.04 |
| NM_001242525 | major histocompatibility complex, class II, DP alpha 1 | HLA-DPA1 | 64.49 | −0.79 | −8.87 |
| NM_002121 | major histocompatibility complex, class II, DP beta 1 | HLA-DPB1 | 71.71 | −3.08 | −9.16 |
| NM_002122 | major histocompatibility complex, class II, DQ alpha 1 | HLA-DQA1 | 30.59 | −0.11 | −9.89 |
| OTTHUMT00000076179 | major histocompatibility complex, class II, DQ alpha 2 | HLA-DQA2 | 69.39 | 0.01 | −5.88 |
| NM_001243961 | major histocompatibility complex, class II, DQ beta 1 | HLA-DQB1 | 44.16 | −0.86 | −6.94 |
| ENST00000411527 | major histocompatibility complex, class II, DQ beta 2 | HLA-DQB2 | 223.91 | −0.52 | −2.14 |
| NM_019111 | major histocompatibility complex, class II, DR alpha | HLA-DRA | 34.59 | −1.70 | −11.94 |
| NM_001243965 | major histocompatibility complex, class II, DR beta 1 | HLA-DRB1 | 44.85 | −3.81 | −10.23 |
| NM_002125 | major histocompatibility complex, class II, DR beta 5 | HLA-DRB5 | 30.10 | −0.98 | −10.77 |
| Co-stimulatory Molecules | |||||
| NM_001250 | CD40 molecule, TNF receptor superfamily member 5 | CD40 | 29.43 | −2.27 | −5.70 |
| NM_005191 | CD80 molecule | CD80/B7-1 | 23.80 | 0.06 | −4.26 |
| NM_001206924 | CD86 molecule | CD86/B7-2 | 12.31 | −0.53 | −6.30 |
| NM_001267706 | CD274 molecule | CD274/B7-H1/PD-L1 | 149.33 | 2.33 | −4.57 |
| NM_001024736 | CD276 molecule | CD276/B7-H3 | 473.67 | −0.96 | 2.16 |
| NM_025239 | programmed cell death 1 ligand 2 | PDCD1LG2/PD-L2/B7-DC | 26.47 | 1.13 | −3.39 |
| NM_001283050 | inducible T-cell co-stimulator ligand | ICOSLG/B7-H2 | 61.92 | 0.00 | 0.00 |
| NM_001253849 | V-set domain containing T-cell activation inhibitor 1 | VTCN1/B7-H4 | 39.01 | 0.42 | 0.49 |
| NM_022153 | chromosome 10 open reading frame 54 | C10orf54/VISTA/B7-H5 | 67.94 | 0.00 | −0.77 |
| NM_003327 | tumor necrosis factor receptor superfamily, member 4 | TNFRSF4/OX40L | 26.44 | 0.68 | −2.68 |
| NM_001252 | CD70 molecule | CD70 | 13.67 | 0.19 | −4.81 |
| NM_001297605 | tumor necrosis factor receptor superfamily, member 14 | TNFRSF14/HVEM | 75.79 | 0.51 | −2.90 |
| NM_005092 | tumor necrosis factor (ligand) superfamily, member 18 | TNFSF18/GITRL | 9.20 | −0.40 | −0.55 |
| NM_003811 | tumor necrosis factor (ligand) superfamily, member 9 | TNFSF9/4-1BBL/CD137L | 240.76 | −0.15 | −0.15 |
| Cytokines and Other Inhibitory Factors/Molecules | |||||
| NM_000660 | transforming growth factor beta 1 | TGFB1 | 453.15 | 2.71 | −3.85 |
| NM_001135599 | transforming growth factor beta 2 | TGFB2 | 20563.82 | 3.85 | 7.86 |
| NM_003239 | transforming growth factor beta 3 | TGFB3 | 30.27 | 0.22 | 0.16 |
| NM_000639 | Fas ligand (TNF superfamily, member 6) | FASLG | 24.71 | −0.24 | −2.87 |
| NM_004878 | prostaglandin E synthase | PTGES | 171.18 | 0.52 | 0.67 |
| NM_001256335 | prostaglandin E synthase 2 | PTGES2 | 234.85 | −0.57 | −0.54 |
| NM_000572 | interleukin 10 | IL10 | 19.01 | −0.20 | −2.77 |
| NM_002164 | indoleamine 2,3-dioxygenase 1 | IDO1 | 37.34 | −4.22 | −8.23 |
| NM_194294 | indoleamine 2,3-dioxygenase 2 | IDO2 | 43.79 | −0.31 | −0.31 |
| NM_003381 | vasoactive intestinal peptide | VIP | 25.96 | 0.19 | 0.00 |
| NM_002415 | macrophage migration inhibitory factor | MIF | 6137.52 | −0.89 | −0.93 |
| NM_002389 | CD46 molecule, complement regulatory protein | CD46 | 3081.43 | −0.78 | −0.35 |
| NM_000574 | CD55 molecule, decay accelerating factor for complement | CD55 | 2793.51 | −1.37 | −1.95 |
| NM_000611 | CD59 molecule, complement regulatory protein | CD59 | 10117.89 | 1.88 | 1.49 |
| NM_003246 | thrombospondin 1 | THBS1/TSP-1 | 10138.58 | 5.59 | 7.24 |
| NM_001048 | somatostatin | SST | 959.93 | 5.88 | 5.90 |
| NM_000577 | interleukin 1 receptor antagonist | IL1RN | 17.64 | 0.26 | −5.78 |
| NM_001777 | CD47 molecule | CD47 | 3346.94 | 1.88 | −1.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edo, A.; Sugita, S.; Futatsugi, Y.; Sho, J.; Onishi, A.; Kiuchi, Y.; Takahashi, M. Capacity of Retinal Ganglion Cells Derived from Human Induced Pluripotent Stem Cells to Suppress T-Cells. Int. J. Mol. Sci. 2020, 21, 7831. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217831
Edo A, Sugita S, Futatsugi Y, Sho J, Onishi A, Kiuchi Y, Takahashi M. Capacity of Retinal Ganglion Cells Derived from Human Induced Pluripotent Stem Cells to Suppress T-Cells. International Journal of Molecular Sciences. 2020; 21(21):7831. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217831
Chicago/Turabian StyleEdo, Ayaka, Sunao Sugita, Yoko Futatsugi, Junki Sho, Akishi Onishi, Yoshiaki Kiuchi, and Masayo Takahashi. 2020. "Capacity of Retinal Ganglion Cells Derived from Human Induced Pluripotent Stem Cells to Suppress T-Cells" International Journal of Molecular Sciences 21, no. 21: 7831. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217831