Protective Effects of Colomast®, a New Formulation of Adelmidrol and Sodium Hyaluronate, in a Mouse Model of Acute Restraint Stress

 , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. In Vivo Adelmidrol Absorption
2.2. Effect of Colomast® Pre-Treatment on Stress Hormones and Fecal Output
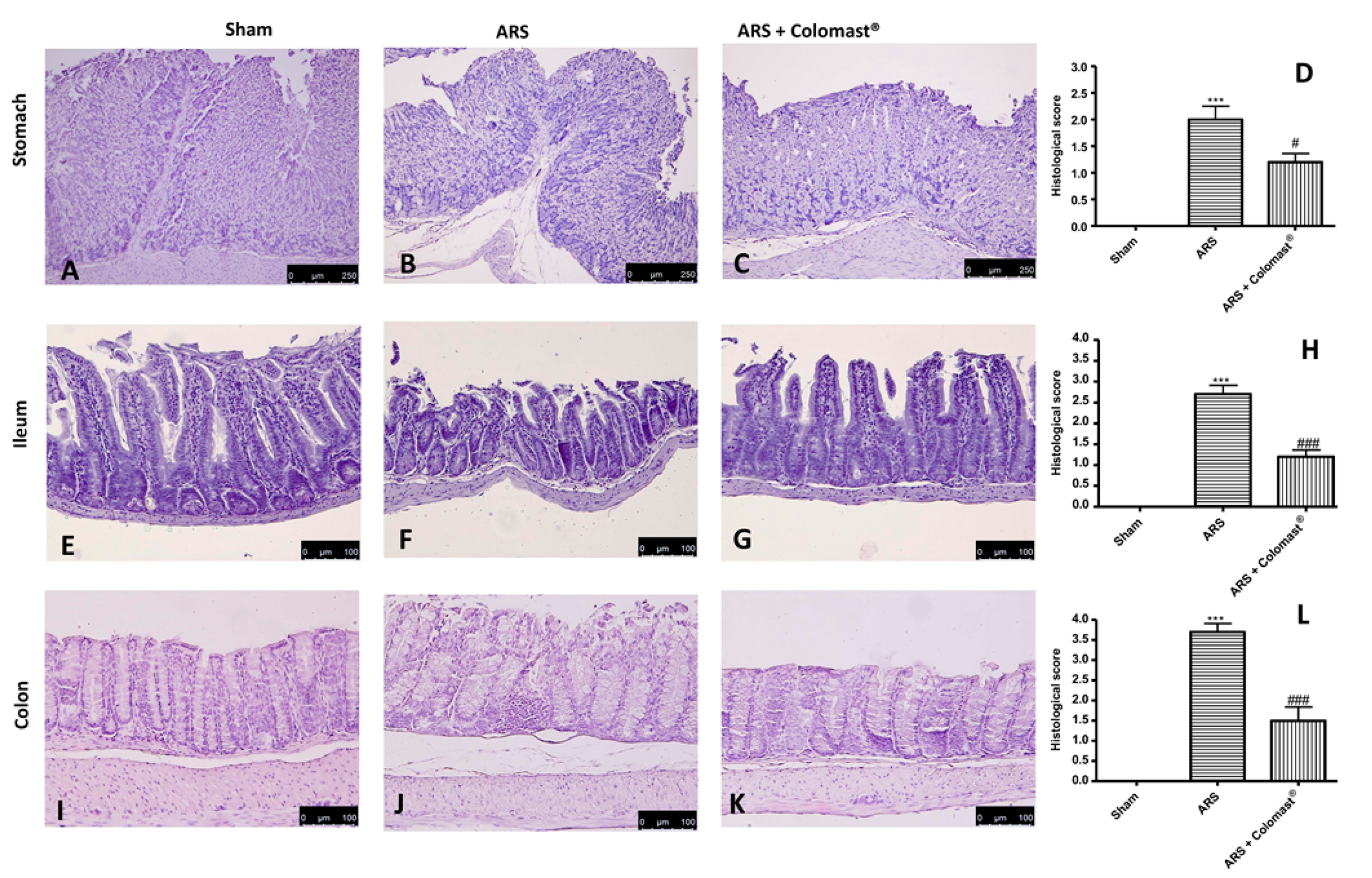
2.3. Effect of Colomast® Pre-Treatment on Tissue Damage
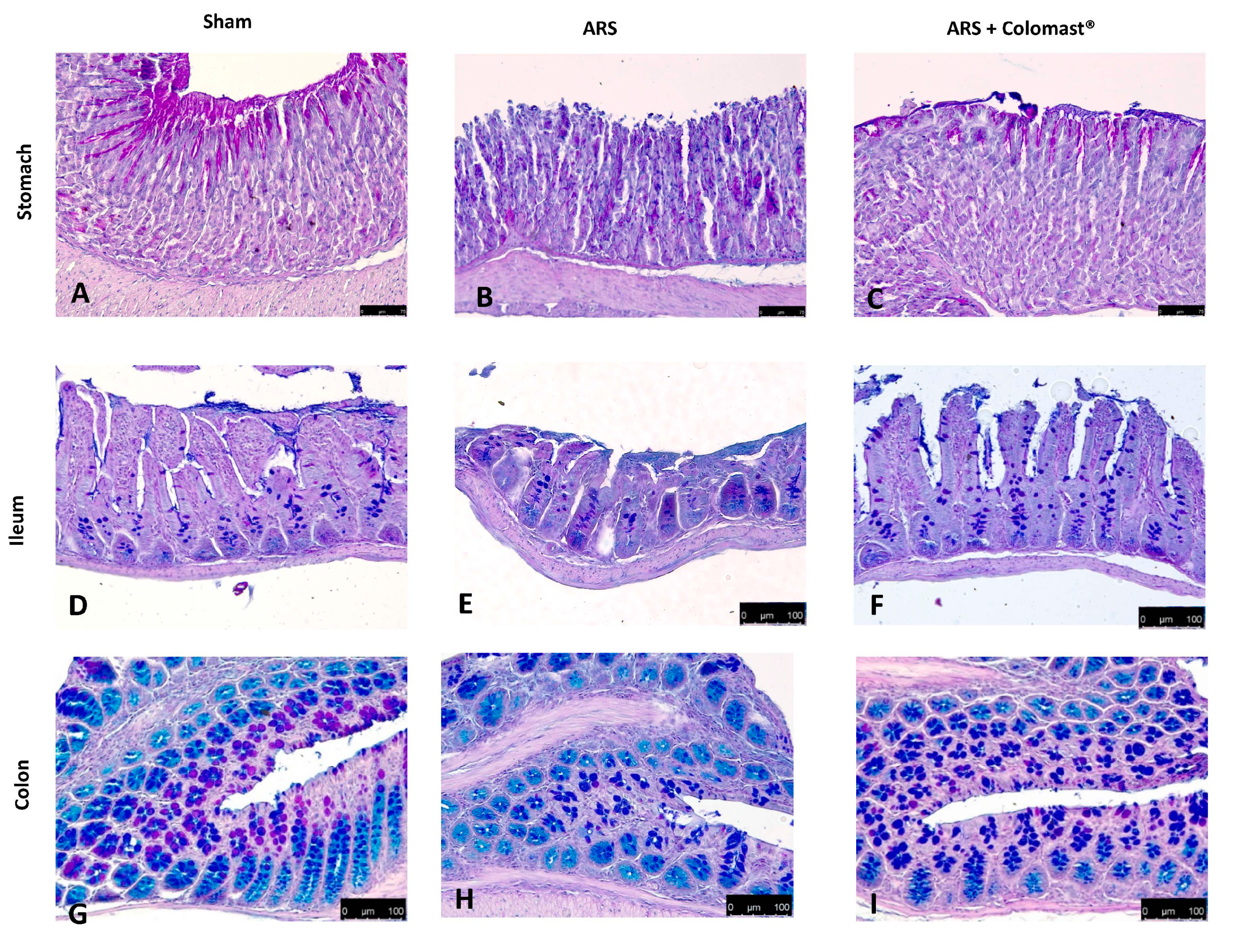
2.4. Effect of Colomast® Pre-Treatment on Mucous Secreting Cells
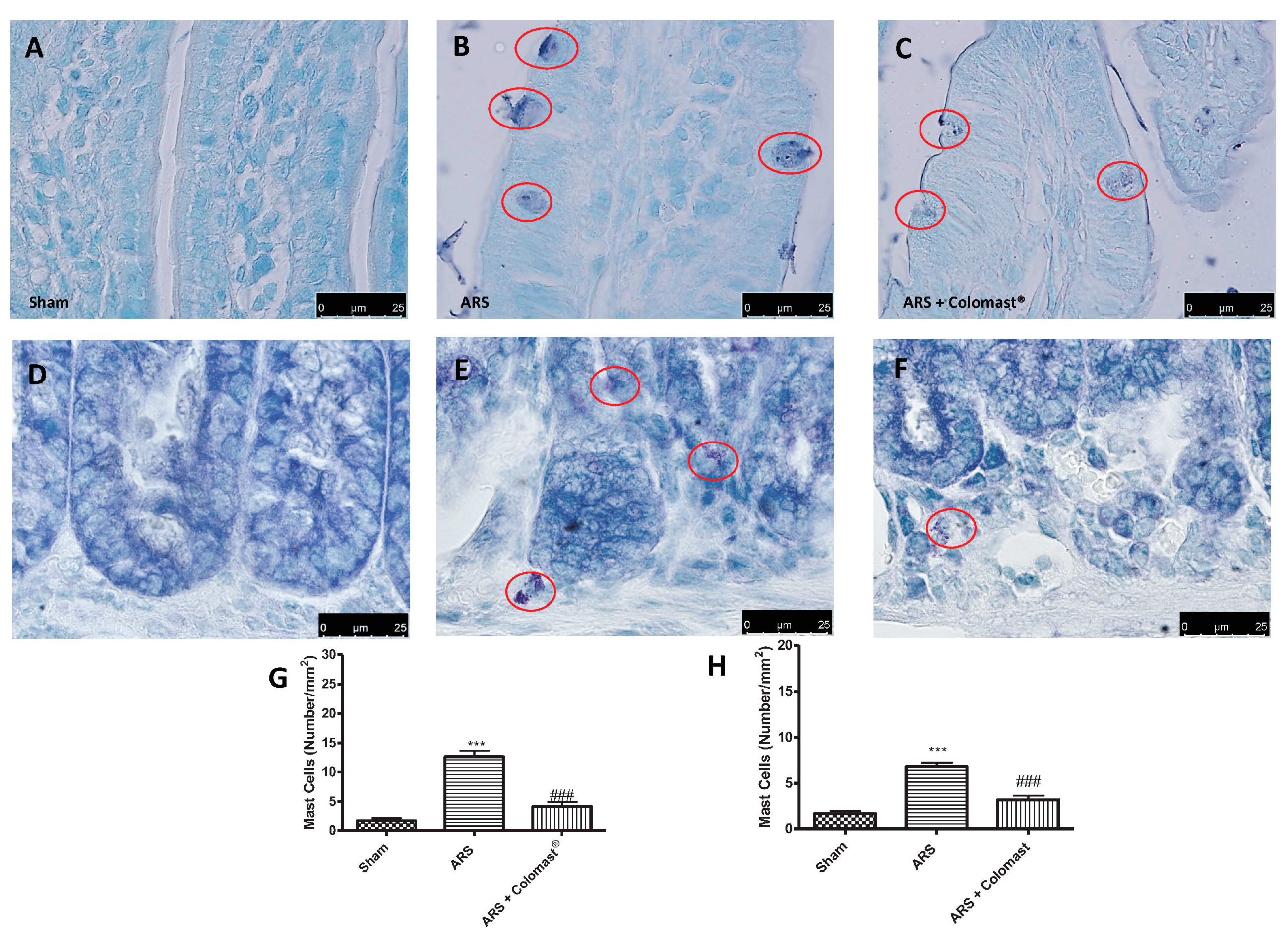
2.5. Effect of Colomast® Pre-Treatment on Ars-Induced Mast Cell Degranulation in Ileum and Colon
2.6. Effect of Colomast® Pre-Treatment on Inflammation Pathway
2.7. Effect of Colomast® Pre-Treatment on Nitrotyrosine Formation, Poly (Adp-Ribose) Polymerase (Parp) Activation, Myeloperoxidase Activity (Mpo), and Lipid Peroxidation in Ileum
2.8. Effect of Colomast® Pre-Treatment on Apoptotic Damage in Ileum Tissue
2.9. Effect of Colomast® Pre-Treatment on Tight Junction Expression in Ileum
2.10. Effect of Colomast® Pre-Treatment on Tj Expression in Brain
2.11. Effect of Colomast® Pre-Treatment on Brain Tissue Damage
2.12. Effect of Colomast® Pre-Treatment on Cell Death and Cellular Proliferation in Hippocampus
2.13. Effect of Colomast® Pre-Treatment on Behavioral Alteration and Sucrose Consumption (%)
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Colomast® Composition and Measurement of Adelmidrol Plasma Absorption Rate by Lc-Ms/Ms (Preliminary Data)
4.3. Animal Model of Restraint Stress
4.4. Experimental Groups
- ARS + Veh: mice were immobilized as described above and vehicle CMC 2.5% was administered;
- ARS + Colomast®: mice were immobilized and Colomast® (20 mg/kg) in CMC 2.5% was administered by oral gavage 30 min before the immobilization.
- Sham + Veh: mice were only briefly anesthetized with isoflurane and thereafter allowed to move freely in their cages over the following 2 h.
- Sham + Colomast®: the same conditions of Sham+Veh group, but Colomast® (20 mg/kg) in CMC 2.5% was administered by oral gavage (data not shown).
4.5. Measurement of Hormones Stress
4.6. Histological Examination
4.7. Alcian Blue/PAS Staining
4.8. Staining of Mast Cells
4.9. Myeloperoxidase Activity
4.10. Western Blot Analysis of Cytosolic and Nuclear Extracts from Ileum Tissue
4.11. Immunohistochemical Localization of iNOS, Nitrotyrosine, Poly (ADP-ribose) Polymerase (PARP)
4.12. Measurement of Cytokines
4.13. Thiobarbituric Acid-Reactant Substances Measurement (MDA Levels)
4.14. Terminal Deoxynucleotidyl Nick-End Labeling (TUNEL) Assay
4.15. Immunofluorescence Localization of ZO-1, Occludin, KI-67
4.16. Behavioral Testing
4.17. Sucrose Consumption (%)
4.18. Materials
4.19. Statistical Evaluation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTH | adrenocorticotrophic hormone |
| ALIAmide | autacoid local injury antagonist amide |
| ARS | acute restraint stress |
| BBB | blood brain barrier |
| CNS | central nervous system |
| CORT | corticosterone |
| CRH | hypothalamic corticotrophin-releasing hormone |
| EPM | elevated plus-maze |
| FST | force swimming test |
| GI | gastrointestinal |
| HPA | hypothalamic-pituitary-adrenal axis |
| IL-1β | interleukin-1β |
| MPO | myeloperoxidase |
| OF | open field |
| PARP | poly (ADP-ribose) polymerase |
| PEA | palmitoylethanolamide |
| TJ | tight junction |
| TNF-α | tumor necrosis factor alpha |
| TUNEL | terminal deoxynucleotidyl transferase dUTP nick end labeling assay |
| ZO-1 | zonula occludens-1 (ZO-1 |
References
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialog. Clin. Neurosci. 2006, 8, 383–395. [Google Scholar]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Central effects of stress hormones in health and disease: Understanding the protective and damaging effects of stress and stress mediators. Eur. J. Pharmacol. 2008, 583, 174–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madrigal, J.L.M.; Hurtado, O.; Moro, M.A.; Lizasoain, I.; Lorenzo, P.; Castrillo, A.; Bosca, L.; Leza, J.C. The increase in TNF-α levels is implicated in NF-κB activation and inducible nitric oxide synthase expression in brain cortex after immobilization stress. Neuropsychopharmacology 2002, 26, 155–163. [Google Scholar] [CrossRef]
- Sugama, S. Stress-induced microglial activation may facilitate the progression of neurodegenerative disorders. Med. Hypotheses 2009, 73, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, X.; Shigenaga, M.K.; Yeo, H.C.; Mori, A.; Ames, B.N. Immobilization stress causes oxidative damage to lipid, protein, and DNA in the brain of rats. FASEB J. 1996, 10, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.T.; Choi, J.H.; Chang, J.W.; Kim, S.W.; Hwang, O. Immobilization stress causes increases in tetrahydrobiopterin, dopamine, and neuromelanin and oxidative damage in the nigrostriatal system. J. Neurochem. 2005, 95, 89–98. [Google Scholar] [CrossRef]
- Sudo, N.; Oyama, N.; Yu, X.N.; Kubo, C. Restraint stress-induced elevation of endogenous glucocorticoids decreases Peyer’s patch cell numbers via mechanisms that are either dependent or independent on apoptotic cell death. Neuroimmunomodulation 2001, 9, 333–339. [Google Scholar] [CrossRef]
- Emerit, J.; Edeas, M.; Bricaire, F. Neurodegenerative diseases and oxidative stress. Eur. Neuropsychopharmacol. 2004, 15, S100–S101. [Google Scholar] [CrossRef]
- Farooqui, T.; Farooqui, A.A. Lipid-mediated oxidative stress and inflammation in the pathogenesis of Parkinson’s disease. Parkinsons Dis. 2011, 2011, 247467. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.B.S.; Gu, H.; Mitchell, T.D.; Endale, L.; Russo, M.; Ryan, D.H. Increased glucocorticoid response to a novel stress in rats that have been restrained. Physiol. Behav. 2004, 81, 557–568. [Google Scholar] [CrossRef]
- Saunders, P.R.; Hanssen, N.P.; Perdue, M.H. Cholinergic nerves mediate stress-induced intestinal transport abnormalities in Wistar-Kyoto rats. Am. J. Physiol. 1997, 273, G486–G490. [Google Scholar] [CrossRef]
- Saunders, P.R.; Kosecka, U.; McKay, D.M.; Perdue, M.H. Acute stressors stimulate ion secretion and increase epithelial permeability in rat intestine. Am. J. Physiol. Liver Physiol. 1994, 267, G794–G799. [Google Scholar] [CrossRef]
- Mazzon, E.; Crisafulli, C.; Galuppo, M.; Cuzzocrea, S. Role of peroxisome proliferator-activated receptor-α in ileum tight junction alteration in mouse model of restraint stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G488–G505. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Di Paola, R.; Cordaro, M.; Gugliandolo, E.; Casili, G.; Morittu, V.M.; Britti, D.; Esposito, E.; Cuzzocrea, S. Adelmidrol, a palmitoylethanolamide analogue, as a new pharmacological treatment for the management of acute and chronic inflammation. Biochem. Pharmacol. 2016, 119, 27–41. [Google Scholar] [CrossRef]
- Ostardo, E.; Impellizzeri, D.; Cervigni, M.; Porru, D.; Sommariva, M.; Cordaro, M.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; Crupi, R.; et al. Adelmidrol + sodium hyaluronate in IC/BPS or conditions associated to chronic urothelial inflammation. A translational study. Pharmacol. Res. 2018, 134, 16–30. [Google Scholar] [CrossRef]
- Cerrato, S.; Brazis, P.; Della Valle, M.F.; Miolo, A.; Puigdemont, A. Inhibitory effect of topical Adelmidrol on antigen-induced skin wheal and mast cell behavior in a canine model of allergic dermatitis. BMC Veter. Res. 2012, 8, 230. [Google Scholar] [CrossRef] [Green Version]
- Pulvirenti, N.; Nasca, M.R.; Micali, G. Topical adelmidrol 2% emulsion, a novel aliamide, in the treatment of mild atopic dermatitis in pediatric subjects: A pilot study. Acta Dermatovenerol. Croat. ADC 2007, 15, 80–83. [Google Scholar] [PubMed]
- Fusco, R.; Cordaro, M.; Genovese, T.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Peritore, A.F.; D’Amico, R.; Crupi, R.; Cuzzocrea, S.; et al. Adelmidrol: A new promising antioxidant and anti-inflammatory therapeutic tool in pulmonary fibrosis. Antioxidants 2020, 9, 601. [Google Scholar] [CrossRef]
- De Filippis, D.; D’Amico, A.; Cinelli, M.P.; Esposito, G.; Di Marzo, V.; Iuvone, T. Adelmidrol, a palmitoylethanolamide analogue, reduces chronic inflammation in a carrageenin-granuloma model in rats. J. Cell. Mol. Med. 2009, 13, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Puigdemont, A.; Della Valle, M.; Fusco, M.; Verde, R.; Allarà, M.; Aveta, T.; Orlando, P.; Di Marzo, V. Adelmidrol increases the endogenous concentrations of palmitoylethanolamide in canine keratinocytes and down-regulates an inflammatory reaction in an in vitro model of contact allergic dermatitis. Veter. J. 2016, 207, 85–91. [Google Scholar] [CrossRef]
- Di Paola, R.; Fusco, R.; Impellizzeri, D.; Cordaro, M.; Britti, D.; Morittu, V.M.; Evangelista, M.; Cuzzocrea, S. Adelmidrol, in combination with hyaluronic acid, displays increased anti-inflammatory and analgesic effects against monosodium iodoacetate-induced osteoarthritis in rats. Arthritis Res. 2016, 18, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campolo, M.; Siracusa, R.; Cordaro, M.; Filippone, A.; Gugliandolo, E.; Peritore, A.F.; Impellizzeri, D.; Crupi, R.; Paterniti, I.; Cuzzocrea, S. The association of adelmidrol with sodium hyaluronate displays beneficial properties against bladder changes following spinal cord injury in mice. PLoS ONE 2019, 14, e0208730. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.; Yang, P.-C.; Söderholm, J.D.; Benjamin, M.; Perdue, M.H. Role of mast cells in chronic stress induced colonic epithelial barrier dysfunction in the rat. Gut 2001, 48, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Fink, M.P.; Delude, R.L. Epithelial barrier dysfunction: A unifying theme to explain the pathogenesis of multiple organ dysfunction at the cellular level. Crit. Care Clin. 2005, 21, 177–196. [Google Scholar] [CrossRef]
- Jacob, C.; Yang, P.-C.; Darmoul, D.; Amadesi, S.; Saito, T.; Cottrell, G.S.; Coelho, A.M.; Singh, P.; Grady, E.F.; Perdue, M.; et al. Mast cell tryptase controls paracellular permeability of the intestine. Role of protease-activated receptor 2 and beta-arrestins. J. Biol. Chem. 2005, 280, 31936–31948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and function of the blood–brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell Physiol. 2000, 182, 311–322. [Google Scholar] [CrossRef]
- Wilder, R.L. Neuroendocrine-immune system interactions and autoimmunity. Annu. Rev. Immunol. 1995, 13, 307–338. [Google Scholar] [CrossRef]
- Sapolsky, R.M. Why stress is bad for your brain. Science 1996, 273, 749–750. [Google Scholar] [CrossRef]
- Szabo, S. Hans selye 70 years later: Steroids, stress ulcers & H. pylori. Ideggyogy. Sz. 2014, 67, 91–94. [Google Scholar]
- Buechel, H.M.; Epopovic, J.; Staggs, K.H.; Anderson, K.L.; Thibault, O.; Blalock, E.M. Aged rats are hypo-responsive to acute restraint: Implications for psychosocial stress in aging. Front. Aging Neurosci. 2014, 6, 13. [Google Scholar] [CrossRef]
- Cohen, S.; Janicki-Deverts, D.; Miller, G.E. Psychological stress and disease. JAMA 2007, 298, 1685–1687. [Google Scholar] [CrossRef]
- Magarinos, A.M.; Verdugo, J.M.G.; McEwen, B.S. Chronic stress alters synaptic terminal structure in hippocampus. Proc. Natl. Acad. Sci. USA 1997, 94, 14002–14008. [Google Scholar] [CrossRef] [Green Version]
- Vuong, H.E.; Yano, J.M.; Fung, T.C.; Hsiao, E.Y. The microbiome and host behavior. Annu. Rev. Neurosci. 2017, 40, 21–49. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Microbes, immunity, and behavior: Psychoneuroimmunology meets the microbiome. Neuropsychopharmacology 2016, 42, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef]
- Matsumoto, K.; Yobimoto, K.; Huong, N.T.T.; Abdel-Fattah, M.; Van Hien, T.; Watanabe, H. Psychological stress-induced enhancement of brain lipid peroxidation via nitric oxide systems and its modulation by anxiolytic and anxiogenic drugs in mice. Brain Res. 1999, 839, 74–84. [Google Scholar] [CrossRef]
- Mastrofrancesco, A.; Ottaviani, M.; Aspite, N.; Cardinali, G.; Izzo, E.; Graupe, K.; Zouboulis, C.C.; Camera, E.; Picardo, M. Azelaic acid modulates the inflammatory response in normal human keratinocytes through PPARgamma activation. Exp. Dermatol. 2010, 19, 813–820. [Google Scholar] [CrossRef]
- Siracusa, R.; Impellizzeri, D.; Cordaro, M.; Gugliandolo, E.; Peritore, A.F.; Di Paola, R.; Cuzzocrea, S. Topical application of adelmidrol + trans-traumatic acid enhances skin wound healing in a streptozotocin-induced diabetic mouse model. Front. Pharmacol. 2018, 9, 871. [Google Scholar] [CrossRef]
- Cordaro, M.; Impellizzeri, D.; Gugliandolo, E.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. Adelmidrol, a palmitoylethanolamide analogue, as a new pharmacological treatment for the management of inflammatory bowel disease. Mol. Pharmacol. 2016, 90, 549–561. [Google Scholar] [CrossRef] [Green Version]
- Aloe, L.; Leon, A.; Levi-Montalcini, R. A proposed autacoid mechanism controlling mastocyte behaviour. Agents Actions 1993, 39, C145–C147. [Google Scholar] [CrossRef]
- De Filippis, D.; Luongo, L.; Cipriano, M.; Palazzo, E.; Cinelli, M.P.; De Novellis, V.; Maione, S.; Iuvone, T. Palmitoylethanolamide reduces granuloma-induced hyperalgesia by modulation of mast cell activation in Rats. Mol. Pain 2011, 7, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suda, K.; Setoyama, H.; Nanno, M.; Matsumoto, S.; Kawai, M. Involvement of parasympathetic pelvic efferent pathway in psychological stress-induced defecation. World J. Gastroenterol. 2013, 19, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.Y.; Chen, X.N.; Tong, D.L.; Zhou, J.N. Stress and glucocorticoids regulated corticotropin releasing factor in rat prefrontal cortex. Mol. Cell. Endocrinol. 2011, 342, 54–63. [Google Scholar] [CrossRef]
- Han, X.; Fink, M.P.; Uchiyama, T.; Yang, R.; Delude, R.L. Increased iNOS activity is essential for hepatic epithelial tight junction dysfunction in endotoxemic mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G126–G136. [Google Scholar] [CrossRef]
- Yang, R.; Harada, T.; Li, J.; Uchiyama, T.; Han, Y.; Englert, J.A.; Fink, M.P. Bile modulates intestinal epithelial barrier function via an extracellular signal related kinase 1/2 dependent mechanism. Intensive Care Med. 2005, 31, 709–717. [Google Scholar] [CrossRef]
- Zheng, G.; Fon, G.V.; Meixner, W.; Creekmore, A.; Zong, Y.; Dame, M.K.; Colacino, J.; Dedhia, P.H.; Hong, S.; Wiley, J.W. Chronic stress and intestinal barrier dysfunction: Glucocorticoid receptor and transcription repressor HES1 regulate tight junction protein Claudin-1 promoter. Sci. Rep. 2017, 7, 4502. [Google Scholar] [CrossRef] [Green Version]
- Szabo, C.; Zingarelli, B.; O’Connor, M.; Salzman, A.L. DNA strand breakage, activation of poly (ADP-ribose) synthetase, and cellular energy depletion are involved in the cytotoxicity of macrophages and smooth muscle cells exposed to peroxynitrite. Proc. Natl. Acad. Sci. USA 1996, 93, 1753–1758. [Google Scholar] [CrossRef] [Green Version]
- Szabó, É.; Virág, L.; Bakondi, E.; Gyüre, L.; Haskó, G.; Bai, P.; Hunyadi, J.; Gergely, P.; Szabo, C. Peroxynitrite production, DNA breakage, and poly(ADP-ribose) polymerase activation in a mouse model of oxazolone-induced contact hypersensitivity. J. Investig. Dermatol. 2001, 117, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Khalaj, L.; Nejad, S.C.; Mohammadi, M.; Zadeh, S.S.; Pour, M.H.; Ahmadiani, A.; Khodagholi, F.; Ashabi, G.; Alamdary, S.Z.; Samami, E. Gemfibrozil pretreatment proved protection against acute restraint stress-induced changes in the male rats’ hippocampus. Brain Res. 2013, 1527, 117–130. [Google Scholar] [CrossRef]
- Mazzon, E.; Cuzzocrea, S. Role of TNF-α in ileum tight junction alteration in mouse model of restraint stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1268–G1280. [Google Scholar] [CrossRef]
- Abbott, N.J. Blood–brain barrier structure and function and the challenges for CNS drug delivery. J. Inherit. Metab. Dis. 2013, 36, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Wolburg, H.; Noell, S.; Mack, A.; Wolburg-Buchholz, K.; Fallier-Becker, P. Brain endothelial cells and the glio-vascular complex. Cell Tissue Res. 2008, 335, 75–96. [Google Scholar] [CrossRef] [PubMed]
- Rosenbrock, H.; Koros, E.; Bloching, A.; Podhorna, J.; Borsini, F. Effect of chronic intermittent restraint stress on hippocampal expression of marker proteins for synaptic plasticity and progenitor cell proliferation in rats. Brain Res. 2005, 1040, 55–63. [Google Scholar] [CrossRef]
- Wu, H.H.; Wang, S. Strain differences in the chronic mild stress animal model of depression. Behav. Brain Res. 2010, 213, 94–102. [Google Scholar] [CrossRef]
- Caraci, F.; Calabrese, F.; Molteni, R.; Bartova, L.; Dold, M.; Leggio, G.M.; Fabbri, C.; Mendlewicz, J.; Racagni, G.; Kasper, S.; et al. International union of basic and clinical pharmacology CIV: The neurobiology of treatment-resistant depression: From antidepressant classifications to novel pharmacological targets. Pharmacol. Rev. 2018, 70, 475–504. [Google Scholar] [CrossRef] [Green Version]
- Caraci, F.; Spampinato, S.F.; Morgese, M.G.; Tascedda, F.; Salluzzo, M.G.; Giambirtone, M.C.; Caruso, G.; Munafò, A.; Torrisi, S.A.; Leggio, G.M.; et al. Neurobiological links between depression and AD: The role of TGF-β1 signaling as a new pharmacological target. Pharmacol. Res. 2018, 130, 374–384. [Google Scholar] [CrossRef]
- Petrosino, S.; Moriello, A.S.; Cerrato, S.; Fusco, M.; Puigdemont, A.; De Petrocellis, L.; Di Marzo, V. The anti-inflammatory mediator palmitoylethanolamide enhances the levels of 2-arachidonoyl-glycerol and potentiates its actions at TRPV1 cation channels. Br. J. Pharmacol. 2015, 173, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Cordaro, M.; Scuto, M.; Siracusa, R.; D’Amico, R.; Peritore, A.F.; Gugliandolo, E.; Fusco, R.; Crupi, R.; Impellizzeri, D.; Pozzebon, M.; et al. Effect of N-palmitoylethanolamine-oxazoline on comorbid neuropsychiatric disturbance associated with inflammatory bowel disease. FASEB J. 2020, 34, 4085–4106. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, M.; Szymaszkiewicz, A.; Salaga, M.; Zatorski, H.; Wlodarczyk, J.; Jacenik, D.; Kordek, R.; Krajewska, W.M.; Misicka, A.; Fichna, J.; et al. High activity of the endogenous opioid system and acute but not chronic stress influence experimental colitis development in mice. J. Physiol. Pharmacol. 2019, 69, 769–778. [Google Scholar]
- Boligon, A.A.; De Freitas, R.B.; De Brum, T.F.; Waczuk, E.P.; Klimaczewski, C.V.; De Ávila, D.S.; Athayde, M.L.; Bauermann, L.D.F. Antiulcerogenic activity of Scutia buxifolia on gastric ulcers induced by ethanol in rats. Acta Pharm. Sin. B 2014, 4, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Esrefoglu, M.; Akinci, A.; Taslidere, E.; Elbe, H.; Cetin, A.; Ateş, B. Ascorbic acid and beta-carotene reduce stress-induced oxidative organ damage in rats. Biotech. Histochem. 2016, 91, 455–464. [Google Scholar] [CrossRef]
- Koh, S.-J.; Kim, J.W.; Kim, B.G.; Lee, K.L.; Kim, J.S. Restraint stress induces and exacerbates intestinal inflammation in interleukin-10 deficient mice. World J. Gastroenterol. 2015, 21, 8580–8587. [Google Scholar] [CrossRef]
- Siracusa, R.; Fusco, R.; Peritore, A.F.; Cordaro, M.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. The antioxidant and anti-inflammatory properties of anacardium occidentale l. cashew nuts in a mouse model of colitis. Nutrients 2020, 12, 834. [Google Scholar] [CrossRef] [Green Version]
- Petrosino, S.; Campolo, M.; Impellizzeri, D.; Paterniti, I.; Allarà, M.; Gugliandolo, E.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Esposito, E.; et al. 2-Pentadecyl-2-oxazoline, the oxazoline of pea, modulates carrageenan-induced acute inflammation. Front. Pharmacol. 2017, 8, 308. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.R.; Du, L.J.; He, H.Q.; Kim, J.J.; Zhao, Y.; Zhang, Y.W.; Luo, L.; Dai, N. Fructo-oligosaccharide intensifies visceral hypersensitivity and intestinal inflammation in a stress-induced irritable bowel syndrome mouse model. World J. Gastroenterol. 2017, 23, 8321–8333. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Bruschetta, G.; Di Paola, R.; Ahmad, A.; Campolo, M.; Cuzzocrea, S.; Esposito, E.; Navarra, M. The anti-inflammatory and antioxidant effects of bergamot juice extract (BJe) in an experimental model of inflammatory bowel disease. Clin. Nutr. 2015, 34, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; Fusco, R.; D’Amico, R.; Peditto, M.; Oteri, G.; Di Paola, R.; Cuzzocrea, S.; Navarra, M. Treatment with a flavonoid-rich fraction of bergamot juice improved lipopolysaccharide-induced periodontitis in rats. Front. Pharmacol. 2019, 9, 1563. [Google Scholar] [CrossRef] [Green Version]
- Impellizzeri, D.; Peritore, A.F.; Cordaro, M.; Gugliandolo, E.; Siracusa, R.; Crupi, R.; D’Amico, R.; Fusco, R.; Evangelista, M.; Cuzzocrea, S.; et al. The neuroprotective effects of micronized PEA (PEA-m) formulation on diabetic peripheral neuropathy in mice. FASEB J. 2019, 33, 11364–11380. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, R.; Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Scuto, M.; Cuzzocrea, S.; Di Paola, R.; et al. Modulation of NLRP3 inflammasome through formyl peptide receptor 1 (Fpr-1) pathway as a new therapeutic target in bronchiolitis obliterans syndrome. Int. J. Mol. Sci. 2020, 21, 2144. [Google Scholar] [CrossRef] [Green Version]
- Gugliandolo, E.; D’Amico, R.; Cordaro, M.; Fusco, R.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Neuroprotective effect of artesunate in experimental model of traumatic brain injury. Front. Neurol. 2018, 9, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawant, S.; Gokulan, R.; Dongre, H.; Vaidya, M.; Chaukar, D.; Prabhash, K.; Ingle, A.; Joshi, S.; Dange, P.; Joshi, S.; et al. Prognostic role of Oct4, CD44 and c-Myc in radio–chemo-resistant oral cancer patients and their tumourigenic potential in immunodeficient mice. Clin. Oral Investig. 2015, 20, 43–56. [Google Scholar] [CrossRef]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef] [Green Version]
- Campolo, M.; Di Paola, R.; Impellizzeri, D.; Crupi, R.; Morittu, V.M.; Procopio, A.; Perri, E.; Britti, D.; Peli, A.; Esposito, E.; et al. Effects of a polyphenol present in olive oil, oleuropein aglycone, in a murine model of intestinal ischemia/reperfusion injury. J. Leukoc. Biol. 2012, 93, 277–287. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- D’Amico, R.; Fusco, R.; Gugliandolo, E.; Cordaro, M.; Siracusa, R.; Impellizzeri, D.; Peritore, A.F.; Crupi, R.; Cuzzocrea, S.; Di Paola, R. Effects of a new compound containing Palmitoylethanolamide and Baicalein in myocardial ischaemia/reperfusion injury in vivo. Phytomedicine 2019, 54, 27–42. [Google Scholar] [CrossRef]
- Fusco, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Gugliandolo, E.; Siracusa, R.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. N-Palmitoylethanolamide-oxazoline protects against middle cerebral artery occlusion injury in diabetic rats by regulating the SIRT1 pathway. Int. J. Mol. Sci. 2019, 20, 4845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordaro, M.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Peritore, A.F.; D’Amico, R.; Gugliandolo, E.; Di Paola, R.; Cuzzocrea, S. 2-Pentadecyl-2-oxazoline reduces neuroinflammatory environment in the MPTP model of parkinson disease. Mol. Neurobiol. 2018, 55, 9251–9266. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.; Liu, L.; Wei, J.L.; Hu, Z.L.; Li, L.; Wang, S.; Xu, J.M.; Zhou, X.F.; Li, C.Q.; Yang, Z.Y.; et al. Brain-derived neurotrophic factor precursor in the hippocampus regulates both depressive and anxiety-like behaviors in Rats. Front. Psychiatry 2019, 9, 776. [Google Scholar] [CrossRef] [Green Version]
- Siracusa, R.; Paterniti, I.; Cordaro, M.; Crupi, R.; Bruschetta, G.; Campolo, M.; Cuzzocrea, S.; Esposito, E. Neuroprotective effects of temsirolimus in animal models of parkinson’s disease. Mol. Neurobiol. 2017, 55, 2403–2419. [Google Scholar] [CrossRef]
- Zhao, J.; Niu, C.; Wang, J.; Yang, H.; Du, Y.; Wei, L.; Li, C. The depressive-like behaviors of chronic unpredictable mild stress-treated mice ameliorated by Tibetan medicine Zuotai: Involvement in the hypothalamic–pituitary–adrenal (HPA) axis pathway. Neuropsychiatr. Dis. Treat. 2018, 14, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse | Time | LOQ (ng/mL) |
|---|---|---|
| 1 | T0 | <71.4 |
| 30 min | <71.4 | |
| T3 | <71.4 | |
| T6 | <71.4 | |
| T24 | <71.4 | |
| 2 | T0 | <71.4 |
| 30 min | <71.4 | |
| T3 | <71.4 | |
| T6 | <71.4 | |
| T24 | <71.4 | |
| 3 | T0 | <71.4 |
| 30 min | <71.4 | |
| T3 | <71.4 | |
| T6 | <71.4 | |
| T24 | <71.4 | |
| 4 | T0 | <71.4 |
| 30 min | <71.4 | |
| T3 | <71.4 | |
| T6 | <71.4 | |
| T24 | <71.4 | |
| 5 | T0 | <71.4 |
| 30 min | <71.4 | |
| T3 | <71.4 | |
| T6 | <71.4 | |
| T24 | <71.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amico, R.; Siracusa, R.; Fusco, R.; Cordaro, M.; Genovese, T.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Protective Effects of Colomast®, a New Formulation of Adelmidrol and Sodium Hyaluronate, in a Mouse Model of Acute Restraint Stress. Int. J. Mol. Sci. 2020, 21, 8136. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218136
D’Amico R, Siracusa R, Fusco R, Cordaro M, Genovese T, Peritore AF, Gugliandolo E, Crupi R, Impellizzeri D, Cuzzocrea S, et al. Protective Effects of Colomast®, a New Formulation of Adelmidrol and Sodium Hyaluronate, in a Mouse Model of Acute Restraint Stress. International Journal of Molecular Sciences. 2020; 21(21):8136. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218136
Chicago/Turabian StyleD’Amico, Ramona, Rosalba Siracusa, Roberta Fusco, Marika Cordaro, Tiziana Genovese, Alessio Filippo Peritore, Enrico Gugliandolo, Rosalia Crupi, Daniela Impellizzeri, Salvatore Cuzzocrea, and et al. 2020. "Protective Effects of Colomast®, a New Formulation of Adelmidrol and Sodium Hyaluronate, in a Mouse Model of Acute Restraint Stress" International Journal of Molecular Sciences 21, no. 21: 8136. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218136