Review of PIP2 in Cellular Signaling, Functions and Diseases

Institute for Medicine and Engineering, University of Pennsylvania, Philadelphia, PA 19104, USA
Int. J. Mol. Sci. 2020, 21(21), 8342; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218342
Submission received: 21 September 2020
/
Revised: 30 October 2020
/
Accepted: 3 November 2020
/
Published: 6 November 2020
(This article belongs to the Special Issue Phosphoinositides and Downstream Signalling Molecules)
Abstract
:Phosphoinositides play a crucial role in regulating many cellular functions, such as actin dynamics, signaling, intracellular trafficking, membrane dynamics, and cell–matrix adhesion. Central to this process is phosphatidylinositol bisphosphate (PIP2). The levels of PIP2 in the membrane are rapidly altered by the activity of phosphoinositide-directed kinases and phosphatases, and it binds to dozens of different intracellular proteins. Despite the vast literature dedicated to understanding the regulation of PIP2 in cells over past 30 years, much remains to be learned about its cellular functions. In this review, we focus on past and recent exciting results on different molecular mechanisms that regulate cellular functions by binding of specific proteins to PIP2 or by stabilizing phosphoinositide pools in different cellular compartments. Moreover, this review summarizes recent findings that implicate dysregulation of PIP2 in many diseases
1. Introduction
Phosphoinositides (PPIs), are inositol-containing glycerophospholipids bearing variable numbers of phosphate groups on their headgroups [1]. PPIs are multifaceted molecules that have recently become an interesting player in regulating cell function due to their involvement in cellular functions such as actin dynamics, membrane trafficking, regulation of transmembrane proteins and signal transduction [2,3,4,5,6,7,8]. Although the total amount of PPIs in eukaryotic cell membranes is low, they play critical roles in cellular dynamics by regulating multiprotein complexes [9,10]. The spatiotemporal regulation of PPI-mediated biological processes is achieved by interconversion (Figure 1) of the phosphorylation states of PPIs by specific kinases and phosphatases, followed by recruitment of PPI-specific effectors. Inter-conversion of the phosphate groups is spatially controlled by phosphoinositide-metabolizing kinases and phosphatases as required for cellular functions [11,12]. PPIs generate seven possible isoforms by phosphorylating the inositol ring at positions 3, 4, and 5. Three isoforms of PPIs with two phosphate groups connected to the inositol ring, phosphatidylinositol-(4,5)-bisphosphate (PI(4, 5)P2), phosphatidylinositol-(3,5)-bisphosphate (PI(3, 5)P2) and phosphatidylinositol-(3,4)-bisphosphate (PI(3, 4)P2) are the focus of this review. PI(3,4)P2 and PI(3,5)P2 are produced by the phosphorylation of PI3P, and PI(4,5)P2 is produced by the phosphorylation of PI4P or PI5P. The synthesis of PIP2 by phosphorylation of PI5P is regulated by PIP4K, which is one of the less studied pathways. In addition, PIP4K is able to phosphorylate PI3P, but only able to alter PI5P levels in vivo [13] (PI(4, 5)P2) can be further phosphorylated to (PI(3,4,5)P3) and (PI(3,4,5)P3) is converted to (PI(4,5)P2) by the enzyme phosphate and tensin homolog deleted from chromosome 10 (PTEN) [14]. PIP3 stimulates the activity of PDK1 and phosphorylates Akt [15,16]. This PIP3/Akt pathway is intensively studied and regulates many crucial processes in cells. PIP3 and PTEN have been the subjects of excellent recent reviews, and the focus here is on PIP2 [7].
Phosphoinositides control intracellular trafficking, membrane dynamics and cytoskeletal organization by interacting with many different proteins [17,18,19]. PIP2 regulates other membrane phospholipids and their signaling functions [7,17]. The major roles it plays in the cell membrane include cytoskeletal linkage, regulation of ion channels, and intracellular trafficking [20]. PI dynamics and mechanism are precisely controlled by kinase and phosphatase [21,22]. Recent studies showed the direct implication of these enzymes in diseases including liver cancer, glioblastoma or neurodegeneration [7,11,23]. Thus, many studies target phosphoinositide kinase inhibitors for pathological studies.
In this review we summarize the recent development in understanding the role of PIP2 in cellular function and signaling. We first discuss the effect of PIP2 on actin binding proteins, addressing the mechanism of the actin cytoskeletal dynamics such as polymerization or depolymerization of the filamentous network or the coupling to membrane to generate forces. Next, we outline the role of PIP2 in membrane dynamics. We summarized how the membrane organization depends upon PIP2 in the presence of ions or transmembrane proteins that are sensitive to membrane curvature. We discuss how clathrin coated pits interact with adaptor proteins during the endocytosis process, which is facilitated by PIP2. Finally, we discuss the role of PIP2 in cell signaling and diseases.
2. PIP2 in Actin Dynamics
Cytoskeletal dynamics play an important role in many cellular functions such as force generation, intracellular transport, or migration [24,25,26,27]. Actin forms the network inside the cell which is the most responsible for cellular architecture providing the cell a mechanical scaffold [24,27,28]. Accumulated evidence suggests that membrane PIP2 regulates the function of many acting binding proteins including formin, gelsolin, cofilin, profilin, filamin, WASP, ezrin, α- actinin, and others, which control the dynamical organization of the actin network [9,29,30,31,32,33]. PIP2 mostly inactivates actin binding proteins that inhibit actin polymerization and activates proteins which promote filamentous assembly [30,34]. Proteins bind to PIPs via numerus different structures, including the pleckstrin homology (PH) domain of phospholipase C-delta1, the Gag precursor protein Pr55 of HIV-1, phox homology (PX), C2, SH2, protein tyrosine binding, FYVE, PHD, GRAM, BAR, and espin N-terminal homology (ENTH)/ANTH domains, forming a large family of domains collectively [35,36].
Actin polymerization is regulated by a variety of actin binding proteins [28,37]. Actin dynamics depend upon the continuous attachment of G- actin at the barbed (+) end and dissociation at the pointed (−) end, and that defines the filament length [38]. Cofilin is an actin binding protein that binds to both F-actin and G-actin and is a severing protein responsible for actin depolymerization (Figure 2) [38,39,40]. ADF/ cofilin binds to PIP2 through a multivalent mechanism and the dissociation of ADF/cofilin to actin filament can accurately be regulated by changing PIP2 density at the cell membrane [41]. One study found that cofilin binds to PIP2 via a specific pocket which is pH dependent [42]. However, this result contrasts with recent finding showing that cofilin interaction with PIP2 is not pH dependent, but the interaction of profilin with membrane, actin and multiple PIP2 headgroup (clustering) is affected somewhat when pH is increased [41]. Cofilin’s activity depends on phosphorylation, which is regulated by Rho-GTPase and LIM kinase (LIMK) and by binding PPIs [43]. The rho-family small GTPases, Rho, Rac, and Cdc42, play a central role in regulating actin reorganization through their various downstream effectors [44]. LIMK1 and LIMK2 are activated by the GTPase-dependent protein kinases ROCK and PAK1 by phosphorylation of Thr508 and Thr-505, respectively, in the activation loop of the kinase domain [4,45]. LIMK1 and LIMK2 both regulate actin cytoskeletal reorganization by phosphorylating and inactivating cofilin/ADF [4,6]. Hence, cofilin is regulated by the signals from both the Rho and Rac pathways. Epidermal growth factor (EGF) induces sudden loss of PIP2 in membrane that activates local cofilin pool in membrane in carcinoma [46]. These altogether lead to a dramatic turnover of actin monomers (Figure 2).
Gelsolin is another actin severing and capping protein which binds to the barbed end of actin filaments [47]. The barbed end of the filament capped by gelsolin becomes available again through the binding of phosphatidylinositol lipids, such as PIP2, leading to filament elongation. Three PIP2 binding sites for gelsolin have been characterized. Two of the binding sites compete with F-actin and G-actin sites [5,48]. Thus, the severing function of gelsolin can be inhibited by PIP2 [5,47]. Gelsolin can bind to the cell membrane by PIP2 which abrogates the gelsolin interaction with actin. Not only does the amount of phosphoinositide alter the free or actin-bound-gelsolin in cells, but also the lateral distribution of PIP2 controls inactivation of gelsolin [49,50]. Recent studies showed that ATP competes with PIP2 to bind with gelsolin [51,52]. Interaction of gelsolin with PIP2 can be abrogated chemically in vitro by including profilin which is a competing PIP2 binding protein [48]. PIP2 binding to the gelsolin family of capping proteins is enhanced by calcium ions [47]. Ca2+ potentiates gelsolin’s binding to the end of the filament and promotes the polymerization of monomeric actin into filaments [29,48]. Antibacterial activity of rhodamine B (RhB)-conjugated peptides based on the PIP2 binding site of gelsolin, which are cell membrane-permeant, has been shown to kill microorganisms, such as Escherichia, Pseudomonas aeruginosa, and Streptococcus pneumoniae [53,54].
Another important PPI-sensitive player for actin dynamics is the Arp2/3 complex, which regulates nucleation and branching of actin filaments. Lateral organization of PIP2 in the lipid bilayer regulates nucleation [55]. Arp2/3 is primarily activated by the Wiskott-Aldrich syndrome protein (WASP) family, multidomain proteins, and PIP2 promotes this activation (Figure 2). WASP family proteins integrate PIP2 and other signals to regulate cytoskeletal response through the Arp2/3 complex. Moreover, PIP2 interaction with PH domain of WASP regulates the stabilization of WASP at the membrane. In Xenopus egg extracts N-WASP interacts with Cdc42, which is a small GTPase protein of the Rho family and is required for actin polymerization. An increase in N-WASP activity is coordinated by Cdc42 and PIP2 synergistically [34,56,57].
Profilin is another essential actin regulatory protein which interacts with many other proteins [58]. Decreased of profilin1 expression increases cellular motility by regulating PIP2 along with lammellipodin accumulation at the cell leading edge [59]. An in vitro study showed that profilin, isolated from platelets, binds to PIP2 along with other phospholipids in lipid bilayers [60]. Profilin regulates tyrosine kinase-coupled PIP2 hydrolysis [61]. PLC-γ1 hydrolyzes profilin-bound PIP2 by competing the inhibitory effect of profilin [60]. Profilin binds to G-actin and increases the ATP binding to actin [61]. This leads to ATP-actin binding at (+) end of filamentous actin. Profilin binding to actin competes with binding of PIP2. Profilin interacts with actin and poly-L-proline (PLP) stretches, which is essential for profilin function in fission yeast [62]. Profilin binds to both PLP and actin monomers simultaneously [7]. In addition, profilin binds to the cell membrane by PI(4,5)P2, which prevents actin and PLP interaction [7,58].
A large body of literature shows that PIP2 turnover regulates the activity of both gelsolin and profilin. It is clear by now that phosphoinositides and these actin binding proteins interact. However, the mechanism at the molecular level remains elusive. A recent study focuses on different actin binding proteins such as mDia2, N-WASP and gelsolin interaction with PIP2 at the membrane by using molecular dynamics simulation and experimental approaches. The study showed that the cholesterol and PIP2 distribution alters the interaction between actin binding protein and PIP2. With large unilamellar vesicle containing PIP2, a multivalent binding model showed that PIP2 activates mDia2 and NWASP to nucleate straight and branched actin filaments, respectively, but inhibits gelsolin’s ability to cap the fast-growing barbed end of F-actin or to sever the actin filament [49]. Cortactin is also an actin associated protein that can bind and regulate Arp2/3 and N-WASP [63]. Cortactin mutant cells show reduced binding of Arp2/3 complex or dynamin2 to actin. By performing in vitro experiments it is shown that dynamin2 enhances the nucleation of actin by Arp2/3 and cortactin in PIP2 containing vesicles [64].
3. PIP2 in Adhesion Dynamics
PIP2 binds to many focal adhesion proteins, such as vinculin, talin, and the focal adhesion kinase FAK. PIP2 serves as linkage to focal adhesion and actin binding proteins. There are actin binding proteins such as α-actinin, ezrin or filamin which also bind to focal adhesions. A synthetic peptide of α-actinin inhibits PLC-γ1 and PLC-δ1 activity and inhibition is induced by PIP2 competition [40]. PIP2 binding to α-actinin is inhibited by the treatment of cells with platelet derived growth factor, resulting in actin depolymerization. A recent study showed that the architecture of α-actinin-2 and 3 provides a suitable spatial orientation platform for PIP2 bonding by performing molecular dynamics (MD) simulations [65]. In smooth muscle in which α-actinin was discovered, PIP2 is found in large amounts which facilitates gelation of actin [66,67]. The length of smooth muscle depends upon the PIP2 synthesis, which regulates inositol phospholipid turnover [68]. Filamin A is another crosslinker protein which forms contacts between focal adhesions and F-actin. Filamin is associated to the cell membrane by β integrins. PIP2 bound to filamin A inhibits the gel formation of actin. Filamin has three isoforms called FLNa, FLNb, and FLNc. FLNa is recruited by CD28 followed by lipid raft accumulation at the immunological synapsis in T lymphocyte activation. PIP2 is essential for the clustering of lipid raft [69]. Ezrin is one of the ERM (ezrin, radixin, moesin) family proteins, which also forms linkages between the cellular membrane and cytoskeleton. Ezrin exists in both active and inactive states within cells. PIP2 activates Ezrin by binding with it and becomes available for phosphorylation by Rho-kinase and many PKC isoforms [70]. Neutron scattering experiment showed for the first-time, the conformational changes of ezrin when it simultaneously binds to PIP2 and F-actin [71].
Focal adhesion kinase (FAK) is a protein tyrosine kinase implicated in many signaling pathways to regulate cellular functions including migration. When a cell binds to the extracellular matrix (ECM), FAK is recruited to focal adhesion (FA) sites and undergoes conformational change, which is activated by phospholipids such as PIP2 by unblocking the FERM domain and kinase domain. Simulation results show that FAK transiently binds to PIP2 through electrostatic interactions [72]. Molecular dynamics simulation and fluorescence resonance energy transfer (FRET) experiments both showed that FAK binding to ATP decreases the FRET signal confirming that the PIP2 binding acts in the reverse direction [73,74]. Phosphatidylinositol 4-phosphate 5-kinase type Iγ (PIP5KIγ) is required for efficient FAK activation and generates PIP2 locally in FAs by PIP5KIγ. Thus, PIP2 is a strong mediator in integrin-FAK signaling pathways [73].
Talin plays a crucial role in activating integrins [75,76]. Within the cytosol talin is in an inactivated form, where its C-terminal rod domain binds to the N-terminal head domain. Many pathways lead to disruption of the interaction between talin’s C-terminal and N-terminal including binding with PIP5KIγ which generates PIP2 from PI4P [77]. Ye et al. delineate a detailed account of PIP2 in activating talin by using FRET. They showed interaction of talin with lipid bilayers is altered by PIP2 [78]. The FERM domain of talin-1 binds to the cytosolic domain of β3 -integrin weakly (Figure 3). However, the interaction affinity increases three-fold when it synergistically binds to acidic PIP2 [8,79,80]. Membrane bound talin recruits and activates vinculin. Vinculin localizes at the adhesion complex and interacts with PIP2 to associate with the membrane (Figure 3) [81]. Simulation data shows that PIP2 is not required for vinculin localization at FAs but is needed for the activation of FA turnover during mechanotransduction processes [81]. Other studies mentioned that PIP2 is required for FA formation and vinculin phosphorylation and trafficking [82].
4. PIP2 in Membrane Dynamics and Organization
Phosphoinositides are minor component of the lipid bilayer that forms the plasma membrane, constituting 1% of total cellular phospholipid. Eukaryotic cell plasma membranes maintain a balanced composition of sterols, phospho- and sphingo- lipids that is distinct from other cellular membranes, which is required for cellular integrity. All seven PIPs are spatially localized uniquely in the plasma membrane. However, PIP2 is the most abundant among all seven species of PIPs. Many PIP2 binding proteins are characterized as high affinity ligands for these lipids to regulate signaling [83] and are activated by agonists for numerous cell surface receptors [33]. Several studies reported that PIP2 is highly enriched in the plasma membrane within segregated domains with an approximate size of 73 nm by showing PC12 cell staining with anti-PIP2 antibody and high-resolution STED imaging [84,85]. The plasma membrane is fluid like with proteins and lipids co-existing within in a heterogeneous distribution. Also, the negative charge on PIP2 plays a crucial role in the interaction with membrane bound proteins.
4.1. Charge Dependence and Electrostatic Interaction
Over 30 years the electrostatic properties of membranes have been highlighted in the literature [86,87]. Many theoretical models have been proposed based on the smeared charge model of Gouy-Chapman theory, Finite-difference Poisson-Boltzmann (FDPB) method, based on dielectric properties of the solvent [88,89]. Afterwards, it was proposed that flat lipid bilayers can be considered for the electrostatic calculations of the present PIP-based systems when proper choice of orientations are made, concluding that the specific charge of PIP2 with respect to the cell membrane is required for lipid signaling events to occur [90]. Effort has been made to understand the atomic level structural of PIP2 such as its protonation state and binding to cations, by using hybrid quantum mechanics and molecular mechanics simulation methods which determine the optimal geometry of PIP2 [91]. PIP2 has high negative charge density obtained by deprotonation of two phosphomonoester groups, which can range from −3e to −5e, depending on pH and the counterions present, which brings the net lipid charge to -3.99 ± 0.10e. The charges on PIP2 regulate its interaction with proteins [55].
Another important characteristic of, phosphatidylinositol-(4,5)-bisphosphate (PI(4, 5)P2) is that the different lateral organizations, such as small clusters or large stable aggregates, which are interconvertible, within the region of the membrane are associated with diverse functionality [92,93]. PI(4, 5)P2 turnover at the plasma membrane has been observed by immunofluorescence probes suggesting the evidence of spatially segregated of PI(4, 5)P2 pools [93]. Non-homogeneous distribution of PI(4, 5)P2 in membranes is due to electrostatic interaction between neighboring lipids [85,94]. Levental et al. showed that of PIP2 clustering depends upon the multivalency of the counterion and high charge density of the lipids by using lipid monolayers [93]. Lateral organization on a large range of length scales can be remodeled when Ca2+ is introduced to PIP2 containing membrane monolayers at different concentrations. This leads to domain formation and reduces phase co-existence surface pressure in of PIP2 containing monolayer [93]. The formation of the domains or nano clusters has relevance in cellular function, and is regulated by the Ca2+ ions in the absence of proteins [94,95]. Not only Ca2+ but other divalent ions such as Mg2+ and Zn2+ also affect lateral organization of PIP2 in the asymmetric membrane at physiological concentration which in turn regulate PIP2 protein interaction [94]. Bradley et al. have characterized multivalent lipid cation interaction by the number of lipids bound within a specific distance (called N-bridge), showing the largest cluster formations up to 60 lipids for the combination of PIP2 and Ca2+ (Figure 4). The formation of clusters is also dependent on physiological trivalent ions such as Fe3+ and Al3+ [96]. These findings suggest that the electrostatic sequestration and condensation of PIP2 by divalent and trivalent ions resulted in increasing the molecular packing and ordering the more disordered phase, which has important biological relevance.
In the lipid bilayer, lateral distribution of PIP2 has been affected by both electrostatic interaction and cholesterol dependent phase mixing [50]. The cholesterol enriched region in the membrane forms heterogeneous nanoscale clusters having a size of 10–200 nm, known as lipid rafts which are compartmentalized in the plasma membrane and regulate different cellular functions [84]. Nano clusters of phosphoinositide, localized in the membrane can be visualized by fluorescently labeled pleckstrin homology (PH) domains, which allow PIP2 visualization by protein-domain–GFP chimeras in live cells and PLCδ1-PH at plasma membrane and OSBP-PH at Golgi membrane; and for PI(3,4)P2 visualization with Akt-PH at plasma membrane [36,97].
4.2. PIP2 Regulation in Membrane Curvature Sensing and Transport
PIP2 interacts with many transmembrane proteins such as Bin-Amphiphysin-Rvs (BAR) domain proteins, curvature sensing proteins that are important in regulating membrane shape transitions during endocytosis and membrane trafficking [98,99]. These BAR domain protein interactions with PIP2 are charge dependent. By coarse grain modeling Li et al. showed that the electrostatic interaction between PIPs head group which contains large negative charges and many positive charged residues in the BAR is the origin of membrane binding [100]. PIP2 binds to both sides of BAR proteins to form membrane protrusion by synergistically binding to actinm [101]. Experimentally and by simulation it has been shown that PIP2 has preference in binding to the negatively curved membrane over positively curved membranes (Figure 5) [98]. Thus, membrane curvature can promote the spatial regulation on PIP2 binding and local enrichment of lipids. It has been shown in vitro and also in cells that the phosphoinositide binding domain of BIN1 targets the membrane by interacting with PIP2.The N-BAR domain of BIN1 clusters with PIP2 to promote the recruitment of its downstream partner dynamin and is responsible for membrane tubulation2. Amphiphysin1 (BIN1) in PIP2-containing membranes induces curvature [102]. Membrane curvature sensing and generation of BIN1 is abrogated in membranes lacking PIP2. However, BIN1 alone can initiate membrane tubulation (Figure 6). BIN1 membrane curvature sensing and generation show autoinhibition regulated by downstream ligands and PIP2 [102]. A recent study demonstrates that mutation of BIN1 N-BAR impairs membrane T tubulation [103]. This affects the regulation of muscle functioning or nuclear positioning, leading to diseases like centronuclear myopathies [2,104].
PIP2 is a major regulator of voltage gated ion channels, in which PIP2 binds to the transmembrane domain [20]. Kobayashi et al. showed in skeletal muscle that PIP2 is a major activator of Ca2+ channels. Depletion of PIP2 induces increases in voltage sensitivity and a decrease in voltage amplitude in K+ ion channels in Xenopus oocytes. PIP2 controls both movement and stability of the channels by interacting through linkers [21]. ATP-sensitive K+ channel rundown, the process by which a channel steadily decreases in conductance until the channel inactivates, is induced by Ca2+, and this process is shown to be regulated by PIP2. KCNQ is another family of channels that absolutely requires PIP2. The importance of PIP2 in modulating KNCQ channels is well studied in neurons, showing that PIP2 hydrolysis increases neuroexcitability and in cardiac arrythmias in patients by showing PIP2 dependent channel activation [22,105].
A potentially important event that occurs at the cell surface is the interaction between the lipid bilayer with Ras, a small GTPase and with its effectors. These interactions are shown by molecular dynamics simulation or FRET in live cells. Ras proteins, such as H-Ras, N- Ras, and K-Ras, operate in the inner plasma membrane and are mutated in many cancer types [106]. Recent studies have shown Ras enrichment in nano clusters within phosphatidylserine-rich regions. PIP2 binds to Ras G-domain and K-Ras4b HVR, which is one of the isoforms of Ras3. Experimental or computational studies showed the tight binding between PI(4, 5)P2 and K-Ras4b by measuring rotational dynamics by random amine labeling and by atomic force microscopy. The rotational dynamics of K-Ras are important for signaling in cancer cells [3,107].
5. Intracellular Trafficking
PPIs are spatially localized in different compartments in intracellular organelles [7,108]. For example, the Golgi is enriched with PI(3)P or PI(4)P, which are also enriched in early endosomes [109,110]. The Golgi plays a crucial role in membrane trafficking [18,25]. Phosphoinositide 3- kinase converts PI(4, 5)P2 to PI(3,4,5)P3 which is important for vesicular trafficking [111]. New studies demonstrate that the cell surface membrane is a major site of action for PIP2, and the localization of it in different compartments is directly correlated to intracellular trafficking such endocytosis and exocytosis [112]. By specifically interacting with proteins, PIP2 controls the formation and spatiotemporal organization of many protein complexes that are involved in intracellular trafficking.
Clathrin mediated endocytosis, in which cargo is packaged into vesicles with clathrin coating, plays a crucial role in cell signaling, migration and cell-cell interactions. PIP2 has been implicated in clathrin-mediated endocytosis [113]. However, clathrin does not directly bind to the membrane or cargos but to adaptor proteins such as adaptor protein 2 (AP2) or accessory protein AP180 and espin [114]. A recent study reported that during clathrin coated pit (CCP) assembly initiation AP2 is recruited to the plasma membrane and colocalizes with the nucleation complex which binds to both cargo and PIP2, when stained with anti- PIP2 antibody111. The formation of PI(3,4)P2 by class II PI3-kinase C2α (PI(3)K C2α) spatiotemporally controls clathrin-mediated endocytosis. The depletion of PI(3,4)P2 hinders the maturation of CCPs before fission. PIP5K is associated with the initiation of CCPs but its activity is not found to mature them. Another study shows that PIP2 is an established regulator of endocytosis. Endosomal PI(4,5)P2 is required for the sorting of active epidermal growth factor receptor (EGFR) towards multivesicular bodies (MVB) and further termination of the signal (Figure 7). Sun et al. showed that type I gamma phosphatidylinositol phosphate kinase (PIPKIγ) is an enzyme that synthesizes PIP2 by phosphorylation of PtdIns4P and regulates EGFR sorting from endosomes to lysosomes [115]. This was done by performing flow cytometry analysis and quantification of internalized Alexa Fluor 488-labelled EGF in control and PIPIγi5- knockdown cells [111,115]. PIPIγi5 interacts with sorting nexin 5 (ANX5) which is the effector of PIP2 in the early endosome, but cells lacking SNX5 still localize PIPIγi5 to endosomes [116]. SNX5 has been reported to inhibit EGFR degradation when overexpressed. However, knockdown of SNX5 does not affect EGFR trafficking to early endosomes, but blocks trafficking to the late endosome/lysosome [115,117]. On the other hand, EGFR regulates Ras activity, which is implicated in PIP3 and MAP kinase pathways [118].
Another crucial role of PIP2 is in bidirectional homeoprotein trafficking. Homeoproteins are a class of transcription factor that predominantly resides in the nucleus. Chick engrailed 2 (EN2) is a homeoprotein that shuttles between the nucleus and cytosol. In the cytosol, EN2 associates with those membrane fractions enriched in cholesterol and glycosphingolipids. EN2 directly binds to PIP2. Dephosphorylation of PIP2 reduces EN2 secretion. Moreover, PIP2 is involved in EN2 internalization [119].
Phosphoinositides are interconvertible and the balance of production and usage is tightly controlled in a specific organelle. Contrary to the plasma membrane, the Golgi membrane has less PIP2 and high abundance of PI4P and PI4K enzymes [1,120,121,122]. There is a possibility that the plasma membrane PI4P pool is due to vesicular trafficking of PI4P from Golgi membrane. The recent discovery of lipid binding domains enables the lifetime monitoring of lipid synthesis by fusing with green fluorescent protein GFP. GFP-tagged PKB/Akt PH and GFP-PH (PLCδ) are possible markers for live monitoring PIP3 and PIP2. Utilizing recent advancements, a study demonstrated PIP2 level decreases in the plasma membrane when stimulated by angiotensin II (AngII) by showing the change in PLCδ1PH-GFP expression level in HEK-293 cells and an increase of GFP in cytosol [123,124]. Recovery experiment shows that Golgi PI4P eliminated cells recover slowly compare to control. These studies confirm that although PI4P takes part in the maintenance of the PIP2 level pool at plasma membrane, it is not requisite for the process [109].
Phosphatidylinositol 3,5-bisphosphate PI(3,5)P2 is synthesized from PI3P by FYVE-domain-containing PI kinase (PIKfyve) in mammalian cells [125]. FYVE domain appears to target the enzyme to PI3P -rich membranes [126]. However, a similar process occurs in yeast called Saccharomyces cerevisiae, and the PI(3,5)P2 synthesis is found to be processed by Fab1p. Since Fab1 is not responsible for the full synthesis, additional unknown effector proteins are expected to be involved. PI(3,5)P2 is involved in vacuole to lysosome membrane trafficking and packaging of proteins in multivesicular bodies (MVBs) [1,127].
6. PIP2 in Signaling and Diseases
Accumulating evidence suggests that PIP2 dysregulation contributes to cancer including melanoma, breast cancer, leukemia, prostate cancer (Table 1) [128,129]. Literature suggests that PIP2 is implicated in many pathways and binds to signaling proteins such as lamellipodin/RAPH1, tandem PH domain-containing proteins TPP1 and TAPP2 and PIP3 binding proteins including protein kinase Akt/PKB [59,130]. The PIP3-Akt signaling pathway is implicated in many diseases [130,131].
Phosphoinositides play a major role in intracellular signaling pathways which are implicated in carcinogenesis such as hepatocellular carcinoma (HCC) or melanoma [33,128,139,140]. Thus, many signaling pathways are targeted for therapies including phosphoinositide 3-kinase (PI3K)/Akt, mitogen-activated protein kinase (MAPK) pathways [141]. Numerous proteins are regulated downstream of these pathways. Generally, pathways are activated by the alteration of the cell’s microenvironment or genetic alteration [33]. Our recent work shows that, when Huh7 cells, a hepatocellular carcinoma cell line, adhere to soft hyaluronic acid (HA) gels, they show similar behavior as cells adhered on stiff polyacrylamide gels by regulating phosphoinositide signaling. The result is confirmed by pAkt expression level by immunoblotting and by quantifying the total amount of PIP3 on HA and Poly acrylic acid (PAA) substrates by using mCherry Grp1, a fluorescent protein that specifically binds to PIP3 [33,142]. PIP3 which increases Akt activity via PDK1, is activated by Ras (Figure 8). PDK1 activates Akt by phosphorylation of threonine site. The overexpression of PAK, which is one of the downstream effectors of PIP3, is correlated with many cancer types such as ovarian cancer [15,106]. PI3K catalyzes PIP2 into PIP3 [143] (Figure 8). One of the major downstream effectors of Akt is mTORc1 which is deregulated in many cancers when phosphatase and tensin homolog deleted on chromosome 10 (PTEN) gene dephosphorylates PIP3 to PIP2 (Figure 8) [15].
The PIP3-Akt pathway also synergistically activates MAPK signaling pathway in melanoma cancer development [118]. The MAPK pathway is one of the most investigated signaling pathways in melanoma cancer [143]. Thus, a series of inhibitors for these pathways are targeted for the therapeutics of melanoma. It has been observed that PI3K activity is increased in melanoma due to loss of PTEN (Figure 8) or increased levels of Akt3 activity, and that plays a crucial role in early melanoma development. A recent study has shown that Akt3 phosphorylates B-Raf (Figure 8) which is often mutated in melanoma cancer [118]. Lipid binding domains such as PH domains of Akt/PKB, are important in signaling, which depends upon PIP synthesis. Akt/PKB binds to PI(3,4)P2 and PIP3 to regulate cell survival and growth, which is independent of PI(4,5)P2. In cases of melanoma, 50% of patients progress to metastatic stress due to upregulation of protein tyrosine phosphate (PTP) promoting cell migration and invasiveness [139]. PTPs bear phosphatase activity toward lipidic substrates, including phosphoinositides. PRL-3 is one of the dual specificity phosphatases which is associated with intracellular membranes and cellular migration [129]. PRL-3 dephosphorylates PI(4.5)P2 and thus alters the phosphoinositide level in cells [144]. PRLs are overexpressed in many cancer types and have become the target of many cancer therapies including melanoma [129,139,145].
ORCL (oculo-cerebro-renal syndrome of Lowe) enzymatic activity is found in many compartments in cells especially concentrated at the Golgi network. The mutation of ORCL causes oculo-cerebro-renal syndrome of Lowe which is an X-linked condition [133]. Lowe syndrome leads to many diseases including renal Fancomi syndrome, glaucoma, cataracts, blindness, mental retardation (Table 1). ORCL is a key component of endocytic trafficking which is involved in clathrin coated pits and other binding motifs, such as AP2, APPL1, and Rab GTPase, including Rab5, Rab6, and Rab14. Therefore, inactivation of ORCL leading to deregulation of PI(4,5)P2 level influences trans- Golgi network and endosomal activity. Imbalance of PIP2 levels further affects actin dynamics and actin binding proteins. Moreover, ORCL controls reabsorption of proteins via PI(4,5)—phosphatase in renal proximal tubule cells (PTCs) [134]. Another regulator for Down’s syndrome is synaptojanin1 which acts on both PI(4,5)P2 and PI(3,5)P2, found in endocytic intermediate nerve terminals. Synaptojanin regulates the actin pool, as well as the de-coating of cathrin mediated endocytic vesicles and synaptic vesicles. In synaptojanin deficient mice, the PI(4,5)P2 level increases whereas a decreased level of cytosolic inositol 5 phosphatases in neurons is observed. Also, an increase in clathrin coated vesicles in nerve terminals is observed [135]. Inositol polyphosphate-4-phosphatase (INPP4) which binds to PI(4,5)P2, shows a reduced level in an asthma mouse model due to restrictive stress [130]. Oxidative stress which is generated by reactive oxygen species (ROS) stimulates the accumulation of PI(4,5)P2. ROS has been implicated in airway inflammation. INPP4 deficiency also leads to cancer, including breast cancer and neurodegeneration [131].
The mutation of myotubularin (MTM) causes several disorders such as failure of skeletal muscle development. MTM related proteins, MTMR1-13 which is an inactive partner of MTMR2 causes the same mutation as active member. Each MTM protein regulates a specific pool of PI(3)P and PI(3,5)P2. Another disease where PI(4,5)P2 regulation is important involves the human immunodeficiency virus-1 (HIV-1). Viral entry into the host cell requires actin cytoskeletal reorganization. Viral receptor clustering is regulated by actin adaptor proteins, such as moesin, filamin A, gelsolin, tailn, vinculin, profilin, WASP, and Arp2/3, that are controlled by PI(4,5)P2. PI(4,5)P2 production is regulated by HIV-1 attachment and promotes viral infection. Hence, the virus controls actin dynamics during cycle, by facilitating actin polymerization and depolymerization [137]. In HIV-1 infection, CD4 and coreceptors clustering at the cell surface is induced by glycoprotein g120, which facilitates virus envelope and cell membrane fusion. PI(4,5)P2 is required to recruit the gag protein at the cell membrane to facilitate invasion. A high density of PI(4,5)P2 is not only required for HIV-1 recruitment but also to maintain glycoprotein at the membrane [136]. PIP2 plays a central role in many neuronal and synaptic functions by regulating endocytosis, exocytosis, cytoskeletal reorganization, and ion channels. In Alzheimer’s disease, Amyloid- β peptide(Aβ) oligomers cause impairment of synaptic function. Elevation of Aβ in the Alzheimer’s diseased brain results in decreased levels of PI(4,5)P2 [138].
7. Conclusions
This review summarizes the role of PIP2 and other PPIs in cell membrane dynamics, focal adhesion, actin organization, intracellular signaling and disease. PIP2 regulates actin binding protein activity which either promotes polymerization and depolymerization of actin filament. Past evidence suggests that actin is connected to the membrane via actin binding proteins such as α-actinin or filamin which are regulated by phosphoinositides. These interactions also affect the binding of actin filaments with focal adhesion proteins such as paxillin, talin, FAK, or vinculin. The distribution of PIP2 in the membrane regulates cell signaling. PIP2 activity depends upon the concentration of cholesterol and divalent ions such as Ca2+, Mg2+, or Zn2+. In addition, PIP2 plays a crucial role in modulating many signaling pathways such as PIP3/Akt, mTORc1, or Rho dependent pathways that have implications in many diseases including cancer, neurodegenerative disease, or down syndrome.
Although PPIs are essential for many cellular functions, there are disparities in many processes which need further studies. PIP2 plays an important role in actin reorganization and filament dynamics. However, the role of PIP2 in any other cytoskeletal component has not yet been well studied. Among PIP2 binding actin proteins, LIMK1 and LIMK2 play an overlapping role in actin reorganization in the Rho-ROCK pathway. Further studies are required to differentiate the functional role of LIMK1 and LIMK2. Moreover, it is unclear if members of ROCK and PAK family proteins function as LIMK- activating kinases. Cortactin shows dependencies on PIP2 and Rac in dissociating from actin–myosin complex, although the direct implication of PIP2 in regulating cortactin still remains controversial [146], and other activators such as the endocytic protein Abp1p remain unclear. It has been shown that a synthetic peptide of α-actinin inhibits PLC-γ1 and PLC-δ1. It is ambiguous whether PIP2 bound to α-actinin is hydrolyzed by activated PLC-γ1 or not. The interaction of vinculin and membrane is based upon either full length or tail domain of vinculin in lipid bilayers or in cells. However, a specific lipid binding site has yet to be discovered.
Funding
This research was funded by the Center for Engineering MechanoBiology (CEMB), an NSF Science and Technology Center, under grant agreement CMMI:15-48571 and Physical Science Oncology Center (PSOC) grant U54-CA193417.
Acknowledgments
I thank Paul A. Janmey for his valuable feedback on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Downes, C.P.; Gray, A.; Lucocq, J.M. Probing phosphoinositide functions in signaling and membrane trafficking. Trends Cell Biol. 2005, 15, 259–268. [Google Scholar]
- Picas, L.; Viaud, J.; Schauer, K.; Vanni, S.; Hnia, K.; Fraisier, V.; Roux, A.; Bassereau, P.; Gaits-Iacovoni, F.; Payrastre, B.; et al. BIN1/M-Amphiphysin2 induces clustering of phosphoinositides to recruit its downstream partner dynamin. Nat. Commun. 2014, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- McLean, M.A.; Stephen, A.G.; Sligar, S.G. PIP2 Influences the Conformational Dynamics of Membrane-Bound KRAS4b. Biochemistry 2019, 58, 3537–3545. [Google Scholar] [CrossRef] [PubMed]
- Romarowski, A.; Battistone, M.A.; La Spina, F.A.; del Puga Molina, L.C.; Luque, G.M.; Vitale, A.M.; Cuasnicu, P.S.; Visconti, P.E.; Krapf, D.; Buffone, M.G. PKA-dependent phosphorylation of LIMK1 and Cofilin is essential for mouse sperm acrosomal exocytosis. Dev. Biol. 2015, 405, 237–249. [Google Scholar] [CrossRef] [Green Version]
- Liepiņa, I.; Czaplewski, C.; Janmey, P.; Liwo, A. Molecular dynamics study of a gelsolin-derived peptide binding to a lipid bilayer containing phosphatidylinositol 4,5-bisphosphate. Biopolymers 2003, 71, 49–70. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Tanabe, K.; Eto, T.; Narumiya, S.; Mizuno, K. LIM-kinase 2 induces formation of stress fibres, focal adhesions and membrane blebs, dependent on its activation by Rho-associated kinase-catalysed phosphorylation at threonine-505. Biochem. J. 2001, 354, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Walter, L.M.; Franz, P.; Lindner, R.; Tsiavaliaris, G.; Hensel, N.; Claus, P. Profilin2a-phosphorylation as a regulatory mechanism for actin dynamics. FASEB J. 2020, 34, 2147–2160. [Google Scholar] [CrossRef] [Green Version]
- Saltel, F.; Mortier, E.; Hytönen, V.P.; Jacquier, M.C.; Zimmermann, P.; Vogel, V.; Liu, W.; Wehrle-Haller, B. New PI(4,5)P2- and membrane proximal integrin-binding motifs in the talin head control β3-integrin clustering. J. Cell Biol. 2009, 187, 715–731. [Google Scholar] [CrossRef] [Green Version]
- Janmey, P.A.; Bucki, R.; Radhakrishnan, R. Regulation of actin assembly by PI(4,5)P2 and other inositol phospholipids: An update on possible mechanisms. Biochem. Biophys. Res. Commun. 2018, 506, 307–314. [Google Scholar]
- Reversi, A.; Loeser, E.; Subramanian, D.; Schultz, C.; De Renzis, S. Plasma membrane phosphoinositide balance regulates cell shape during Drosophila embryo morphogenesis. J. Cell Biol. 2014, 205, 395–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viaud, J.; Mansour, R.; Antkowiak, A.; Mujalli, A.; Valet, C.; Chicanne, G.; Xuereb, J.M.; Terrisse, A.D.; Séverin, S.; Gratacap, M.P.; et al. Phosphoinositides: Important lipids in the coordination of cell dynamics. Biochimie 2016, 125, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Goud, B.; Picas, L.; Gaits-Iacovoni, F. The emerging role of phosphoinositide clustering in intracellular trafficking and signal transduction. F1000Research 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Rameh, L.E.; Tolias, K.F.; Duckworth, B.C.; Cantley, L.C. A new pathway for synthesis of phosphatidylinositol-4,5-bisphosphate. Nature 1997, 390, 192–196. [Google Scholar] [CrossRef]
- Liu, C.; Deb, S.; Ferreira, V.S.; Xu, E.; Baumgart, T. Kinetics of PTEN-mediated PI(3,4,5)P3 hydrolysis on solid supported membranes. PLoS ONE 2018, 13, e0192667. [Google Scholar] [CrossRef] [Green Version]
- Thillai, K.; Lam, H.; Sarker, D.; Wells, C.M. Deciphering the link between PI3K and PAK: An opportunity to target key pathways in pancreatic cancer? Oncotarget 2017, 8, 14173–14191. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, K.M.; Barbie, D.A.; Davies, M.A.; Rabinovsky, R.; McNear, C.J.; Kim, J.J.; Hennessy, B.T.; Tseng, H.; Pochanard, P.; Kim, S.Y.; et al. AKT-independent signaling downstream of oncogenic PIK3CA mutations in human cancer. Cancer Cell 2009, 16, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Balla, T. Phosphoinositides: Tiny lipids with giant impact on cell regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef]
- Goud, B.; Liu, S.; Storrie, B. Rab proteins as major determinants of the Golgi complex structure. Small GTPases 2018, 9, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, M.; Tian, A.; Esposito, C.; Baumgart, T. Dynamic sorting of lipids and proteins in membrane tubes with a moving phase boundary. Proc. Natl. Acad. Sci. USA 2010, 107, 7208–7213. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.L. Complex roles of PIP2 in the regulation of ion channels and transporters. Am. J. Physiol. Ren. Physiol. 2007, 293, 1761–1765. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Menchaca, A.A.; Adney, S.K.; Tang, Q.-Y.; Meng, X.-Y.; Rosenhouse-Dantsker, A.; Cui, M.; Logothetis, D.E.; Designed, D.E.L.; Performed, M.C. PIP 2 controls voltage-sensor movement and pore opening of Kv channels through the S4-S5 linker. Proc. Natl. Acad. Sci. USA 2012, 109, E2399–E2408. [Google Scholar] [CrossRef] [Green Version]
- Zaydman, M.A.; Silva, J.R.; Delaloye, K.; Li, Y.; Liang, H.; Larsson, H.P.; Shi, J.; Cui, J. Kv7.1 ion channels require a lipid to couple voltage sensing to pore opening. Proc. Natl. Acad. Sci. USA 2013, 110, 13180–13185. [Google Scholar] [CrossRef] [Green Version]
- Erle, D.J.; Sheppard, D. The cell biology of asthma. J. Cell Biol. 2014, 205, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Mandal, K.; Wang, I.; Vitiello, E.; Orellana, L.A.C.; Balland, M. Cell dipole behaviour revealed by ECM sub-cellular geometry. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Guet, D.; Mandal, K.; Pinot, M.; Hoffmann, J.; Abidine, Y.; Sigaut, W.; Bardin, S.; Schauer, K.; Goud, B.; Manneville, J.B. Mechanical role of actin dynamics in the rheology of the Golgi complex and in Golgi-associated trafficking events. Curr. Biol. 2014, 24, 1700–1711. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, D.; Tardin, C.; Schmidt, C.F.; Mackintosh, F.C. Nonequilibrium mechanics of active cytoskeletal networks. Science 2007, 315, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Mandal, K.; Balland, M.; Bureau, L. Thermoresponsive micropatterned substrates for single cell studies. PLoS ONE 2012, 7, e37548. [Google Scholar] [CrossRef] [Green Version]
- Mandal, K.; Asnacios, A.; Goud, B.; Manneville, J.-B. Mapping intracellular mechanics on micropatterned substrates. Proc. Natl. Acad. Sci. USA 2016, 113, E7159–E7168. [Google Scholar] [CrossRef] [Green Version]
- Honigmann, A.; Van Den Bogaart, G.; Iraheta, E.; Risselada, H.J.; Milovanovic, D.; Mueller, V.; Müllar, S.; Diederichsen, U.; Fasshauer, D.; Grubmüller, H.; et al. Phosphatidylinositol 4,5-bisphosphate clusters act as molecular beacons for vesicle recruitment. Nat. Struct. Mol. Biol. 2013, 20, 679–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.L.; Janmey, P.A. Phosphoinositide Regulation of the Actin Cytoskeleton. Annu. Rev. Physiol. 2003, 65, 761–789. [Google Scholar] [CrossRef] [PubMed]
- Pogoda, K.; Bucki, R.; Byfield, F.J.; Cruz, K.; Lee, T.; Marcinkiewicz, C.; Janmey, P.A. Soft Substrates Containing Hyaluronan Mimic the Effects of Increased Stiffness on Morphology, Motility, and Proliferation of Glioma Cells. Biomacromolecules 2017, 18, 3040–3051. [Google Scholar] [CrossRef] [PubMed]
- Gowrishankar, K.; Ghosh, S.; Saha, S.; Mayor, S.; Rao, M. Active Remodeling of Cortical Actin Regulates Spatiotemporal Organization of Cell Surface Molecules. Cell 2012, 149, 1353–1367. [Google Scholar] [CrossRef] [Green Version]
- Mandal, K.; Raz-Ben Aroush, D.; Graber, Z.T.; Wu, B.; Park, C.Y.; Fredberg, J.J.; Guo, W.; Baumgart, T.; Janmey, P.A. Soft Hyaluronic Gels Promote Cell Spreading, Stress Fibers, Focal Adhesion, and Membrane Tension by Phosphoinositide Signaling, Not Traction Force. ACS Nano 2018, acsnano.8b05286. [Google Scholar] [CrossRef]
- Yamamoto, M.; Hilgemann, D.H.; Feng, S.; Bito, H.; Ishihara, H.; Shibasaki, Y.; Yin, H.L. Phosphatidylinositol 4,5-bisphosphate induces actin stress-fiber formation and inhibits membrane ruffling in CV1 cells. J. Cell Biol. 2001, 152, 867–876. [Google Scholar] [CrossRef] [Green Version]
- Lemmon, M.A. Membrane recognition by phospholipid-binding domains. Nat. Rev. Mol. Cell Biol. 2008, 9, 99–111. [Google Scholar] [CrossRef]
- Levine, T.P.; Munro, S. The pleckstrin homology domain of oxysterol-binding protein recognises a determinant specific to Golgi membranes. Curr. Biol. 1998, 8, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Senju, Y.; Kalimeri, M.; Koskela, E.V.; Somerharju, P.; Zhao, H.; Vattulainen, I.; Lappalainen, P. Mechanistic principles underlying regulation of the actin cytoskeleton by phosphoinositides. Proc. Natl. Acad. Sci. USA 2017, 114, 8977–8986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugyi, B.; Carlier, M.-F. Control of Actin Filament Treadmilling in Cell Motility. Ann. Rev. Biophys. 2010, 39, 449–470. [Google Scholar] [CrossRef]
- Bernstein, B.W.; Bamburg, J.R. ADF/Cofilin: A functional node in cell biology. Trends Cell Biol. 2010, 20, 187–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakisaka, T.; Itoh, T.; Miura, K.; Takenawa, T. Phosphatidylinositol 4,5-bisphosphate phosphatase regulates the rearrangement of actin filaments. Mol. Cell. Biol. 1997, 17, 3841–3849. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Hakala, M.; Lappalainen, P. ADF/cofilin binds phosphoinositides in a multivalent manner to act as a PIP(2)-density sensor. Biophys. J. 2010, 98, 2327–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyman, S.; Sidani, M.; Ritsma, L.; Waterschoot, D.; Eddy, R.; Dewitte, D.; Debeir, O.; Decaestecker, C.; Vandekerckhove, J.; Van Rheenen, J.; et al. Unbalancing the phosphatidylinositol-4,5-bisphosphate-cofilin interaction impairs cell steering. Mol. Biol. Cell 2009, 20, 4509–4523. [Google Scholar] [CrossRef] [Green Version]
- Sumi, T.; Matsumoto, K.; Takai, Y.; Nakamura, T. Cofilin phosphorylation and actin cytoskeletal dynamics regulated by Rho- and Cdc42-activated LIM-kinase 2. J. Cell Biol. 1999, 1519–1532. [Google Scholar] [CrossRef] [Green Version]
- Croisé, P.; Eestay-Aahumada, C.; Gasman, S.; Ory, S. Rho GTPases, phosphoinositides, and actin: A tripartite framework for efficient vesicular trafficking. Small GTPases 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Prunier, C.; Prudent, R.; Kapur, R.; Sadoul, K.; Lafanechère, L. LIM kinases: Cofilin and beyond. Oncotarget 2017, 8, 41749–41763. [Google Scholar] [CrossRef] [Green Version]
- Van Rheenen, J.; Song, X.; Van Roosmalen, W.; Cammer, M.; Chen, X.; DesMarais, V.; Yip, S.C.; Backer, J.M.; Eddy, R.J.; Condeelis, J.S. EGF-induced PIP2 hydrolysis releases and activates cofilin locally in carcinoma cells. J. Cell Biol. 2007, 179, 1247–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuominen, E.K.J.; Holopainen, J.M.; Chen, J.; Prestwich, G.D.; Bachiller, P.R.; Kinnunen, P.K.J.; Janmey, P.A. Fluorescent phosphoinositide derivatives reveal specific binding of gelsolin and other actin regulatory proteins to mixed lipid bilayers. Eur. J. Biochem. 1999, 263, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Janmey, P.A.; Stossel, T.P. Modulation of gelsolin function by phosphatidylinositol 4,5-Bisphosphate. Nature 1987, 325, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Fatunmbi, O.; Bradley, R.P.; Kandy, S.K.; Bucki, R.; Janmey, P.A.; Radhakrishnan, R. A multiscale biophysical model for the recruitment of actin nucleating proteins at the membrane interface. Soft Matter 2020, 16, 4941–4954. [Google Scholar] [CrossRef]
- Wang, Y.H.; Bucki, R.; Janmey, P.A. Cholesterol-Dependent Phase-Demixing in Lipid Bilayers as a Switch for the Activity of the Phosphoinositide-Binding Cytoskeletal Protein Gelsolin. Biochemistry 2016, 55, 3361–3369. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Cao, L.; Wang, C.; Gigant, B.; Knossow, M. Kinesin, 30 years later: Recent insights from structural studies. Protein Sci. 2015, 24, 1047–1056. [Google Scholar] [CrossRef] [Green Version]
- Szatmári, D.; Xue, B.; Kannan, B.; Burtnick, L.D.; Bugyi, B.; Nyitrai, M.; Robinson, R.C. ATP competes with PIP2 for binding to gelsolin. PLoS ONE 2018, 13, e0201826. [Google Scholar] [CrossRef]
- Bucki, R.; Pastore, J.J.; Randhawa, P.; Vegners, R.; Weiner, D.J.; Janmey, P.A. Antibacterial Activities of Rhodamine B-Conjugated Gelsolin-Derived Peptides Compared to Those of the Antimicrobial Peptides Cathelicidin LL37, Magainin II, and Melittin. Antimicrob. Agents Chemother. 2004, 48, 1526–1533. [Google Scholar] [CrossRef] [Green Version]
- Bucki, R.; Niemirowicz-Laskowska, K.; Deptuła, P.; Wilczewska, A.Z.; Misiak, P.; Durnaś, B.; Fiedoruk, K.; Piktel, E.; Mystkowska, J.; Janmey, P.A. Susceptibility of microbial cells to the modified PIP2-binding sequence of gelsolin anchored on the surface of magnetic nanoparticles. J. Nanobiotechnology 2019, 17, 81. [Google Scholar] [CrossRef] [Green Version]
- Bucki, R.; Wang, Y.H.; Yang, C.; Kandy, S.K.; Fatunmbi, O.; Bradley, R.; Pogoda, K.; Svitkina, T.; Radhakrishnan, R.; Janmey, P.A. Lateral distribution of phosphatidylinositol 4,5-bisphosphate in membranes regulates formin- and ARP2/3-mediated actin nucleation. J. Biol. Chem. 2019, 294, 4704–4722. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, R.; Ma, L.; Miki, H.; Lopez, M.; Kirchhausen, T.; Takenawa, T.; Kirschner, M.W. The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell 1999, 97, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, R.; Nollau, P.; Henry Ho, H.Y.; Kirschner, M.W.; Mayer, B.J. Nck and Phosphatidylinositol 4,5-Bisphosphate Synergistically Activate Actin Polymerization through the N-WASP-Arp2/3 Pathway. J. Biol. Chem. 2001, 276, 26448–26452. [Google Scholar] [CrossRef] [Green Version]
- Lassing, I.; Lindberg, U. Specific interaction between phosphatidylinositol 4,5-bisphosphate and profilactin. Nature 1985, 314, 472–474. [Google Scholar] [CrossRef]
- Bae, Y.H.; Ding, Z.; Das, T.; Wells, A.; Gertler, F.; Roy, P. Profilin1 regulates PI(3,4)P2 and lamellipodin accumulation at the leading edge thus influencing motility of MDA-MB-231 cells. Proc. Natl. Acad. Sci. USA 2010, 107, 21547–21552. [Google Scholar] [CrossRef] [Green Version]
- Goldschmidt-Clermont, P.J.; Machesky, L.M.; Baldassare, J.J.; Pollard, T.D. The actin-binding protein profilin binds to PIP2 and inhibits its hydrolysis by phospholipase C. Science 1990, 247, 1575–1578. [Google Scholar] [CrossRef]
- Goldschmidt-Clermont, P.J.; Furman, M.I.; Wachsstock, D.; Safer, D.; Nachmias, V.T.; Pollard, T.D. The control of actin nucleotide ex- change by thymosin beta 4 and profilin. A potential regulatory mechanism for actin polymerization in cells. Mol. Biol. Cell 1992, 3, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Kovar, D.R.; Kuhn, J.R.; Tichy, A.L.; Pollard, T.D. The fission yeast cytokinesis formin Cdc12p is a barbed end actin filament capping protein gated by profilin. J. Cell Biol. 2003, 161, 875–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, J.R.; Egile, C.; Gil, S.; Snapper, S.B.; Li, R.; Thomas, S.M. Cortactin regulates cell migration through activation of N-WASP. J. Cell Sci. 2005, 118, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Schafer, D.A.; Weed, S.A.; Binns, D.; Karginov, A.V.; Parsons, J.T.; Cooper, J.A. Dynamin2 and cortactin regulate actin assembly and filament organization. Curr. Biol. 2002, 12, 1852–1857. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, E.D.A.; Pinotsis, N.; Ghisleni, A.; Salmazo, A.; Konarev, P.V.; Kostan, J.; Sjöblom, B.; Schreiner, C.; Polyansky, A.A.; Gkougkoulia, E.A.; et al. The structure and regulation of human muscle α-Actinin. Cell 2014, 159, 1447–1460. [Google Scholar] [CrossRef] [Green Version]
- Pavalko, F.M.; Otey, C.A.; Simon, K.O.; Burridge, K. α-Actinin: A direct link between actin and integrins. Biochem. Soc. Trans. 1991, 19, 1065–1069. [Google Scholar] [CrossRef] [Green Version]
- Fukami, K.; Sawada, N.; Endo, T.; Takenawa, T. Identification of a Phosphatidylinositol 4,5-Bisphosphate-binding Site in Chicken Skeletal Muscle α-Actinin. J. Biol. Chem. 1996, 271, 2646–2650. [Google Scholar] [CrossRef] [Green Version]
- Sul, D.; Baron, C.B.; Broome, R.; Coburn, R.F. Smooth muscle length-dependent PI(4,5)P2 synthesis and paxillin tyrosine phosphorylation. Am. J. Physiol. Cell Physiol. 2001, 281, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Kallikourdis, M.; Trovato, A.E.; Roselli, G.; Muscolini, M.; Porciello, N.; Tuosto, L.; Viola, A. Phosphatidylinositol 4-Phosphate 5-Kinase β Controls Recruitment of Lipid Rafts into the Immunological Synapse. J. Immunol. 2016, 196, 1955–1963. [Google Scholar] [CrossRef] [Green Version]
- Shabardina, V.; Kramer, C.; Gerdes, B.; Braunger, J.; Cordes, A.; Schäfer, J.; Mey, I.; Grill, D.; Gerke, V.; Steinem, C. Mode of Ezrin-Membrane Interaction as a Function of PIP2 Binding and Pseudophosphorylation. Biophys. J. 2016, 110, 2710–2719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayasundar, J.J.; Ju, J.H.; He, L.; Liu, D.; Meilleur, F.; Zhao, J.; Callaway, D.J.E.; Bu, Z. Open conformation of ezrin bound to phosphatidylinositol 4,5-bisphosphate and to F-actin revealed by neutron scattering. J. Biol. Chem. 2012, 287, 37119–37133. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Mertz, B. Novel Phosphotidylinositol 4,5-Bisphosphate Binding Sites on Focal Adhesion Kinase. PLoS ONE 2015, 10, e0132833. [Google Scholar] [CrossRef] [Green Version]
- Goñi, G.M.; Epifano, C.; Boskovic, J.; Camacho-Artacho, M.; Zhou, J.; Bronowska, A.; Martín, M.T.; Eck, M.J.; Kremer, L.; Gräter, F.; et al. Phosphatidylinositol 4,5-bisphosphate triggers activation of focal adhesion kinase by inducing clustering and conformational changes. Proc. Natl. Acad. Sci. USA 2014, 111, E3177–E3186. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Bronowska, A.; Le Coq, J.; Lietha, D.; Gräter, F. Allosteric regulation of focal adhesion kinase by PIP2 and ATP. Biophys. J. 2015, 108, 698–705. [Google Scholar] [CrossRef] [Green Version]
- Mcnamee, H.P.; Liley, H.G.; Ingber, D.E. Integrin-Dependent Control of Inositol Lipid Synthesis in Vascular Endothelial Cells and Smooth Muscle Cells. Exp. Cell Res. 1996, 224, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Borowsky, M.L.; Hynes, R.O. Layilin, a novel talin-binding transmembrane protein homologous with C-type lectins, is localized in membrane ruffles. J. Cell Biol. 1998, 143, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, L.; Suzuki, A.; Lian, L.; Min, S.H.; Wang, Z.; Litvinov, R.I.; Stalker, T.J.; Yago, T.; Klopocki, A.G.; et al. Platelets lacking PIP5KIγ have normal integrin activation but impaired cytoskeletal-membrane integrity and adhesion. Blood 2013, 121, 2743–2752. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Ithychanda, S.; Qin, J.; Plow, E.F. Mechanisms of talin-dependent integrin signaling and crosstalk. Biochim. Biophys. Acta 2014, 1838, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.T.; Nygren, P.; Jo, H.; Boesze-Battaglia, K.; Bennett, J.S.; DeGrado, W.F. Affinity of talin-1 for the β3-integrin cytosolic domain is modulated by its phospholipid bilayer environment. Proc. Natl. Acad. Sci. USA 2012, 109, 793–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, P.R.; Goult, B.T.; Kopp, P.M.; Bate, N.; Grossmann, J.G.; Roberts, G.C.K.; Critchley, D.R.; Barsukov, I.L. The Structure of the Talin Head Reveals a Novel Extended Conformation of the FERM Domain. Structure 2010, 18, 1289–1299. [Google Scholar] [CrossRef] [Green Version]
- Thompson, P.M.; Ramachandran, S.; Case, L.B.; Tolbert, C.E.; Tandon, A.; Pershad, M.; Dokholyan, N.V.; Waterman, C.M.; Campbell, S.L. A Structural Model for Vinculin Insertion into PIP2-Containing Membranes and the Effect of Insertion on Vinculin Activation and Localization. Structure 2017, 25, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekar, I.; Stradal, T.E.B.; Holt, M.R.; Entschladen, F.; Jockusch, B.M.; Ziegler, W.H. Vinculin acts as a sensor in lipid regulation of adhesion-site turnover. J. Cell Sci. 2005, 118, 1461–1472. [Google Scholar] [CrossRef] [Green Version]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef]
- Wang, J.; Richards, D.A. Segregation of PIP2 and PIP3 into distinct nanoscale regions within the plasma membrane. Biol. Open 2012, 1, 857–862. [Google Scholar] [CrossRef] [Green Version]
- Van Den Bogaart, G.; Meyenberg, K.; Risselada, H.J.; Amin, H.; Willig, K.I.; Hubrich, B.E.; Dier, M.; Hell, S.W.; Grubmüller, H.; Diederichsen, U.; et al. Membrane protein sequestering by ionic protein-lipid interactions. Nature 2011, 479, 552–555. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, S. The Electrostatic Properties of Membranes. Ann. Rev. Biophys. Biophys. Chem. 1989, 18, 113–136. [Google Scholar] [CrossRef]
- Gambhir, A.; Hangyás-Mihályné, G.; Zaitseva, I.; Cafiso, D.S.; Wang, J.; Murray, D.; Pentyala, S.N.; Smith, S.O.; McLaughlin, S. Electrostatic sequestration of PIP2 on phospholipid membranes by basic/aromatic regions of proteins. Biophys. J. 2004, 86, 2188–2207. [Google Scholar] [CrossRef] [Green Version]
- Cevc, G.; Svetina, S.; Žekš, B. Electrostatic potential of bilayer lipid membranes with the structural surface charge smeared perpendicular to the membrane-solution interface. An extension of the Gouy-Chapman diffuse double layer theory. J. Phys. Chem. 1981, 85, 1762–1767. [Google Scholar] [CrossRef]
- Shapovalov, V.L.; Brezesinski, G. Breakdown of the Gouy—Chapman model for highly charged Langmuir monolayers: Counterion size effect. J. Phys. Chem. B 2006, 110, 10032–10040. [Google Scholar] [CrossRef]
- Li, Z.; Venable, R.M.; Rogers, L.A.; Murray, D.; Pastor, R.W. Molecular dynamics simulations of PIP2 and PIP3 in lipid bilayers: Determination of ring orientation, and the effects of surface roughness on a Poisson-Boltzmann description. Biophys. J. 2009, 97, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Slochower, D.R.; Huwe, P.J.; Radhakrishnan, R.; Janmey, P.A. Quantum and all-atom molecular dynamics simulations of protonation and divalent ion binding to phosphatidylinositol 4,5-bisphosphate (PIP2). J. Phys. Chem. B 2013, 117, 8322–8329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin, S.; Wang, J.; Gambhir, A.; Murray, D. PIP 2 and Proteins: Interactions, Organization, and Information Flow. Annu. Rev. Biophys. Biomol. Struct. 2002, 31, 151–175. [Google Scholar] [CrossRef] [Green Version]
- Levental, I.; Christian, D.A.; Wang, Y.H.; Madara, J.J.; Discher, D.E.; Janmey, P.A. Calcium-dependent lateral organization in phosphatidylinositol 4,5-bisphosphate (PIP2)- and cholesterol-containing monolayers. Biochemistry 2009, 48, 8241–8248. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Collins, A.; Guo, L.; Smith-Dupont, K.B.; Gai, F.; Svitkina, T.; Janmey, P.A. Divalent cation-induced cluster formation by polyphosphoinositides in model membranes. J. Am. Chem. Soc. 2012, 134, 3387–3395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, R.P.; Slochower, D.R.; Janmey, P.A.; Radhakrishnan, R. Divalent cations bind to phosphoinositides to induce ion and isomer specific propensities for nano-cluster initiation in bilayer membranes. R. Soc. Open Sci. 2020, 7, 192208. [Google Scholar] [CrossRef]
- Wen, Y.; Vogt, V.M.; Feigenson, G.W. Multivalent Cation-Bridged PI(4,5)P2 Clusters Form at Very Low Concentrations. Biophys. J. 2018, 114, 2630–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balla, T.; Bondeva, T.; Várnai, P. How accurately can we image inositol lipids in living cells? Trends Pharmacol. Sci. 2000, 21, 238–241. [Google Scholar] [CrossRef]
- Sorre, B.; Callan-Jones, A.; Manneville, J.B.; Nassoy, P.; Joanny, J.F.; Prost, J.; Goud, B.; Bassereau, P. Curvature-driven lipid sorting needs proximity to a demixing point and is aided by proteins. Proc. Natl. Acad. Sci. USA 2009, 106, 5622–5626. [Google Scholar] [CrossRef] [Green Version]
- Roux, A.; Koster, G.; Lenz, M.; Sorre, B.; Manneville, J.B.; Nassoy, P.; Bassereau, P. Membrane curvature controls dynamin polymerization. Proc. Natl. Acad. Sci. USA 2010, 107, 4141–4146. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Wang, H.; Lou, Z.; Cao, M.; Zhang, Z.; Gu, N. Roles of PIP 2 in the membrane binding of MIM I- BAR: Insights from molecular dynamics simulations. FEBS Lett. 2018, 592, 2533–2542. [Google Scholar] [CrossRef] [Green Version]
- Simunovic, M.; Manneville, J.-B.; Renard, H.-F.; Evergren, E.; Raghunathan, K.; Bhatia, D.; Kenworthy, A.K.; Voth, G.A.; Prost, J.; McMahon, H.T.; et al. Friction Mediates Scission of Tubular Membranes Scaffolded by BAR Proteins. Cell 2017, 170, 172–184.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Baumgart, T. BIN1 Membrane Curvature Sensing and Generation Show Autoinhibition Regulated by Downstream Ligands and PI(4,5)P 2. Biochemistry 2014. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Shi, Z.; Baumgart, T. Mutations in BIN1 Associated with Centronuclear Myopathy Disrupt Membrane Remodeling by Affecting Protein Density and Oligomerization. PLoS ONE 2014, 9, e93060. [Google Scholar] [CrossRef]
- Nicot, A.S.; Toussaint, A.; Tosch, V.; Kretz, C.; Wallgren-Pettersson, C.; Iwarsson, E.; Kingston, H.; Garnier, J.M.; Biancalana, V.; Oldfors, A.; et al. Mutations in amphiphysin 2 (BIN1) disrupt interaction with dynamin 2 and cause autosomal recessive centronuclear myopathy. Nat. Genet. 2007, 39, 1134–1139. [Google Scholar] [CrossRef]
- Brown, D.A.; Hughes, S.A.; Marsh, S.J.; Tinker, A. Regulation of M(Kv7.2/7.3) channels in neurons by PIP 2 and products of PIP 2 hydrolysis: Significance for receptor-mediated inhibition. J. Physiol. 2007, 582, 917–925. [Google Scholar] [CrossRef]
- Ebi, H.; Costa, C.; Faber, A.C.; Nishtala, M.; Kotani, H.; Juric, D.; Della Pelle, P.; Song, Y.; Yano, S.; Mino-Kenudson, M.; et al. PI3K regulates MEK/ERK signaling in breast cancer via the Rac-GEF, P-Rex1. Proc. Natl. Acad. Sci. USA 2013, 110, 21124–21129. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Prakash, P.; Gorfe, A.A.; Hancock, J.F. Ras and the Plasma Membrane: A Complicated Relationship. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Falkenburger, B.H.; Jensen, J.B.; Dickson, E.J.; Suh, B.C.; Hille, B. Phosphoinositides: Lipid regulators of membrane proteins. J. Physiol. 2010, 588, 3179–3185. [Google Scholar] [CrossRef]
- Szentpetery, Z.; Várnai, P.; Balla, T. Acute manipulation of Golgi phosphoinositides to assess their importance in cellular trafficking and signaling. Proc. Natl. Acad. Sci. USA 2010, 107, 8225–8230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, P.R.; Owen, D.J. Endocytosis and vesicle trafficking. Curr. Opin. Struct. Biol. 2002, 12, 814–821. [Google Scholar] [CrossRef]
- Posor, Y.; Eichhorn-Gruenig, M.; Puchkov, D.; Schöneberg, J.; Ullrich, A.; Lampe, A.; Müller, R.; Zarbakhsh, S.; Gulluni, F.; Hirsch, E.; et al. Spatiotemporal control of endocytosis by phosphatidylinositol-3,4- bisphosphate. Nature 2013, 499, 233–237. [Google Scholar] [CrossRef]
- Goud, B. Why does endocytosis in single cells care which side up? Bioarchitecture 2014, 4, 62–67. [Google Scholar]
- Jost, M.; Simpson, F.; Kavran, J.M.; Lemmon, M.A.; Schmid, S.L. Phosphatidylinositol-4,5-bisphosphate is required for endocytic coated vesicle formation. Curr. Biol. 1998, 8, 1399–1404. [Google Scholar] [CrossRef] [Green Version]
- Czech, M.P. PIP2 and PIP3: Complex roles at the cell surface. Cell 2000, 100, 603–606. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Hedman, A.C.; Tan, X.; Schill, N.J.; Anderson, R.A. Endosomal Type Iγ PIP 5-Kinase Controls EGF Receptor Lysosomal Sorting. Dev. Cell 2013, 25, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Gullapalli, A.; Garrett, T.A.; Paing, M.M.; Griffin, C.T.; Yang, Y.; Trejo, J.A. A Role for Sorting Nexin 2 in Epidermal Growth Factor Receptor Down-regulation: Evidence for Distinct Functions of Sorting Nexin 1 and 2 in Protein Trafficking. Mol. Biol. Cell 2004, 15, 2143–2155. [Google Scholar] [CrossRef] [Green Version]
- Merino-Trigo, A.; Kerr, M.C.; Houghton, F.; Lindberg, A.; Mitchell, C.; Teasdale, R.D.; Gleeson, P.A. Sorting nexin 5 is localized to a subdomain of the early endosomes and is recruited to the plasma membrane following EGF stimulation. J. Cell Sci. 2004, 117, 6413–6424. [Google Scholar] [CrossRef] [Green Version]
- Cheung, M.; Sharma, A.; Madhunapantula, S.R.V.; Robertson, G.P. Akt3 and mutant V600EB-Raf cooperate to promote early melanoma development. Cancer Res. 2008, 68, 3429–3439. [Google Scholar] [CrossRef] [Green Version]
- Amblard, I.; Dupont, E.; Alves, I.; Miralves, J.; Queguiner, I.; Joliot, A. Bidirectional transfer of homeoprotein EN2 across the plasma membrane requires PIP2. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef]
- De Matteis, M.A.; Godi, A. PI-loting membrane traffic. Nat. Cell Biol. 2004, 6, 487–492. [Google Scholar] [CrossRef]
- De Craene, J.O.; Bertazzi, D.L.; Bär, S.; Friant, S. Phosphoinositides, major actors in membrane trafficking and lipid signaling pathways. Int. J. Mol. Sci. 2017, 18, 634. [Google Scholar] [CrossRef]
- Krauß, M.; Haucke, V. Phosphoinositide-metabolizing enzymes at the interface between membrane traffic and cell signalling. EMBO Rep. 2007, 8, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Balla, T.; Várnai, P. Visualization of cellular phosphoinositide pools with GFP-fused protein-domains. Curr. Protoc. Cell Biol. 2009, 42, 24.4.1–24.4.27. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Du, X.N.; Jia, Q.Z.; Zhang, H.L. Binding of PLCδ1PH-GFP to Ptdlns(4,5)P2 prevents inhibition of phospholipase C-mediated hydrolysis of Ptdlns(4,5)P 2 by neomycin. Acta Pharmacol. Sin. 2005, 26, 1485–1491. [Google Scholar] [CrossRef] [Green Version]
- Poli, A.; Zaurito, A.E.; Abdul-Hamid, S.; Fiume, R.; Faenza, I.; Divecha, N. Phosphatidylinositol 5 phosphate (Pi5p): From behind the scenes to the front (nuclear) stage. Int. J. Mol. Sci. 2019, 20, 2080. [Google Scholar] [CrossRef] [Green Version]
- Gary, J.D.; Wurmser, A.E.; Bonangelino, C.J.; Weisman, L.S.; Emr, S.D. Fab1p is essential for PtdIns(3)P 5-kinase activity and the maintenance of vacuolar size and membrane homeostasis. J. Cell Biol. 1998, 143, 65–79. [Google Scholar] [CrossRef]
- Eugster, A.; Pécheur, E.I.; Michel, F.; Winsor, B.; Letourneur, F.; Friant, S. Ent5p is required with Ent3p and Vps27p for ubiquitin-dependent protein sorting into the multivesicular body. Mol. Biol. Cell 2004, 15, 3031–3041. [Google Scholar] [CrossRef]
- Chalhoub, N.; Baker, S.J. PTEN and the PI3-kinase pathway in cancer. Annu. Rev. Pathol. Mech. Dis. 2009, 4, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, M.W.; Homanics, G.E.; Lazo, J.S. Targeted Deletion of the Metastasis-Associated Phosphatase Ptp4a3 (PRL-3) Suppresses Murine Colon Cancer. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Marshall, A.J. Phosphatidylinositol (3,4) bisphosphate-specific phosphatases and effector proteins: A distinct branch of PI3K signaling. Cell. Signal. 2015, 27, 1789–1798. [Google Scholar] [CrossRef]
- Cheung, S.M.S.; Kornelson, J.C.; Al-Alwan, M.; Marshall, A.J. Regulation of phosphoinositide 3-kinase signaling by oxidants: Hydrogen peroxide selectively enhances immunoreceptor-induced recruitment of phosphatidylinositol (3,4) bisphosphate-binding PH domain proteins. Cell. Signal. 2007, 19, 902–912. [Google Scholar] [CrossRef]
- Cunningham, C.C.; Vegners, R.; Bucki, R.; Funaki, M.; Korde, N.; Hartwig, J.H.; Stossel, T.P.; Janmey, P.A. Cell permeant polyphosphoinositide-binding peptides that block cell motility and actin assembly. J. Biol. Chem. 2001, 276, 43390–43399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, R.; Diao, A.; Zhang, F.; Eisenberg, E.; Saint-Pol, A.; Williams, C.; Konstantakopoulos, A.; Lucocq, J.; Johannes, L.; Rabouille, C.; et al. Lowe syndrome protein OCRL1 interacts with clathrin and regulates protein trafficking between endosomes and the trans-Golgi network. Mol. Biol. Cell 2005, 16, 3467–3479. [Google Scholar] [CrossRef] [Green Version]
- Vicinanza, M.; Di Campli, A.; Polishchuk, E.; Santoro, M.; Di Tullio, G.; Godi, A.; Levtchenko, E.; De Leo, M.G.; Polishchuk, R.; Sandoval, L.; et al. OCRL controls trafficking through early endosomes via PtdIns4,5P 2-dependent regulation of endosomal actin. EMBO J. 2011, 30, 4970–4985. [Google Scholar] [CrossRef] [PubMed]
- Cremona, O.; Di Paolo, G.; Wenk, M.R.; Lüthi, A.; Kim, W.T.; Takei, K.; Daniell, L.; Nemoto, Y.; Shears, S.B.; Flavell, R.A.; et al. Essential role of phosphoinositide metabolism in synaptic vesicle recycling. Cell 1999, 99, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Mücksch, F.; Citir, M.; Lüchtenborg, C.; Glass, B.; Traynor-Kaplan, A.; Schultz, C.; Brügger, B.; Kräusslich, H.G. Quantification of phosphoinositides reveals strong enrichment of PIP2 in HIV-1 compared to producer cell membranes. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Rocha-Perugini, V.; Gordon-Alonso, M.; Sánchez-Madrid, F. PIP2: Choreographer of actin-adaptor proteins in the HIV-1 dance. Trends Microbiol. 2014, 22, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Arancio, O. PIP2: A new key player in Alzheimer’s disease. Cellscience 2008, 5, 44–47. [Google Scholar]
- Duciel, L.; Anezo, O.; Mandal, K.; Laurent, C.; Planque, N.; Coquelle, F.M.; Gentien, D.; Manneville, J.-B.; Saule, S. Protein tyrosine phosphatase 4A3 (PTP4A3/PRL-3) promotes the aggressiveness of human uveal melanoma through dephosphorylation of CRMP2. Sci. Rep. 2019, 9, 2990. [Google Scholar] [CrossRef]
- Mandal, K.; Gong, Z.; Rylander, A.; Shenoy, V.B.; Janmey, P. Opposite responses of normal hepatocytes and hepatocellular carcinoma cells to substrate viscoelasticity. Biomater. Sci. 2020, 8, 1316–1328. [Google Scholar] [CrossRef]
- Lan, T.; Pang, J.; Wu, Y.; Zhu, M.; Yao, X.; Wu, M.; Qian, H.; Zhang, Z.; Gao, J.; Chen, Y. Cross-linked hyaluronic acid gel inhibits metastasis and growth of gastric and hepatic cancer cells: In vitro and in vivo studies. Oncotarget 2016, 7, 65418–65428. [Google Scholar] [CrossRef]
- Guillou, H.; Lécureuil, C.; Anderson, K.E.; Suire, S.; Ferguson, G.J.; Ellson, C.D.; Gray, A.; Divecha, N.; Hawkins, P.T.; Stephens, L.R. Use of the GRP1 PH domain as a tool to measure the relative levels of PtdIns(3,4,5)P3 through a protein-lipid overlay approach. J. Lipid Res. 2007, 48, 726–732. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Shao, R.; Zhang, X.D.; Chen, C. Applications of nanotechnology for melanoma treatment, diagnosis, and theranostics. Int. J. Nanomed. 2013, 8, 2677–2688. [Google Scholar] [CrossRef] [Green Version]
- McParland, V.; Varsano, G.; Li, X.; Thornton, J.; Baby, J.; Aravind, A.; Meyer, C.; Pavic, K.; Rios, P.; Köhn, M. The metastasis-promoting phosphatase PRL-3 shows activity toward phosphoinositides. Biochemistry 2011, 50, 7579–7590. [Google Scholar] [CrossRef]
- Liu, J.; Gardel, M.L.; Kroy, K.; Frey, E.; Hoffman, B.D.; Crocker, J.C.; Bausch, A.R.; Weitz, D.A. Microrheology Probes Length Scale Dependent Rheology. Phys. Rev. Lett. 2005. [Google Scholar] [CrossRef] [Green Version]
- Daly, R.J. Cortactin signalling and dynamic actin networks. Biochem. J. 2004, 382, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Isoforms of phosphoinositides. By the action of PIK and phosphatase, phosphatidylinositol (PtdIns) and the three isoforms of PIP2 are formed, as indicated here. The specific action of PI3K I, II III and of the 3-phosphatases are also illustrated.
Figure 1.
Isoforms of phosphoinositides. By the action of PIK and phosphatase, phosphatidylinositol (PtdIns) and the three isoforms of PIP2 are formed, as indicated here. The specific action of PI3K I, II III and of the 3-phosphatases are also illustrated.
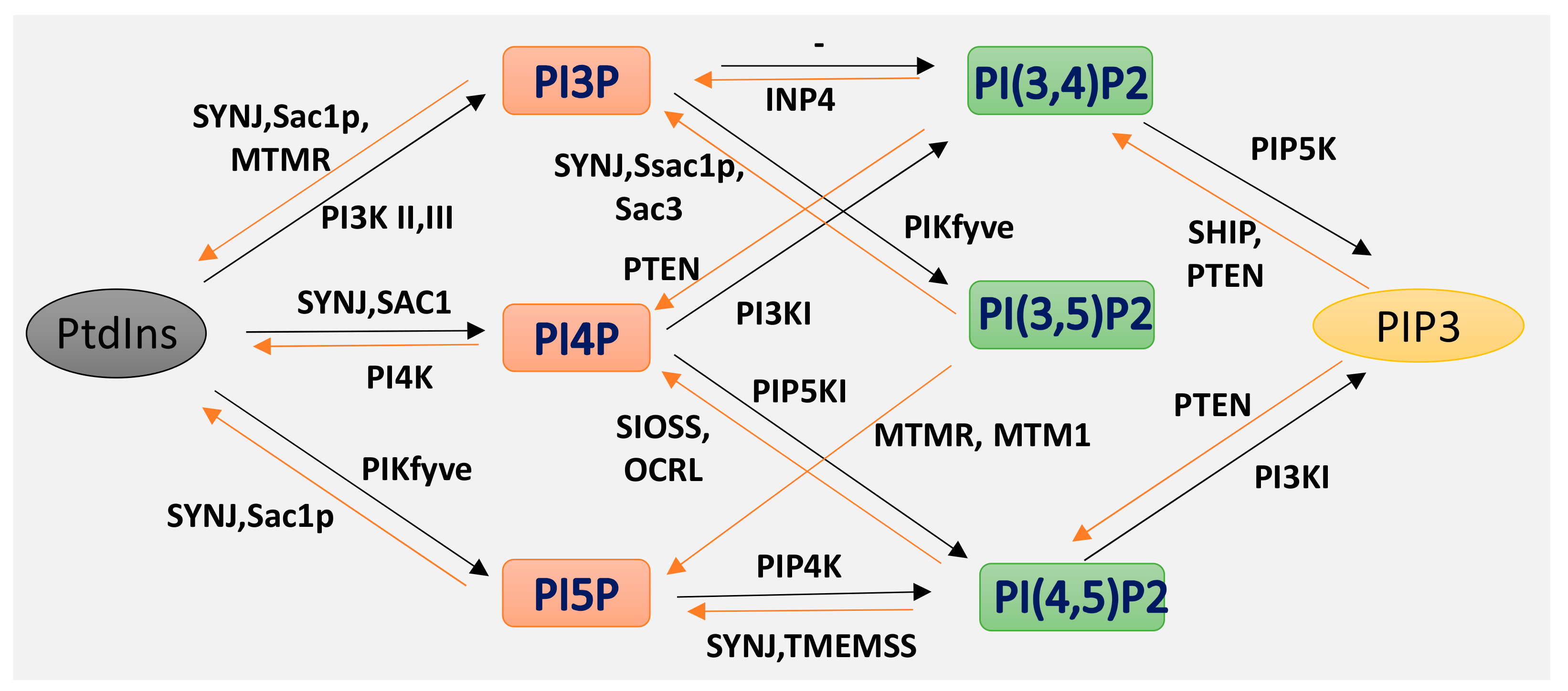
Figure 2.
Role of PIP2 in actin dynamics either by promoting polymerization or inhibiting severing. The figure summarizes gelsolin, profilin, cofilin, Arp2/3, and WASP dynamics in coordination with Rho- ROCK and Rac pathways.
Figure 2.
Role of PIP2 in actin dynamics either by promoting polymerization or inhibiting severing. The figure summarizes gelsolin, profilin, cofilin, Arp2/3, and WASP dynamics in coordination with Rho- ROCK and Rac pathways.

Figure 3.
Role of PIP2 in regulating focal adhesion assembly. Depiction of adhesion molecules talin, vinculin, ezrin, filamin and a-actinin. PIP2 synergistically binds to both talin and integrin and activates both of them. Talin binds directly to actin or activates vinculin and facilitates its binding to actin. PIP2 also binds to FERM domain of FAK and binds to vinculin via paxillin. PIP2 negatively regulates cross-linking activity of filamin and the actin bundle formation mediated by α-actinin.
Figure 3.
Role of PIP2 in regulating focal adhesion assembly. Depiction of adhesion molecules talin, vinculin, ezrin, filamin and a-actinin. PIP2 synergistically binds to both talin and integrin and activates both of them. Talin binds directly to actin or activates vinculin and facilitates its binding to actin. PIP2 also binds to FERM domain of FAK and binds to vinculin via paxillin. PIP2 negatively regulates cross-linking activity of filamin and the actin bundle formation mediated by α-actinin.
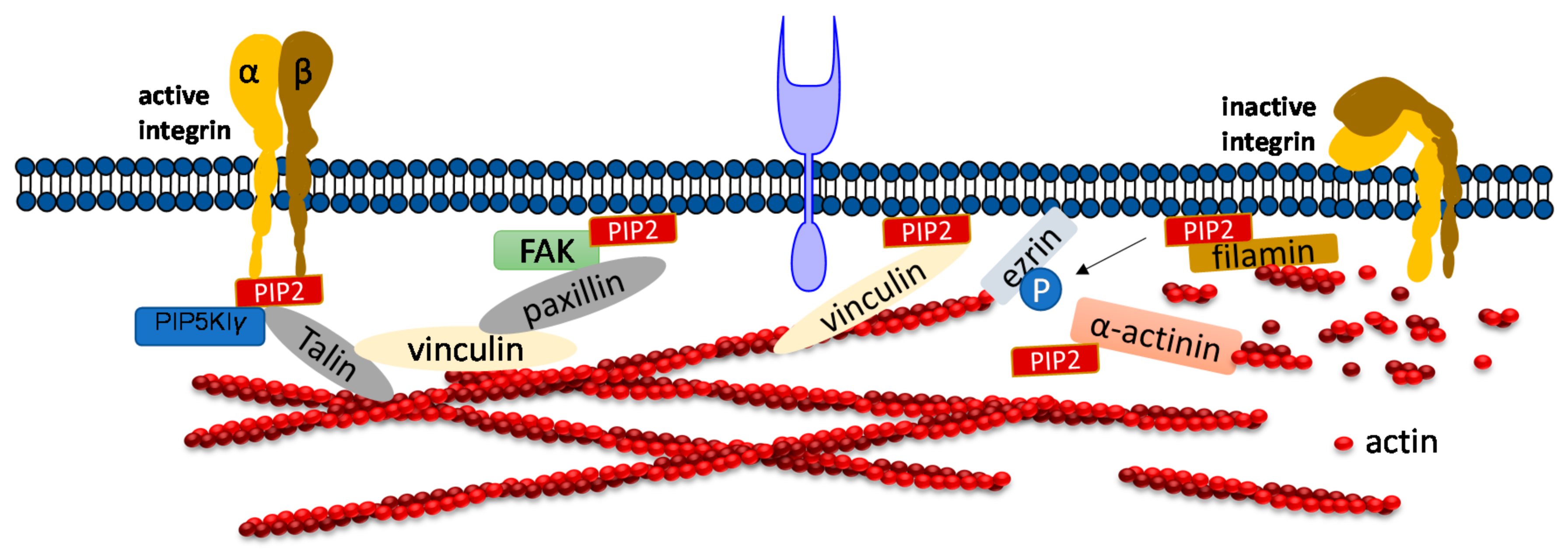
Figure 4.
Histograms of cluster of lipids which is also measured on the vertical axis. Only the unique combination of PI(4, 5)P2 and Ca2+ shows large and growing clusters. The symbol area is proportional to the number of lipids in the cluster (Bradley et al.) [95].
Figure 4.
Histograms of cluster of lipids which is also measured on the vertical axis. Only the unique combination of PI(4, 5)P2 and Ca2+ shows large and growing clusters. The symbol area is proportional to the number of lipids in the cluster (Bradley et al.) [95].
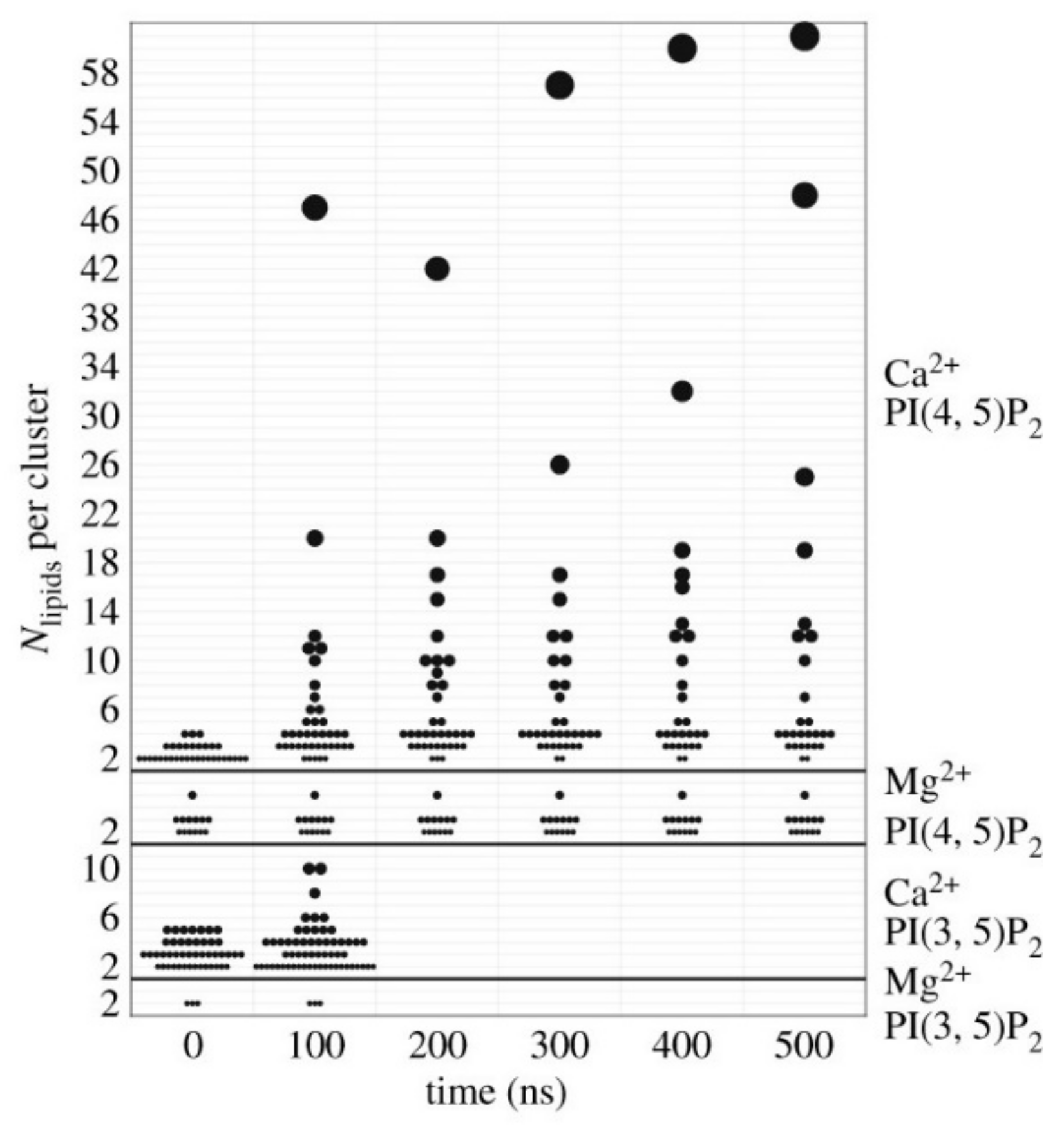
Figure 5.
PIP 2 molecules are necessary to recruit MIM I-BAR, which in turn can induce local PIP 2 clustering at its two ends after binding to the membrane (upper panel). Spontaneous bending of lipid membranes can re-distribute PIP 2 molecules to the negatively curved membrane areas (lower panel), which promotes the recruitment of MIM I-BAR and maintaining the curvature [100].
Figure 5.
PIP 2 molecules are necessary to recruit MIM I-BAR, which in turn can induce local PIP 2 clustering at its two ends after binding to the membrane (upper panel). Spontaneous bending of lipid membranes can re-distribute PIP 2 molecules to the negatively curved membrane areas (lower panel), which promotes the recruitment of MIM I-BAR and maintaining the curvature [100].

Figure 6.
BIN1 mediated membrane tubulation. BAR domain proteins are able to both sense and induce membrane curvature. BIN1 clustering with PIP2 that promote dynamine recruitment and thus forms T- tubule. Binding of dynamin depends upon the amount of PI(4, 5)P2 and enhanced by BIN1.
Figure 6.
BIN1 mediated membrane tubulation. BAR domain proteins are able to both sense and induce membrane curvature. BIN1 clustering with PIP2 that promote dynamine recruitment and thus forms T- tubule. Binding of dynamin depends upon the amount of PI(4, 5)P2 and enhanced by BIN1.
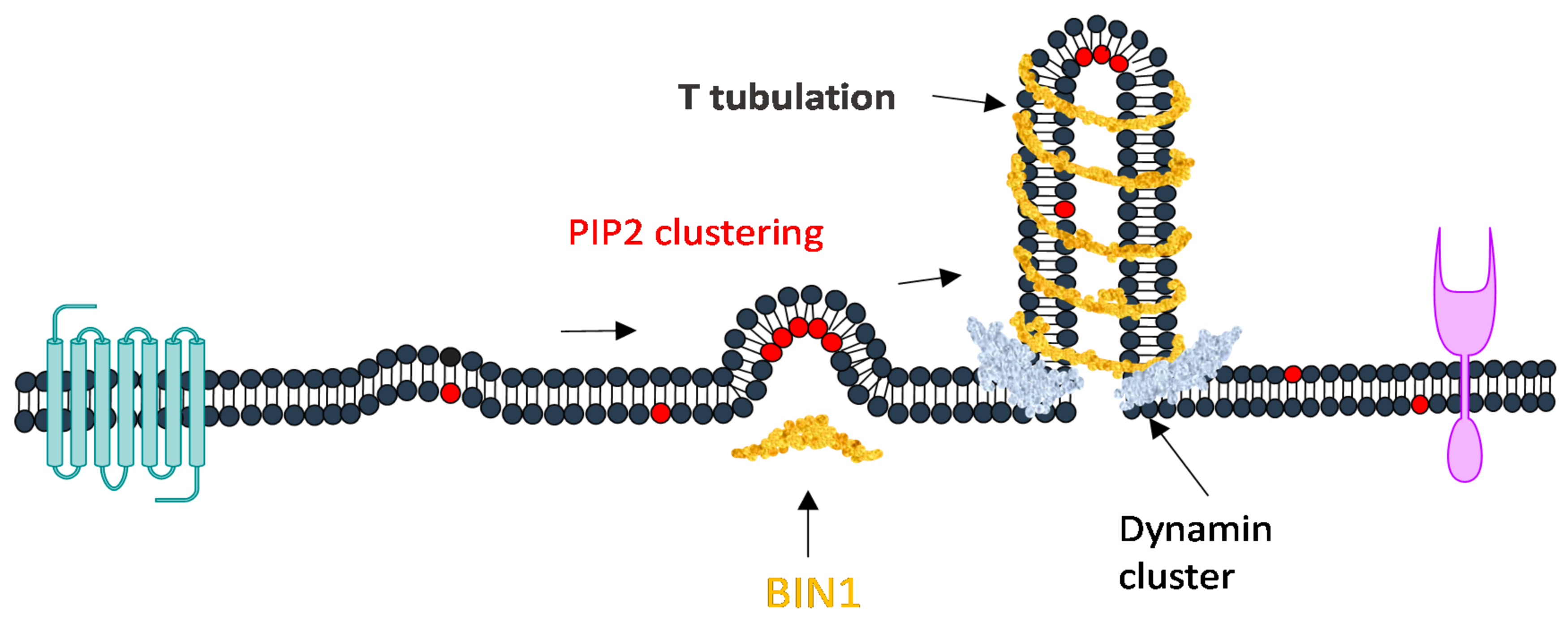
Figure 7.
PIP2 is involved in intracellular trafficking and vesicular transport. PIP2 participates in both clathrin mediated (CCP) and non- clathrin mediated endocytosis. PI(3,5) P2 is involved in exocytosis whereas PI(4,5) P2 and PI(3,4) P2 are involved in endocytic processes.
Figure 7.
PIP2 is involved in intracellular trafficking and vesicular transport. PIP2 participates in both clathrin mediated (CCP) and non- clathrin mediated endocytosis. PI(3,5) P2 is involved in exocytosis whereas PI(4,5) P2 and PI(3,4) P2 are involved in endocytic processes.

Figure 8.
PIP3 and MAP kinase pathways synergistically and independently regulates melanoma cancer or any other carcinoma. Ras regulates both PI3 and Akt kinase pathways. In addition, Ras independently regulates BRAF which is also implicated in Akt3 activity depicted in diagram.
Figure 8.
PIP3 and MAP kinase pathways synergistically and independently regulates melanoma cancer or any other carcinoma. Ras regulates both PI3 and Akt kinase pathways. In addition, Ras independently regulates BRAF which is also implicated in Akt3 activity depicted in diagram.
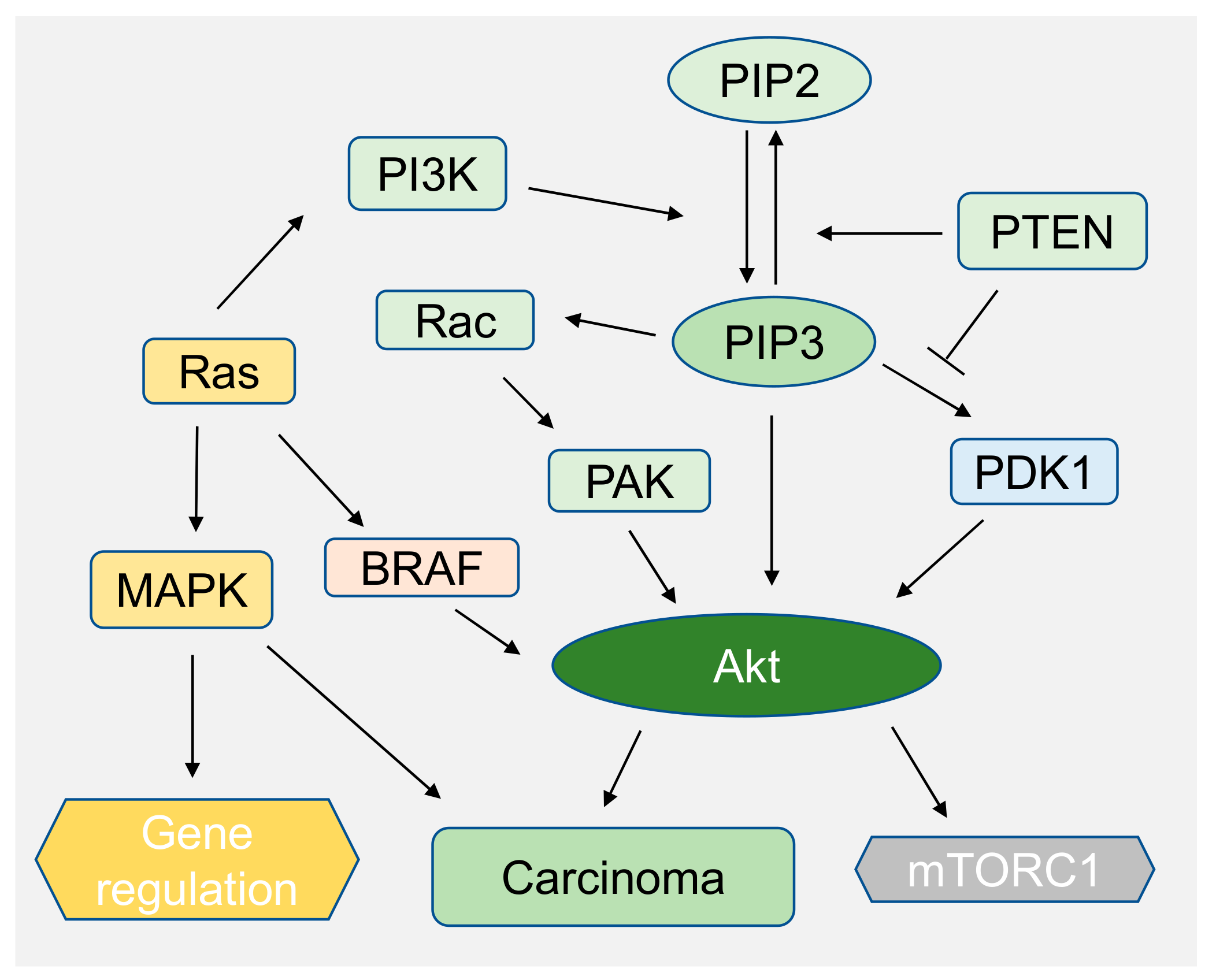

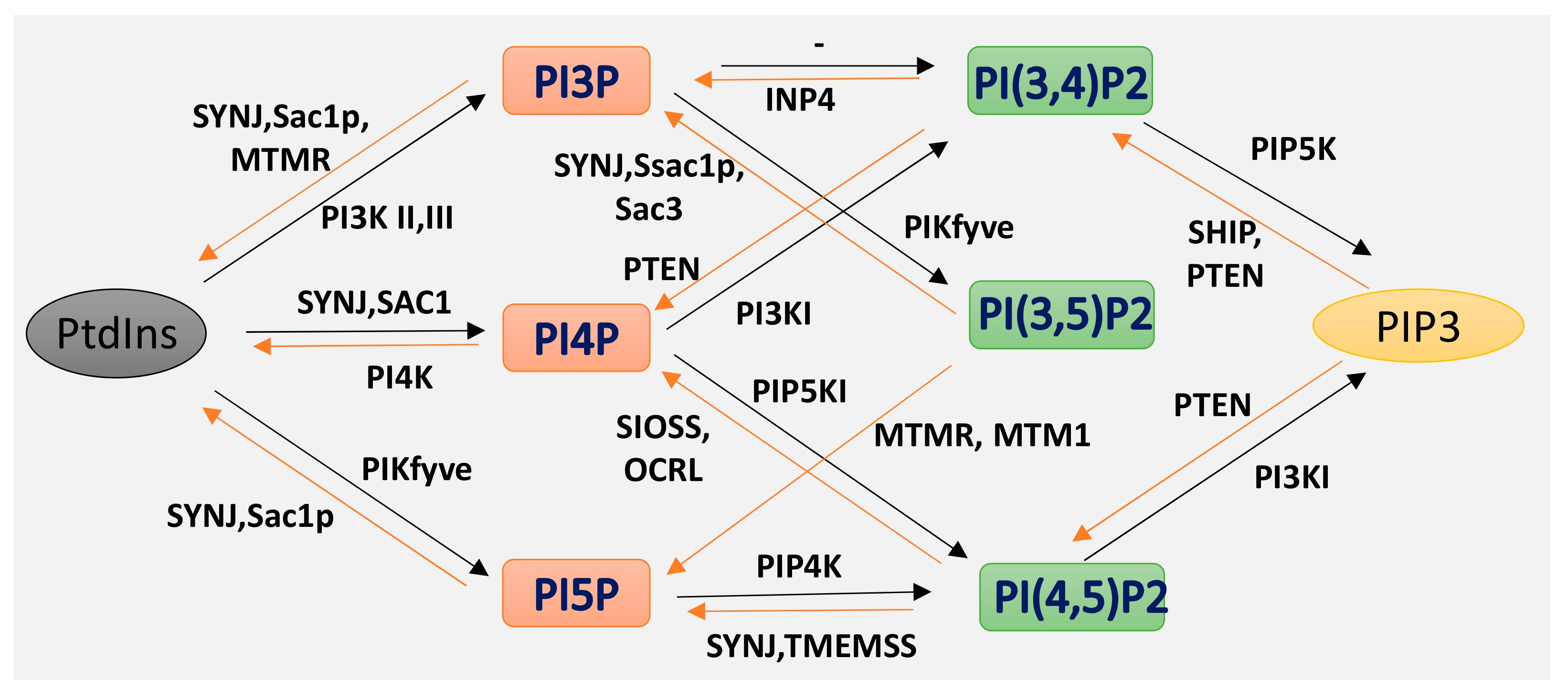{kind=link}
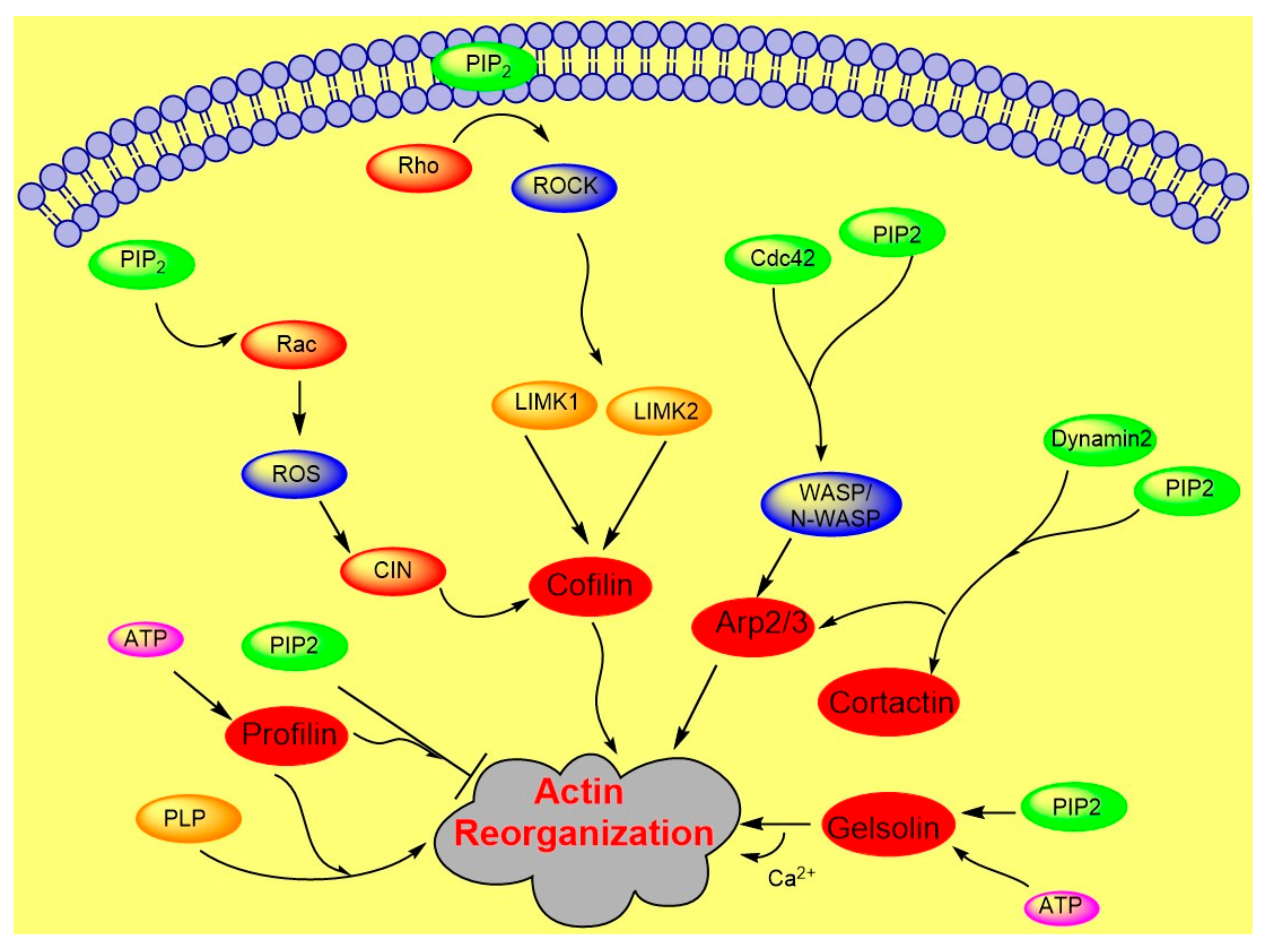{kind=link}
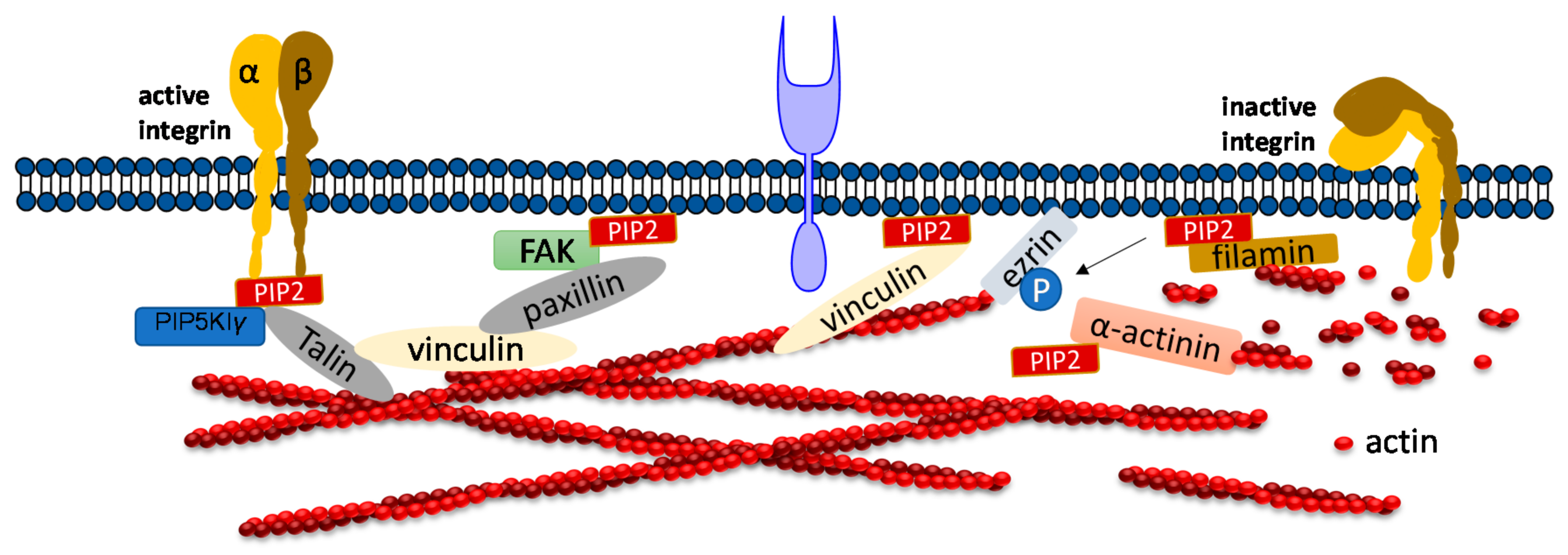{kind=link}
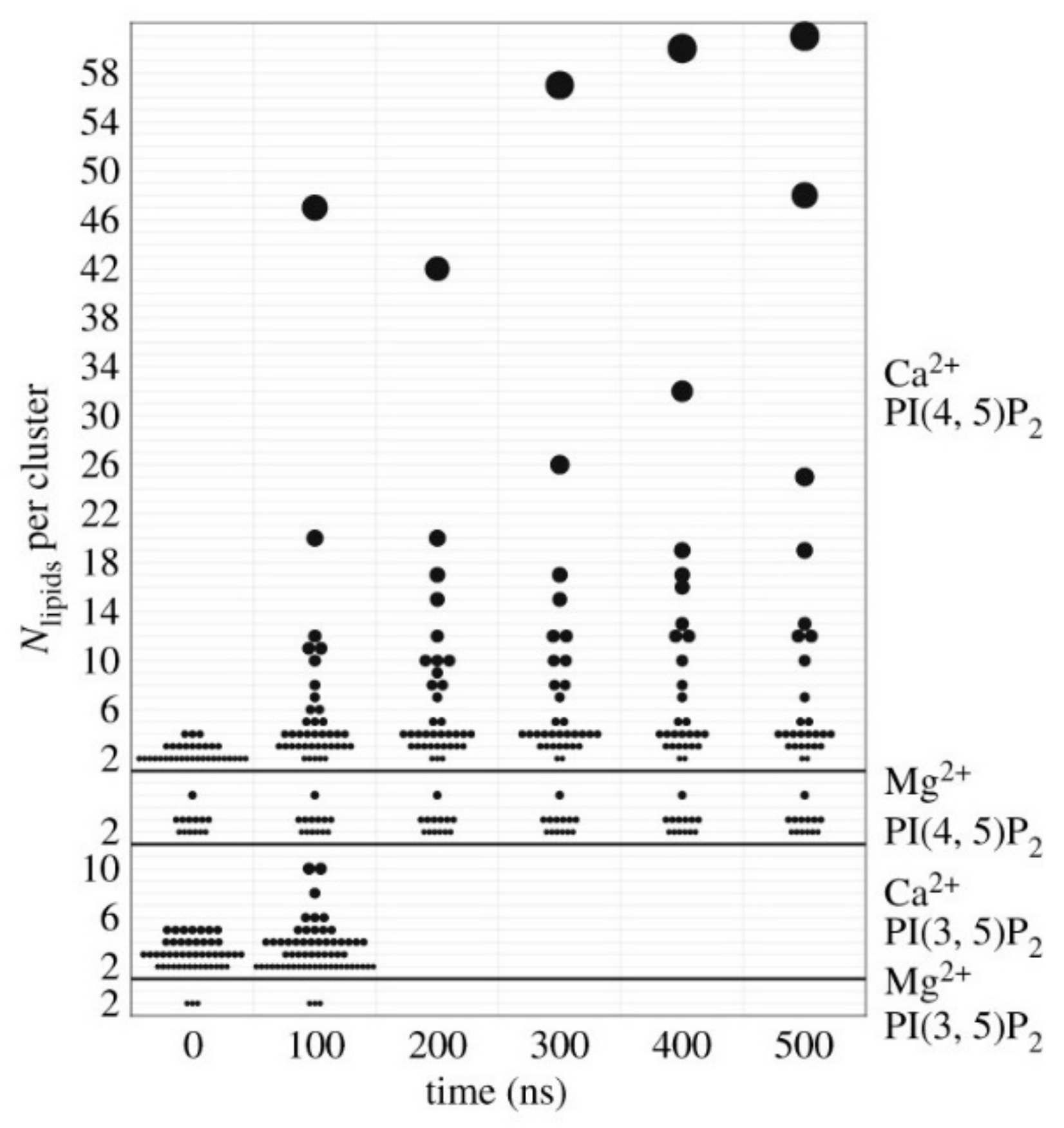{kind=link}
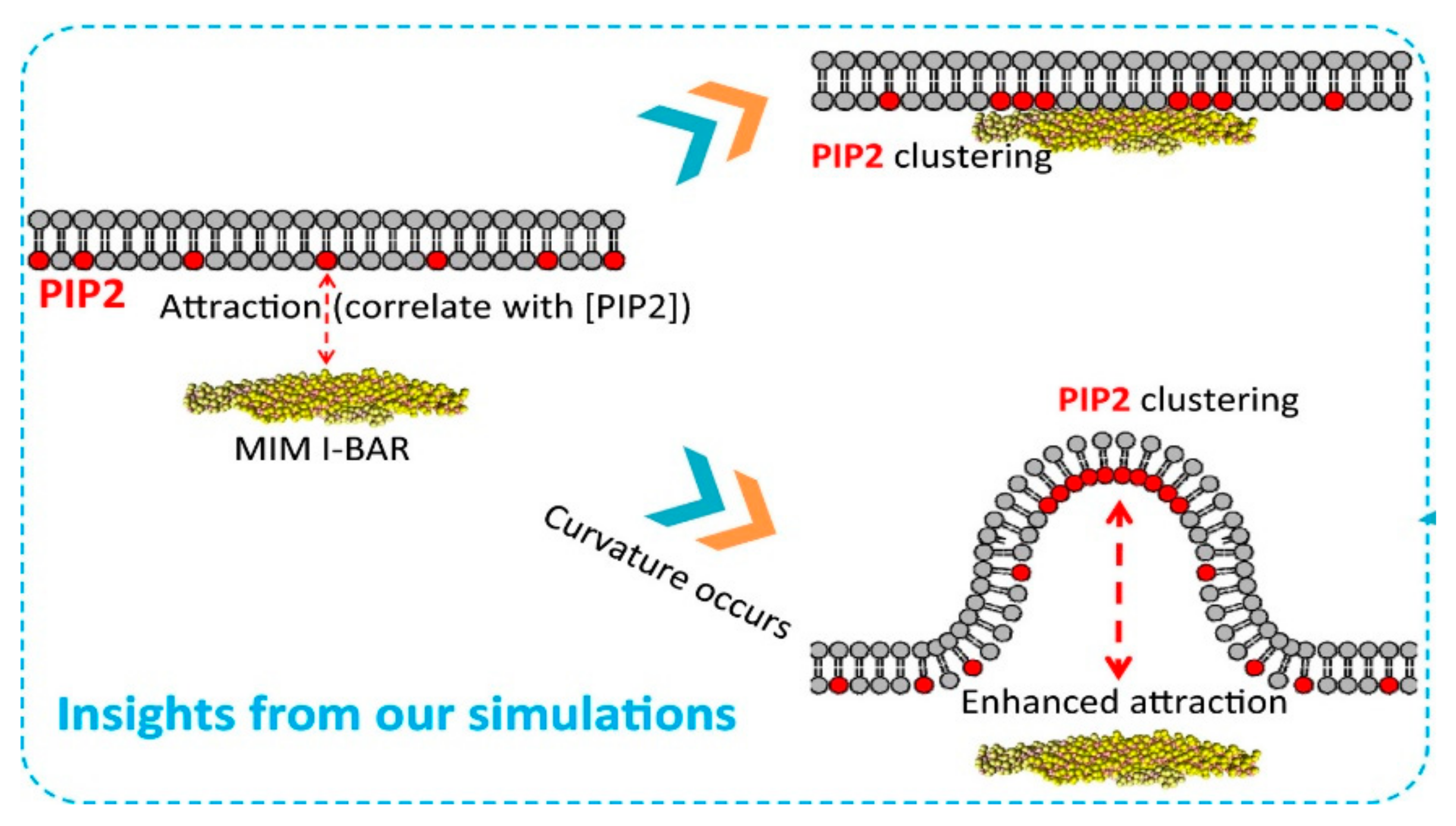{kind=link}
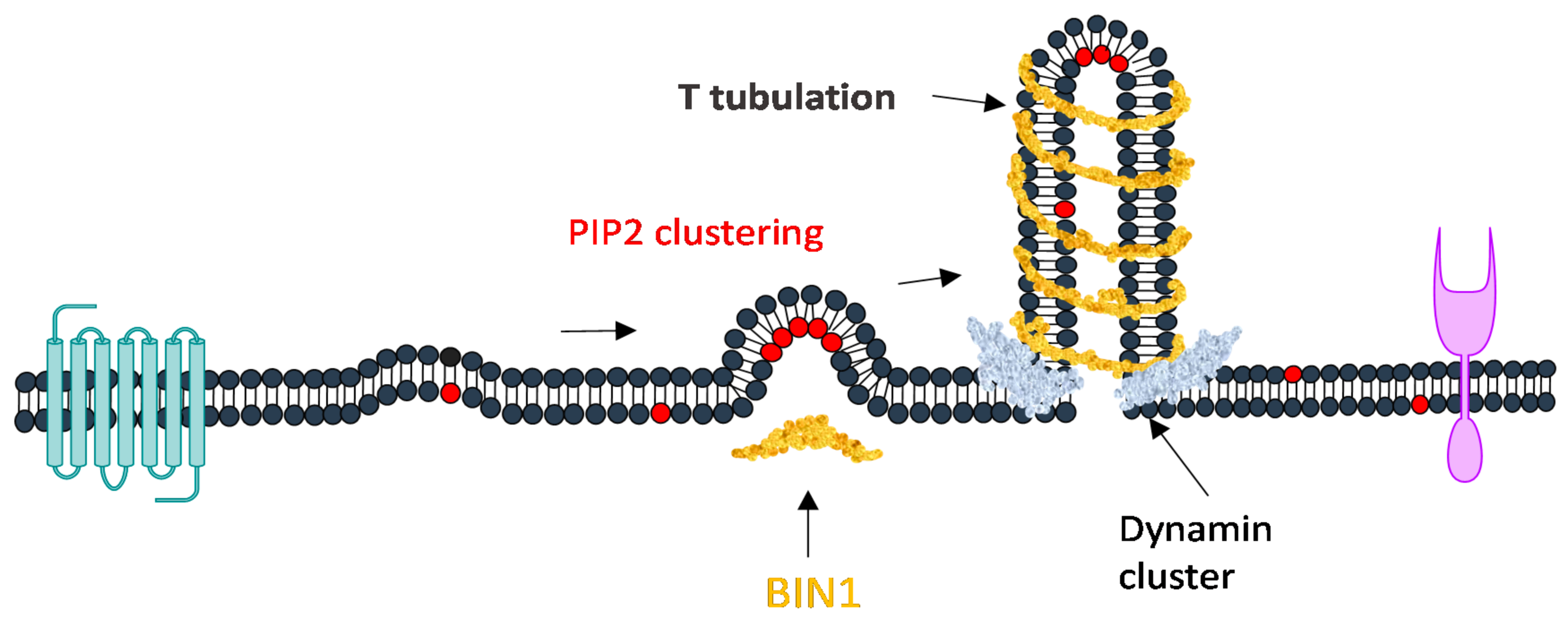{kind=link}
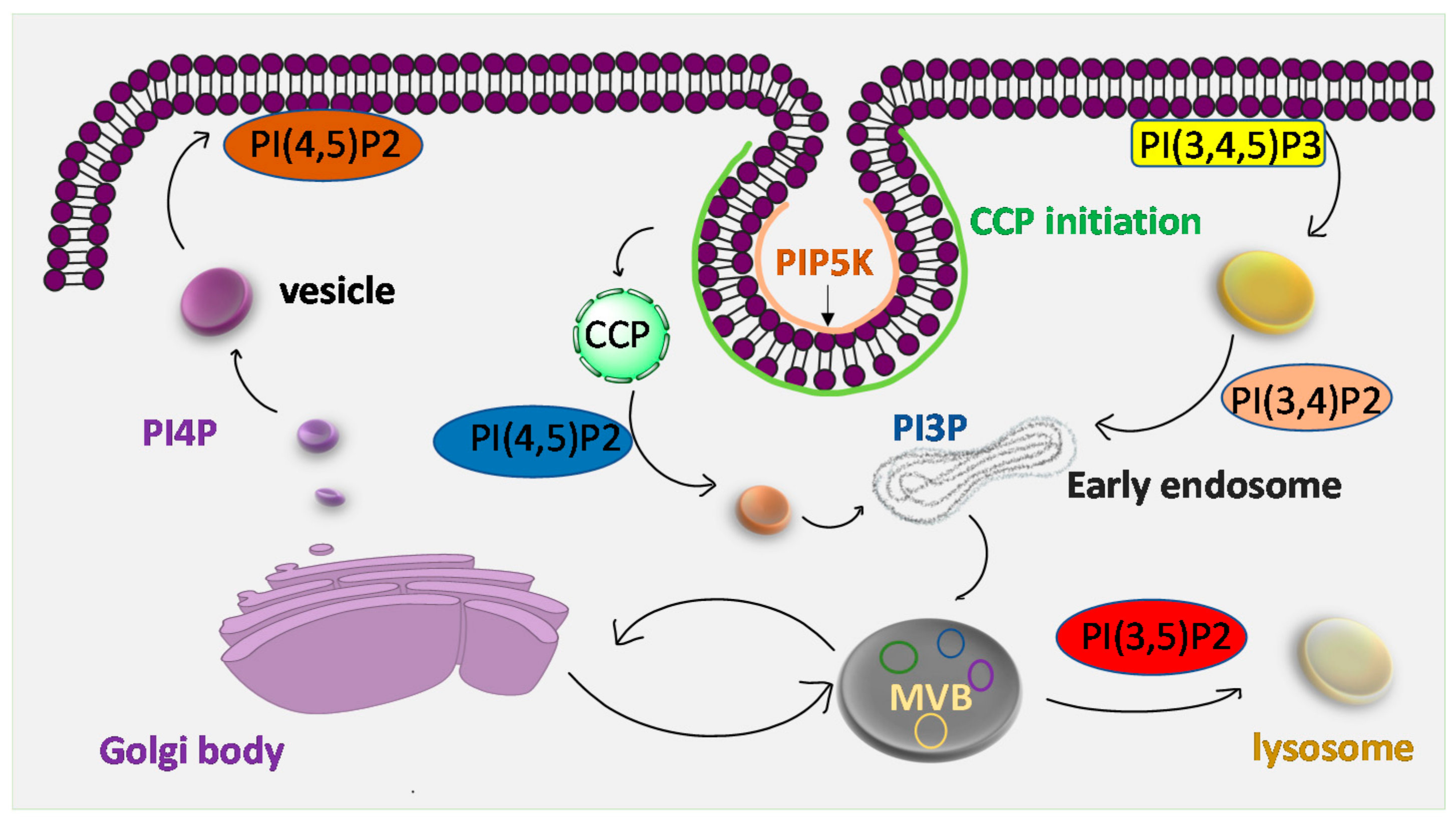{kind=link}
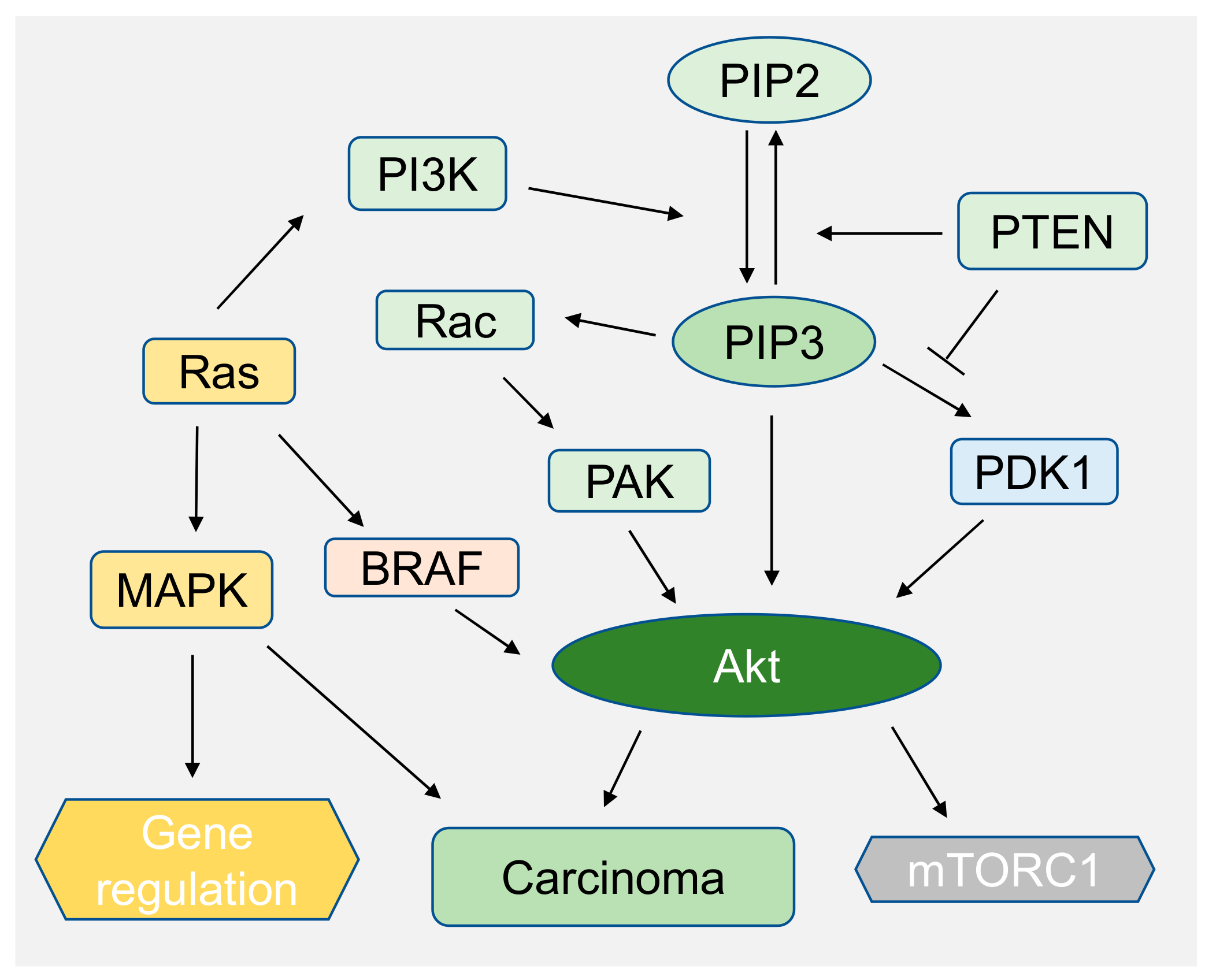{kind=link}
Table 1.
PIP2 and enzymatic activity in different pathways in disease [122].
Table 1.
PIP2 and enzymatic activity in different pathways in disease [122].
| Phospho-Inositides | Pathways/Functions | Enzymatic Activity | Disease Implication | References |
|---|---|---|---|---|
| PIP3, PI(4,5) P2, PI(3,5) P2 | PI3K-Akt | PI3K, PTEN 1,2 | Melanoma cancer, Cowden disease, pancreatic cancer, ovarian cancer. | [132] |
| PI(4,5) P2 | Endocytic trafficking pathways | OCRL, 5 phosphatases | Oculo-cerebro-renal syndrome of Lowe: renal Fancomi syndrome, glaucoma, cataracts, blindness, mental retardation. | [133,134] |
| PI(3,5) P2 | MTM1, PI4P, | Myopathy. | [2,121] | |
| PI(3,5) P2 | Fab1/PIKfyve kinase | Neuropathologies, Charcot-Marie tooth disease. | ||
| PI(4,5) P2, PI(3,5) P2 | Endocytic pathways | Synaptojanin1,2 | Bipolar disorder, Down syndrome, neuronal disorder. | [29,85,135] |
| PI(4,5) P2 | INPP4 | Asthma, nondegeneracy. | [23,130] | |
| PI(4,5) P2 | Actin reorganization | Human immunodeficiency virus-1 (HIV-1). | [136,137] | |
| PI(4,5) P2 | impairment of synaptic function | Amyloid-β peptide oligomers | Alzheimer’s disease | [138] |
| PI(3,4) P2. PIP3 | Akt/PKB | Cell survival and growth, cancer. | [16,130,131] | |
| PI(4,5) P2 | PRL-3 | Melanoma, colon cancer. | [131,136,137] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mandal, K. Review of PIP2 in Cellular Signaling, Functions and Diseases. Int. J. Mol. Sci. 2020, 21, 8342. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218342
AMA Style
Mandal K. Review of PIP2 in Cellular Signaling, Functions and Diseases. International Journal of Molecular Sciences. 2020; 21(21):8342. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218342
Chicago/Turabian StyleMandal, Kalpana. 2020. "Review of PIP2 in Cellular Signaling, Functions and Diseases" International Journal of Molecular Sciences 21, no. 21: 8342. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218342
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.