Identification of a New QTL Region on Mouse Chromosome 1 Responsible for Male Hypofertility: Phenotype Characterization and Candidate Genes

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
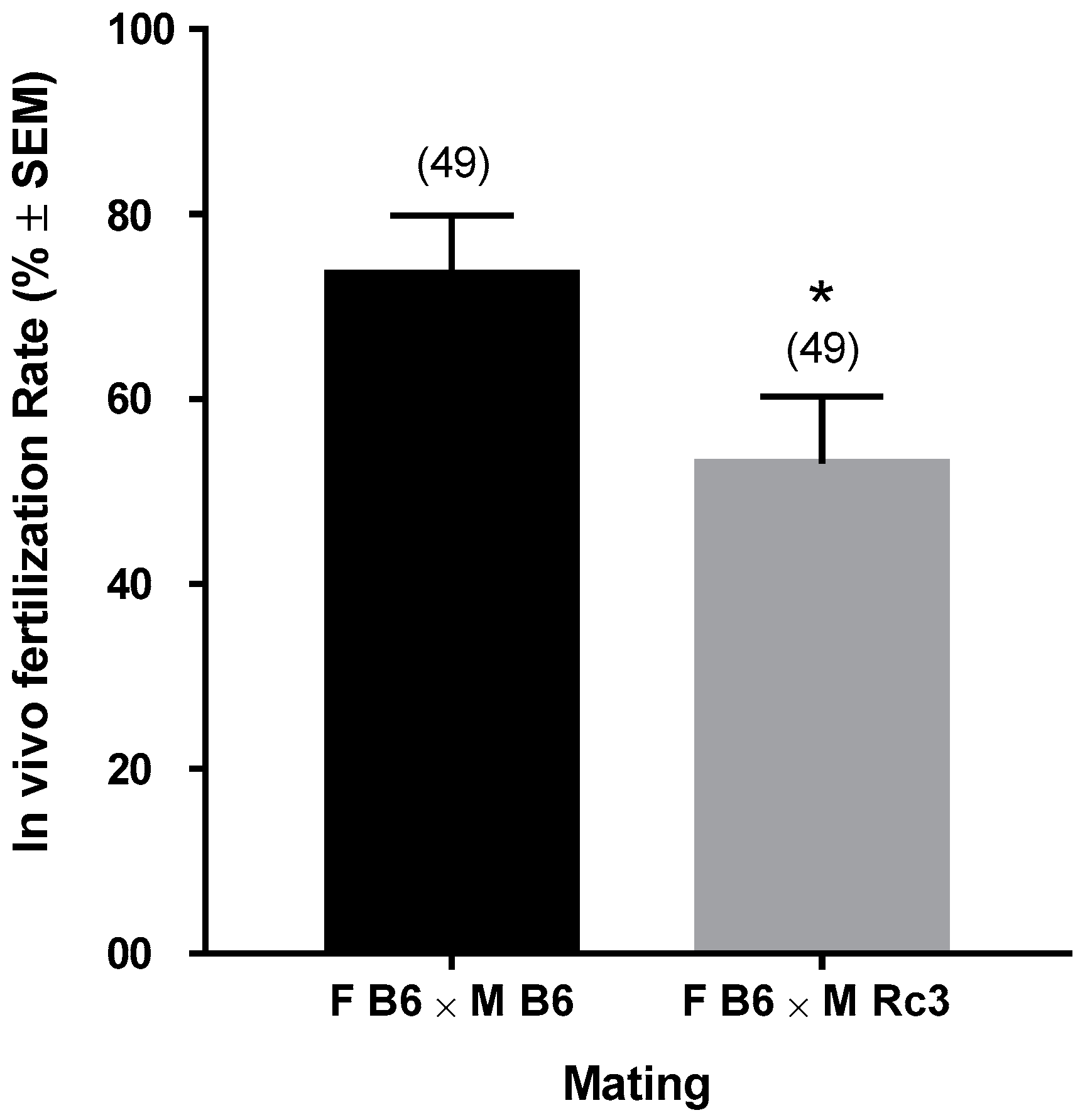
2.1. In Vivo Phenotyping
2.2. Phenotype Characterization
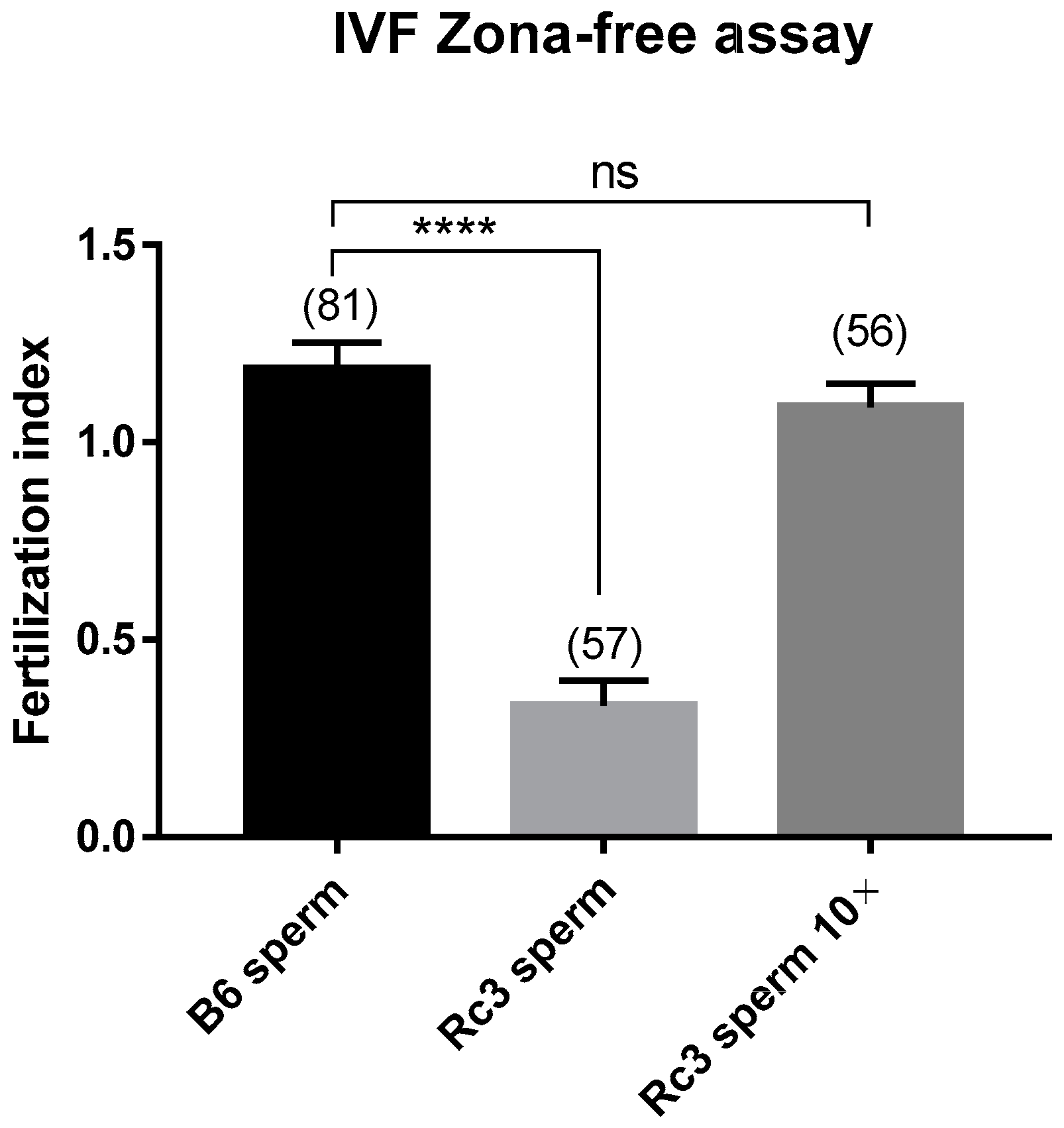
2.3. Characterization of the Sperm Defect of the Rc3 Males
2.4. QTL Fine Mapping
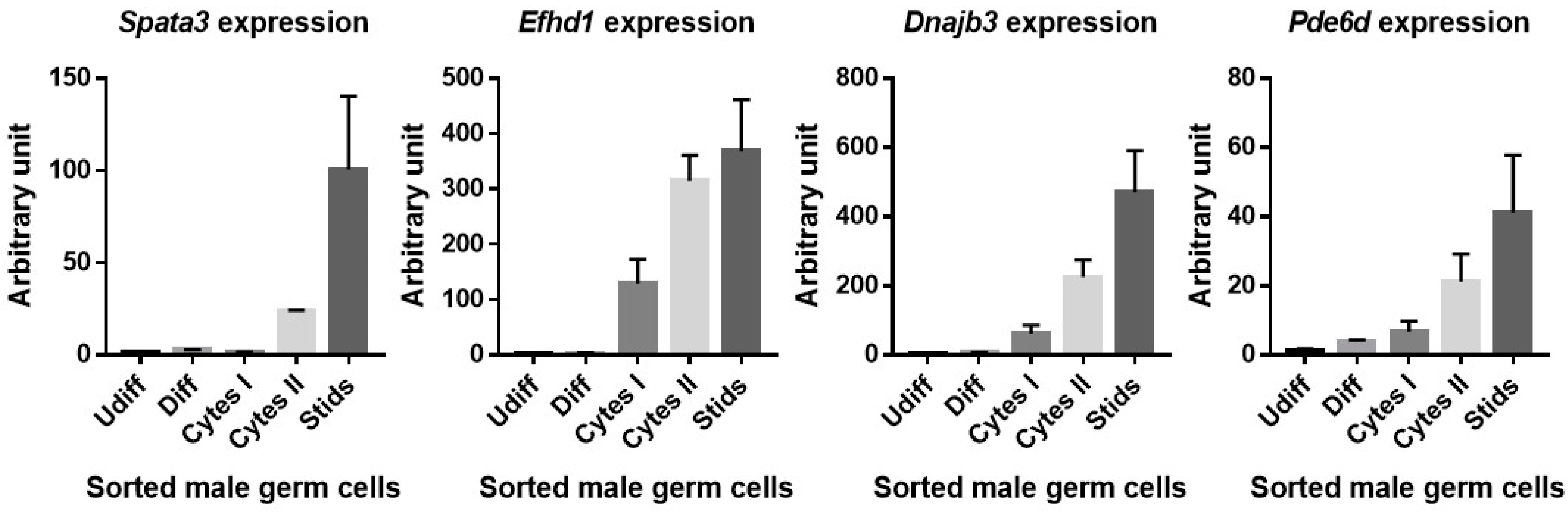
2.5. Identification of Candidate Genes in the QTL Region Responsible for Rc3 Sperm Phenotype
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals
4.3. Phenotyping by High Frequency Ultrasonography Evaluation of in Vivo Fertilization
4.4. Evaluation of in Vivo Fertilization
4.5. In Vitro Fertilization Assays
4.6. Assessment of Sperm Acrosome Reaction
4.7. Transmission Electron Microscopy Analysis of Sperm Cells
4.8. Digital and Bibliographic Tools Used to Shorten the List of Candidate Genes
4.9. Suspension Cell Sorting
4.10. RNA Extraction and Quantitative RT-PCR
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DAPI | 4′,6-diamidino-2-phenylindole |
| sAR | Spontaneous acrosome reaction |
| AR | Acrosome reaction |
| QTL | Quantitative trait locus |
| IRCS | Recombinant interspecific congenic strain |
| IVF | In vitro fertilization |
| KO | Knockout |
| Mafq1 | Male fertility QTL chromosome 1 |
References
- Matzuk, M.M.; Lamb, D.J. The biology of infertility: Research advances and clinical challenges. Nat. Med. 2008, 14, 1197–1213. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Cooke, H.J.; Saunders, P.T. Mouse models of male infertility. Nat. Rev. Genet. 2002, 3, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Borg, C.L.; Wolski, K.M.; Gibbs, G.M.; O’Bryan, M.K. Phenotyping male infertility in the mouse: How to get the most out of a “non-performer”. Hum. Reprod. Update 2010, 16, 205–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgio, G.; Szatanik, M.; Guenet, J.L.; Arnau, M.R.; Panthier, J.J.; Montagutelli, X. Interspecific recombinant congenic strains between C57BL/6 and mice of the Mus spretus species: A powerful tool to dissect genetic control of complex traits. Genetics 2007, 177, 2321–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- L’Hote, D.; Laissue, P.; Serres, C.; Montagutelli, X.; Veitia, R.A.; Vaiman, D. Interspecific resources: A major tool for quantitative trait locus cloning and speciation research. Bioessays 2010, 32, 132–142. [Google Scholar] [CrossRef]
- L’Hote, D.; Serres, C.; Veitia, R.A.; Montagutelli, X.; Oulmouden, A.; Vaiman, D. Gene expression regulation in the context of mouse interspecific mosaic genomes. Genome Biol. 2008, 9, R133. [Google Scholar] [CrossRef]
- L’Hote, D.; Vatin, M.; Auer, J.; Castille, J.; Passet, B.; Montagutelli, X.; Serres, C.; Vaiman, D. Fidgetin-like1 is a strong candidate for a dynamic impairment of male meiosis leading to reduced testis weight in mice. PLoS ONE 2011, 6, e27582. [Google Scholar] [CrossRef]
- Vatin, M.; Burgio, G.; Renault, G.; Laissue, P.; Firlej, V.; Mondon, F.; Montagutelli, X.; Vaiman, D.; Serres, C.; Ziyyat, A. Refined mapping of a quantitative trait locus on chromosome 1 responsible for mouse embryonic death. PLoS ONE 2012, 7, e43356. [Google Scholar] [CrossRef]
- Laissue, P.; Lakhal, B.; Vatin, M.; Batista, F.; Burgio, G.; Mercier, E.; Santos, E.D.; Buffat, C.; Sierra-Diaz, D.C.; Renault, G.; et al. Association of FOXD1 variants with adverse pregnancy outcomes in mice and humans. Open Biol. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Vatin, M.; Bouvier, S.; Bellazi, L.; Montagutelli, X.; Laissue, P.; Ziyyat, A.; Serres, C.; De Mazancourt, P.; Dieudonné, M.N.; Mornet, E.; et al. Polymorphisms of human placental alkaline phosphatase are associated with in vitro fertilization success and recurrent pregnancy loss. Am. J. Pathol. 2014, 184, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Vatin, M.; Bouvier, S.; Bellazzi, L.; Laissue, P.; Burguio, G.; Montagutelli, X.; Serres, C.; Ziyyat, A.; Mornet, E.; Gris, J.C.; et al. Pregnancy failure and alkaline phosphatases: From mouse genetics to human disease. Bull. Group Int. Rech. Sci. Stomatol. Odontol. 2012, 51, e30. [Google Scholar]
- Yuan, J.; Chen, J. FIGNL1-containing protein complex is required for efficient homologous recombination repair. Proc. Natl. Acad. Sci. USA 2013, 110, 10640–10645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Ko, D.S.; Lee, H.C.; Lee, H.J.; Park, W.I.; Kim, S.S.; Park, J.K.; Yang, B.C.; Park, S.B.; Chang, W.K.; et al. Comparison of maturation, fertilization, development, and gene expression of mouse oocytes grown in vitro and in vivo. J. Assist. Reprod. Genet. 2004, 21, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbaux, S.; Ialy-Radio, C.; Chalbi, M.; Dybal, E.; Homps-Legrand, M.; Do Cruzeiro, M.; Vaiman, D.; Wolf, J.P.; Ziyyat, A. Sperm SPACA6 protein is required for mammalian Sperm-Egg Adhesion/Fusion. Sci. Rep. 2020, 10, 5335. [Google Scholar] [CrossRef] [Green Version]
- Valdivia, M.; Sillerico, T.; De Ioannes, A.; Barros, C. Proteolytic activity of rabbit perivitelline spermatozoa. Zygote 1999, 7, 143–149. [Google Scholar] [CrossRef]
- Clift, L.E.; Andrlikova, P.; Frolikova, M.; Stopka, P.; Bryja, J.; Flanagan, B.F.; Johnson, P.M.; Dvorakova-Hortova, K. Absence of spermatozoal CD46 protein expression and associated rapid acrosome reaction rate in striped field mice (Apodemus agrarius). Reprod. Biol. Endocrinol. 2009, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Sebkova, N.; Ded, L.; Vesela, K.; Dvorakova-Hortova, K. Progress of sperm IZUMO1 relocation during spontaneous acrosome reaction. Reproduction 2014, 147, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Wiser, A.; Sachar, S.; Ghetler, Y.; Shulman, A.; Breitbart, H. Assessment of sperm hyperactivated motility and acrosome reaction can discriminate the use of spermatozoa for conventional in vitro fertilisation or intracytoplasmic sperm injection: Preliminary results. Andrologia 2014, 46, 313–315. [Google Scholar] [CrossRef]
- Efrat, M.; Stein, A.; Pinkas, H.; Breitbart, H.; Unger, R.; Birk, R. Paraoxonase 1 (PON1) attenuates sperm hyperactivity and spontaneous acrosome reaction. Andrology 2019, 7, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Inoue, N.; Ikawa, M.; Nakanishi, T.; Matsumoto, M.; Nomura, M.; Seya, T.; Okabe, M. Disruption of mouse CD46 causes an accelerated spontaneous acrosome reaction in sperm. Mol. Cell Biol. 2003, 23, 2614–2622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crapster, J.A.; Rack, P.G.; Hellmann, Z.J.; Le, A.D.; Adams, C.M.; Leib, R.D.; Elias, J.E.; Perrino, J.; Behr, B.; Li, Y.; et al. HIPK4 is essential for murine spermiogenesis. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Song, Z.; Wang, L.; Yu, H.; Liu, W.; Shang, Y.; Xu, Z.; Zhao, H.; Gao, F.; Wen, J.; et al. Sirt1 regulates acrosome biogenesis by modulating autophagic flux during spermiogenesis in mice. Development 2017, 144, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harbuz, R.; Zouari, R.; Pierre, V.; Ben Khelifa, M.; Kharouf, M.; Coutton, C.; Merdassi, G.; Abada, F.; Escoffier, J.; Nikas, Y.; et al. A recurrent deletion of DPY19L2 causes infertility in man by blocking sperm head elongation and acrosome formation. Am. J. Hum. Genet. 2011, 88, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Pierre, V.; Martinez, G.; Coutton, C.; Delaroche, J.; Yassine, S.; Novella, C.; Pernet-Gallay, K.; Hennebicq, S.; Ray, P.F.; Arnoult, C. Absence of Dpy19l2, a new inner nuclear membrane protein, causes globozoospermia in mice by preventing the anchoring of the acrosome to the nucleus. Development 2012, 139, 2955–2965. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.J.; Ying, L.; Ghosh, S.; de Franca, L.R.; Russell, L.D. Sertoli cell cycle: A re-examination of the structural changes during the cycle of the seminiferous epithelium of the rat. Anat. Rec. 1993, 237, 187–198. [Google Scholar] [CrossRef]
- Berruti, G.; Perego, L.; Martegani, E. Molecular cloning and developmental pattern of expression of MSJ-1, a new male germ cell-specific DNAJ homologue. Adv. Exp. Med. Biol. 1998, 444, 145–150. [Google Scholar] [CrossRef]
- Meccariello, R.; Cobellis, G.; Berruti, G.; Junier, M.P.; Ceriani, M.; Boilee, S.; Pierantoni, R.; Fasano, S. Mouse sperm cell-specific DnaJ first homologue: An evolutionarily conserved protein for spermiogenesis. Biol. Reprod. 2002, 66, 1328–1335. [Google Scholar] [CrossRef] [Green Version]
- Meccariello, R.; Berruti, G.; Chianese, R.; De Santis, R.; Di Cunto, F.; Scarpa, D.; Cobellis, G.; Zucchetti, I.; Pierantoni, R.; Altruda, F.; et al. Structure of msj-1 gene in mice and humans: A possible role in the regulation of male reproduction. Gen. Comp. Endocrinol. 2008, 156, 91–103. [Google Scholar] [CrossRef]
- Abu-Farha, M.; Cherian, P.; Al-Khairi, I.; Tiss, A.; Khadir, A.; Kavalakatt, S.; Warsame, S.; Dehbi, M.; Behbehani, K.; Abubaker, J. DNAJB3/HSP-40 cochaperone improves insulin signaling and enhances glucose uptake in vitro through JNK repression. Sci. Rep. 2015, 5, 14448. [Google Scholar] [CrossRef] [Green Version]
- Boillee, S.; Berruti, G.; Meccariello, R.; Grannec, G.; Razan, F.; Pierantoni, R.; Fasano, S.; Junier, M.P. Early defect in the expression of mouse sperm DNAJ 1, a member of the DNAJ/heat shock protein 40 chaperone protein family, in the spinal cord of the wobbler mouse, a murine model of motoneuronal degeneration. Neuroscience 2002, 113, 825–835. [Google Scholar] [CrossRef]
- Gaasch, J.A.; Bolwahnn, A.B.; Lindsey, J.S. Hepatocyte growth factor-regulated genes in differentiated RAW 264.7 osteoclast and undifferentiated cells. Gene 2006, 369, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Mitsugi, R.; Itoh, T.; Fujiwara, R. Expression of Human DNAJ (Heat Shock Protein-40) B3 in Humanized UDP-glucuronosyltransferase 1 Mice. Int. J. Mol. Sci. 2015, 16, 14997–15008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berruti, G.; Martegani, E. The deubiquitinating enzyme mUBPy interacts with the sperm-specific molecular chaperone MSJ-1: The relation with the proteasome, acrosome, and centrosome in mouse male germ cells. Biol. Reprod. 2005, 72, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arredouani, A.; Diane, A.; Khattab, N.; Bensmail, I.; Aoude, I.; Chikri, M.; Mohammad, R.; Abou-Samra, A.B.; Dehbi, M. DNAJB3 attenuates metabolic stress and promotes glucose uptake by eliciting Glut4 translocation. Sci. Rep. 2019, 9, 4772. [Google Scholar] [CrossRef] [Green Version]
- Van De Weghe, J.C.; Rusterholz, T.D.S.; Latour, B.; Grout, M.E.; Aldinger, K.A.; Shaheen, R.; Dempsey, J.C.; Maddirevula, S.; Cheng, Y.H.; Phelps, I.G.; et al. Mutations in ARMC9, which Encodes a Basal Body Protein, Cause Joubert Syndrome in Humans and Ciliopathy Phenotypes in Zebrafish. Am. J. Hum. Genet. 2017, 101, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Brownlow, S.; Webster, R.; Croxen, R.; Brydson, M.; Neville, B.; Lin, J.P.; Vincent, A.; Newsom-Davis, J.; Beeson, D. Acetylcholine receptor delta subunit mutations underlie a fast-channel myasthenic syndrome and arthrogryposis multiplex congenita. J. Clin. Invest. 2001, 108, 125–130. [Google Scholar] [CrossRef]
- Shen, X.M.; Fukuda, T.; Ohno, K.; Sine, S.M.; Engel, A.G. Congenital myasthenia-related AChR delta subunit mutation interferes with intersubunit communication essential for channel gating. J. Clin. Invest. 2008, 118, 1867–1876. [Google Scholar] [CrossRef]
- Shen, X.M.; Ohno, K.; Fukudome, T.; Tsujino, A.; Brengman, J.M.; De Vivo, D.C.; Packer, R.J.; Engel, A.G. Congenital myasthenic syndrome caused by low-expressor fast-channel AChR delta subunit mutation. Neurology 2002, 59, 1881–1888. [Google Scholar] [CrossRef]
- Lassalle, B.; Bastos, H.; Louis, J.P.; Riou, L.; Testart, J.; Dutrillaux, B.; Fouchet, P.; Allemand, I. ‘Side Population’ cells in adult mouse testis express Bcrp1 gene and are enriched in spermatogonia and germinal stem cells. SO Dev. 2004, 131, 479–487. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vatin, M.; Girault, M.-S.; Firlej, V.; Marchiol, C.; Ialy-Radio, C.; Montagutelli, X.; Vaiman, D.; Barbaux, S.; Ziyyat, A. Identification of a New QTL Region on Mouse Chromosome 1 Responsible for Male Hypofertility: Phenotype Characterization and Candidate Genes. Int. J. Mol. Sci. 2020, 21, 8506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228506
Vatin M, Girault M-S, Firlej V, Marchiol C, Ialy-Radio C, Montagutelli X, Vaiman D, Barbaux S, Ziyyat A. Identification of a New QTL Region on Mouse Chromosome 1 Responsible for Male Hypofertility: Phenotype Characterization and Candidate Genes. International Journal of Molecular Sciences. 2020; 21(22):8506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228506
Chicago/Turabian StyleVatin, Magalie, Marie-Sophie Girault, Virginie Firlej, Carmen Marchiol, Côme Ialy-Radio, Xavier Montagutelli, Daniel Vaiman, Sandrine Barbaux, and Ahmed Ziyyat. 2020. "Identification of a New QTL Region on Mouse Chromosome 1 Responsible for Male Hypofertility: Phenotype Characterization and Candidate Genes" International Journal of Molecular Sciences 21, no. 22: 8506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228506