TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation



Koltzov Institute of Developmental Biology, Russian Academy of Sciences, Vavilova str. 26, 119991 Moscow, Russia
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(22), 8871; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228871
Submission received: 30 September 2020
/
Revised: 17 November 2020
/
Accepted: 18 November 2020
/
Published: 23 November 2020
(This article belongs to the Special Issue Structural Variability and Flexibility of the Genome)
Abstract
:Robertsonian (centric-fusion) translocation is the form of chromosomal translocation in which two long arms of acrocentric chromosomes are fused to form one metacentric. These translocations reduce the number of chromosomes while preserving existing genes and are considered to contribute to speciation. We asked whether hypomorphic mutations in genes that disrupt the formation of pericentromeric regions could lead to centric fusion. TBP-related factor 2 (Trf2) encodes an alternative general transcription factor. A decrease of TRF2 expression disrupts the structure of the pericentromeric regions and prevents their association into chromocenter. We revealed several centric fusions in two lines of Drosophila melanogaster with weak Trf2 alleles in genetic experiments. We performed an RNAi-mediated knock-down of Trf2 in Drosophila and S2 cells and demonstrated that Trf2 upregulates expression of D1—one of the major genes responsible for chromocenter formation and nuclear integrity in Drosophila. Our data, for the first time, indicate that Trf2 may be involved in transcription program responsible for structuring of pericentromeric regions and may contribute to new karyotypes formation in particular by promoting centric fusion. Insight into the molecular mechanisms of Trf2 function and its new targets in different tissues will contribute to our understanding of its phenomenon.
1. Introduction
Centric-fusion translocation (a Robertsonian translocation) is a type of chromosome rearrangement, in which two long arms of acrocentric chromosomes fuse to form one metacentric [1]. Centric chromosome fusion often accompanies pathological genetically inherited human syndromes (trisomy 13 (Patau) syndrome and trisomy 21 (Down) syndrome), so it is important to study the source of their occurrence. Such chromosomal rearrangements, like others, can be caused by environmental risk factors (for example, ionizing radiation or chemical carcinogens), but they also may be caused by mutations that destabilize the compact structure of chromatin in pericentromeric regions [2]. It has been suggested that centric fusion may cause changes in the number of chromosomes in the karyotype of species, and thus play a role in speciation and influence the course of evolution in general [3,4,5,6,7,8,9,10]. This is possible due to the unique feature of such rearrangements not to disrupt the total number of genes (which can be fatal for the organism), but to change the number of chromosomes, contributing to the genetic isolation of the population—a condition necessary for new species to evolve [11,12]. In this regard, the search for genes whose mutations may be able to influence the course of the evolutionary process through a change in the chromosome set is of fundamental biological importance. Mutations of such genes should create favorable conditions for the emergence of centric fusion, in other words, they should affect the structure of heterochromatin in pericentromeric regions. Obviously, such mutants will have an increased rate of chromosome nondisjunction compared to karyotypically normal animals. Indeed, the proper segregation of sister chromatids between daughter cells requires coordinated interaction of centrosomes, centromeres, kinetochores, spindle fibrils, topoisomerases, proteolytic processes, and motor proteins [13,14]. A number of genes control each steps of this process. Mutations in these genes lead to chromosome nondisjunction and tumorigenesis [15]. However, it is unlikely that they will contribute to centric fusion. On the other hand, chromosomes must be “prepared” (or structurally organized) when they enter meiosis (or mitosis). Structural disorganization of chromosomes or their regions that control the correct pairing of homologues during meiosis often results in the incorrect chromosome segregation. We believe that it is more likely that mutations in genes that disrupt the organization of pericentromeric regions will promote centric fusion of chromosomes.
One way of eukaryotic genome organization is the formation of a chromocenter, which is evolutionally involved in the regulation of chromosome behavior in dividing cells not only among insects but also among plants, mammals, mollusks, and even yeast [16,17,18,19,20,21]. This nuclear structure arises in differentiated somatic and germ cells during interphase and meiotic prophase and is generated by the association of pericentromeric regions of all or separate groups of chromosomes. Studies on Drosophila and mice have clearly demonstrated that its disorganization leads to genomic disbalance. It has been suggested that the chromocenter performs coordination of chromosome before division and indirectly increases the frequency of crossing over when its forming is delayed [22,23,24]. A detailed study of chromocenters in Drosophila oocytes has shown that their structure is genetically programmed [25,26].
In screening for genes that control the formation and reorganization of chromocenter, the high frequency of chromosome nondisjunction is the main peculiarity in the progeny of mutant parents. Moreover, recent studies on Drosophila and mouse cells have shown that satellite DNA in pericentromeric regions plays a critical role in maintaining genome integrity in the cell nucleus since depletions of two sequence-specific satellite DNA-binding proteins, D1 and Prod, disrupts chromocenters and leads to the formation of micronuclei [7,27]. Nevertheless, it is still not known whether mutations that disrupt chromocenter formation and, in particular, pericentromeric heterochromatin, can cause centric fusion.
In Drosophila, TBP-related factor 2 (Trf2) encodes an alternative general transcription factor that is homologous to vertebrate Trf2 protein and belongs to a conservative TATA box-binding protein (Tbp) gene family [28]. The ubiquitous presence of Trf2 allows studying its functions in Drosophila. Previously discovered lawcp1 (leg-arista-wing complex) mutation appeared to be the only viable mutation that decreases Trf2 expression in D. melanogaster [29,30]. It was demonstrated that lawcp1 suppresses the phenotype of flies with mutations in genes that encode Polycomb Group proteins, which are negative epigenetic regulators of transcription via chromatin modification [31]. At the same time, it was found that lawcp1 increase the effect of transvection (or allelic complementation [32]) caused by disruptions of the homologous chromosome pairing and suppress the position effect variegation. A decrease in Trf2 gene expression in mice and Drosophila could result in the disruption of chromatin condensation [33,34]. Genetic experiments on Drosophila with the use of mass crosses demonstrated a high frequency of chromosome nondisjunction in Trf2 mutant female meiosis and this anomaly accompanied with an abnormal chromatin packaging and particularly with the failure of chromocenter organization [35]. Taking into account the specific Trf2 mutant phenotype associated with disruption of pericentromeric heterochromatin, we hypothesized that even a slight decrease in Trf2 gene activity could cause a chromosome centric fusion.
To test our hypothesis, new genetic screening of flies with X chromosome nondisjunction and attached-X chromosomes in lines with Trf2 lethal mutations were performed. We revealed several flies carrying attached X in two lines with weak Trf2 alleles. We discovered that these events more often occurred in meiosis of females with lower levels of nondisjunction. We also demonstrated that Trf2 upregulates expression of D1—one of two major genes responsible for chromocenter formation and nuclear integrity in Drosophila. Our data, for the first time, indicate that Trf2 may be involved in a transcription program responsible for structuring of pericentromeric regions and contributing to new karyotypes formation in particular by promoting centric fusion.
2. Results
2.1. Frequency of X Chromosome Nondisjunction and Existence of Attached-X Chromosomes in Females with Lethal Trf2 Mutations
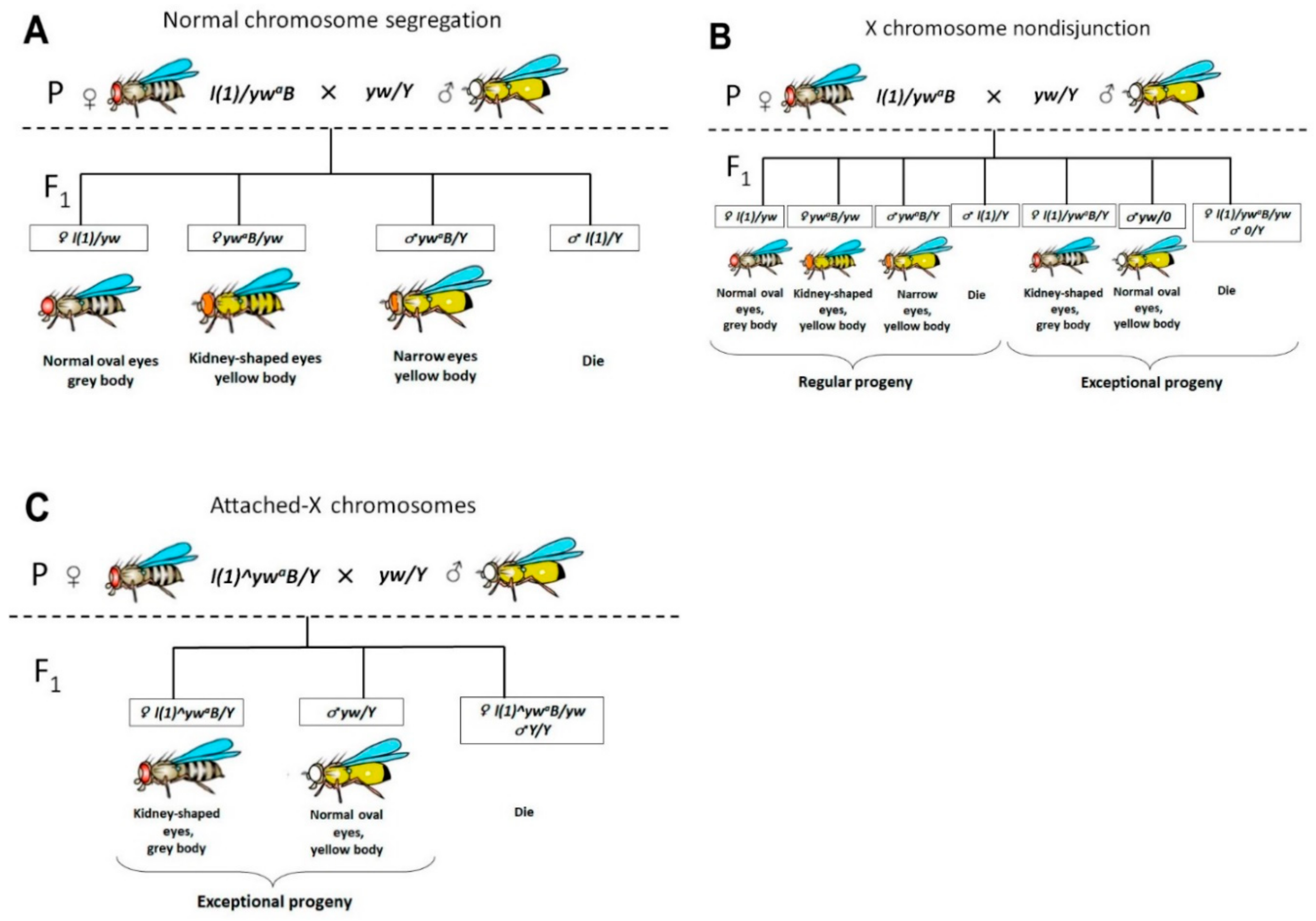
Earlier, in genetic experiments on D. melanogaster using mass crosses, we demonstrated a high frequency of X chromosome nondisjunction in females with Trf2 lethal mutants [35]. However, mass crosses do not allow identifying females with attached-X chromosomes. To provide this we performed individual crosses of about 30 females from seven lines that contain Trf2 lethal mutations. Further, phenotypic analysis of F1 progeny was carried out to determine their belonging to normal or exceptional classes.
In the case of normal chromosome segregation in female meiosis, only regular classes will be present in F1 generation (Figure 1A). In the case of X chromosome nondisjunction in females during meiosis, splitting of traits will be observed in their F1 generation, and both regular and exceptional classes will be present (Figure 1B). Attached-X chromosomes consist of two full-length X chromosomes sharing a common centromere so that they are always inherited together. In crosses of normal males to females carrying attached-X chromosomes, male progeny will inherit their X from their father and their Y from their mother (X^X/Y). It means that the only exceptional flies will be found in the offspring if the parent female carries attached X (Figure 1C). By following this principle, we could determine if the parent female had chromosome nondisjunction or if the female was carrying attached-X chromosomes.
Phenotypic analysis revealed that in seven lines tested, two, (l(1)G0425 and l(1)G0356), exhibited the presence of females with attached X (Table 1 and Table S1). Their male progeny had the expected patroclinous phenotype (“yellow body”).
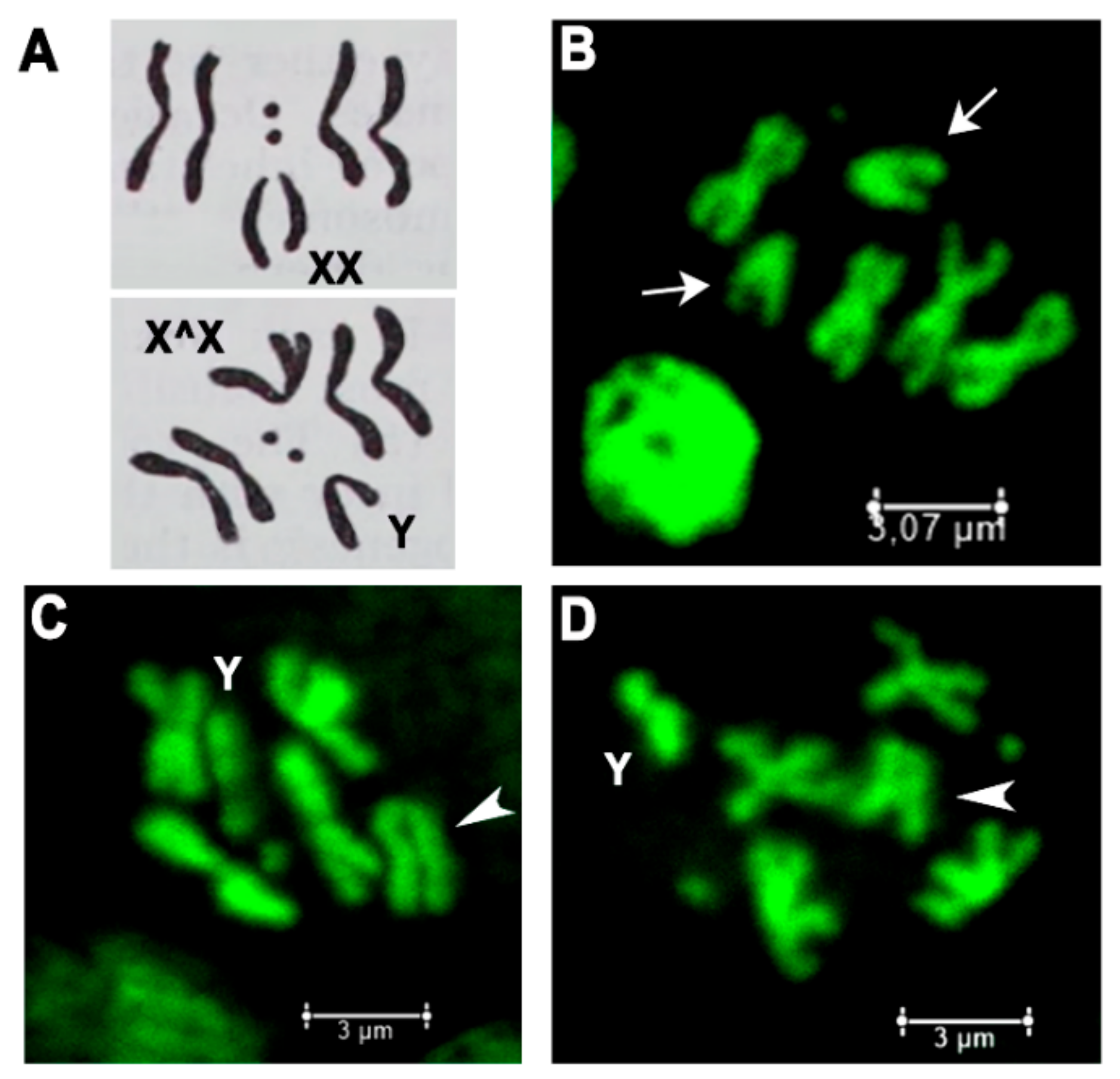
To confirm the presence of attached-X chromosomes (X^X/Y) in discovered l(1)G0425 and l(1)G0356 females, they were bred in individual crosses with yw/Y males and metaphase chromosome plates of larval neuroblasts were prepared (Figure 2). The mitotic karyotype of D. melanogaster females comprises four pairs of chromosomes (2n = 8), three autosomal pairs plus the pair of X chromosomes (Figure 2A). Two pairs of metacentric autosomes II and III form four cruciform-like structures. One pair of small acrocentric autosomes IV forms two dot-like structures and one pair of large submetacentric X chromosomes forms two V-like structures (Figure 2B). Metaphase figures of l(1)G0425 and l(1)G0356 females with attached-X do not show V-like structures, but demonstrate five cruciform-like structures (two pairs of autosomes II, III and one X^X) plus J-shaped Y-chromosome (Figure 2C,D).
We cannot strictly assert that there were no such females in other five lines, since the selection of females for analysis was carried out randomly. Perhaps, if we increase the sample of analyzed females, we will be able to find such unique individuals. Nevertheless, our experiments demonstrate that a decrease in Trf2 gene expression may indeed lead to such a phenomenon as chromosome centric fusion.
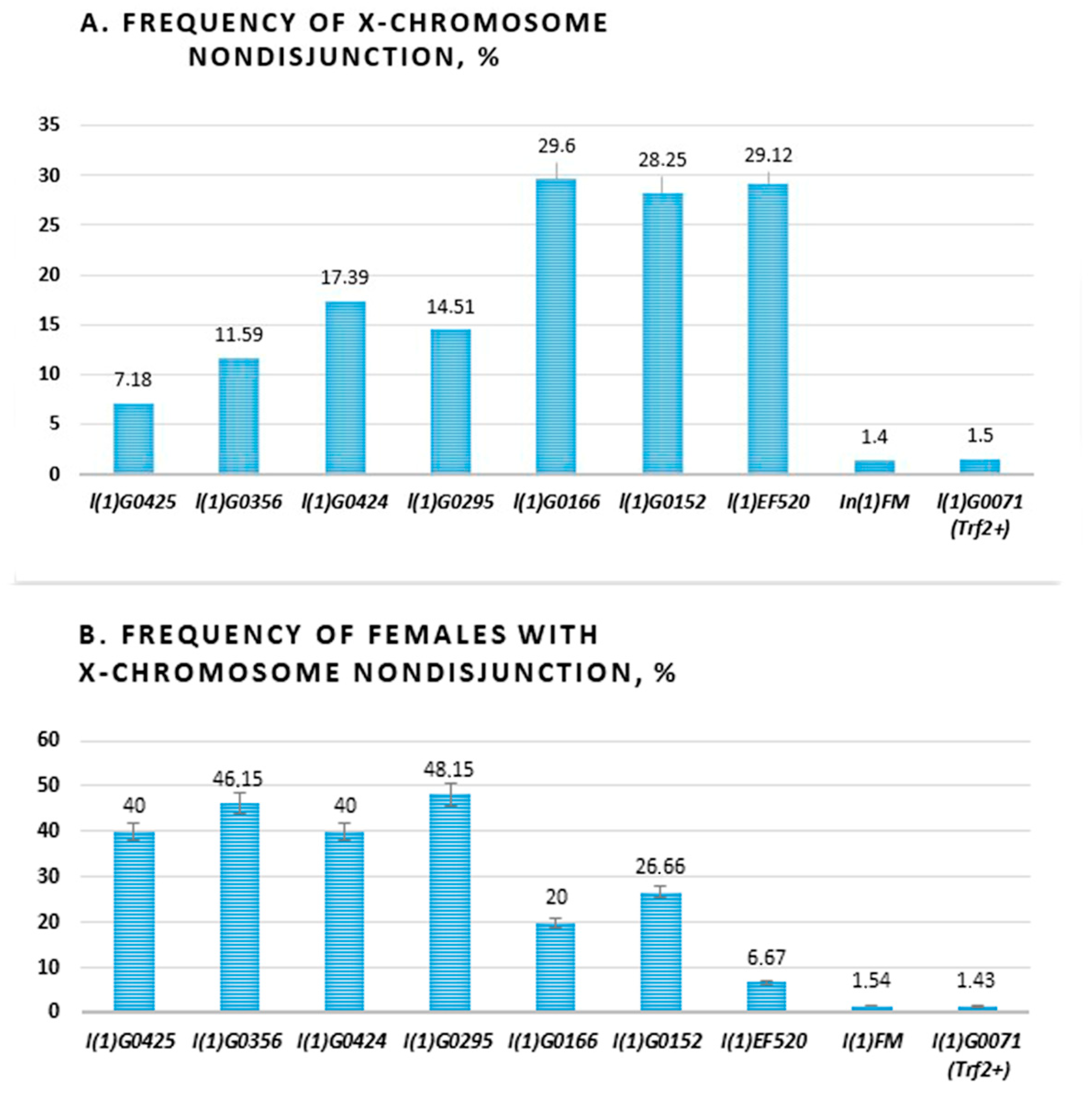
Then, we carried out a total count of females and males for each phenotypic class and calculated the frequency of X chromosome nondisjunction. A range of strengths was seen among the various alleles. The maximal levels of X chromosome nondisjunction were in three Trf2 alleles (l(1)G0166, l(1)G0152, l(1)EF520) which exceeds the frequency of nondisjunction in controls by approximately 20 times. In total, Four alleles (l(1)G0425, l(1)G0356, l(1)G0424, l(1)G0295) were weak, giving levels of 7.18%, 11.59% 17.39%, and 14,51%, respectively (Table 1, Table S1, Figure 3A).
We also calculated the frequency of parent females, which produced exceptional progeny in tested lines (Figure 3B). It turned out that in four of seven lines (l(1)G0425, l(1)G0356, (1)G0424, l(1)G0295) about half (40–48.15%) of females have X chromosome nondisjunction. It is interesting that these were lines with flies bearing weaker alleles. In two lines (l(1)G0166, l(1)G0152) only a quarter (20% and 26.66%) of females have X chromosome nondisjunction and in the l(1)EF520 line, 6.67% of females produced exceptional progeny. The last three alleles demonstrated high levels of X chromosome nondisjunction. Thus, we found a negative correlation between the level of chromosome nondisjunction and the number of females producing exclusive offspring in different lines with Trf2 lethal mutation. Moreover, we have demonstrated that decrease of Trf2 gene expression may provide chromosome centric fusion.
2.2. Decrease of Trf2 Gene Expression Downregulates D1 Chromosomal Protein
Earlier, we demonstrated that the depletion of Trf2 leads to a failure of chromocenter formation and chromatin condensation [35]. On the other hand, chromocenter splitting assumes the disruption of interchromosomal ectopic contacts in the pericentromeric heterochromatin region. D1 chromosomal protein (D1) encodes a multi-AT-hook chromosomal protein that associates with AT-rich satellites, including the SAT-III repeats of the X chromosome. The depletion of D1 leads to chromocenter disruption and micronuclei formation [7]. As Trf2 encodes the general transcription factor, we suggested that it could drive D1 gene expression.
It was previously demonstrated that Trf2-containing complexes selectively recognize TATA-less promoters and initiate not TATA-, but DPE- or TCT-depending transcriptions [36]. To evaluate the potential role of Trf2 in the regulation of D1 gene transcription we analyzed D1 promoter regions using bioinformatics approach as described in Materials and Methods. According to Flybase release FB2020_04, the Drosophila D1 gene encodes three transcripts and has two alternative promoters. We revealed that there are several initiators (Inr) and Bridges (a bipartite core promoter element BridgeI and BridgeII) within predicted promoters in 5′-flanking regulatory regions of D1 transcripts and only one TATA-box and TFIIB Recognition element (BRE) at the 3′-end -termini of D1-RC splice variant of the first exon (Figure 4 and Figure S1). It indicates that Trf2 could be a good candidate to initiate D1 transcription.
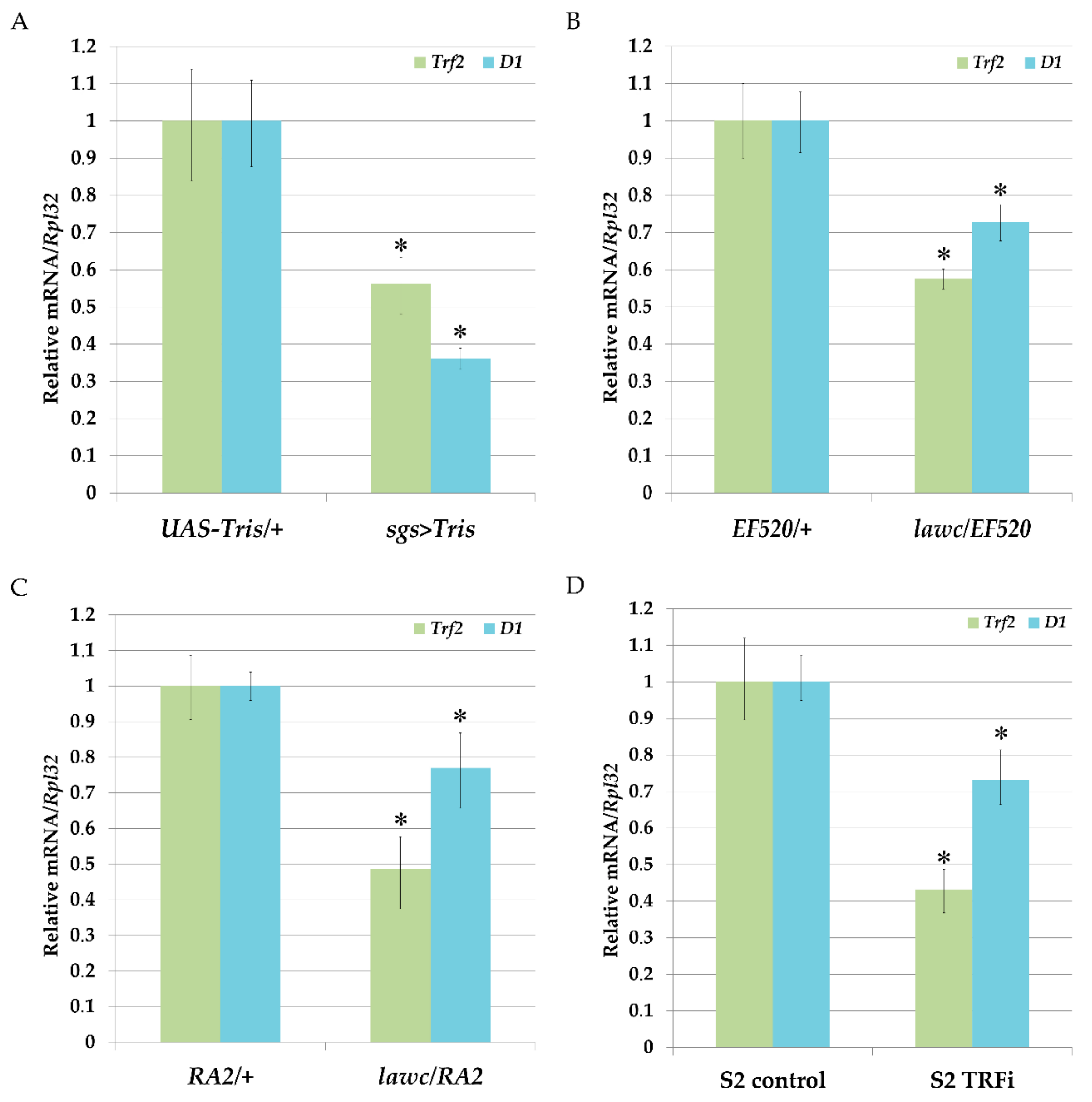
To confirm the ability of Trf2 to affect D1 gene expression in Drosophila we carried out RNAi-mediated knock-down of Trf2 using DNA-directed RNA interference (ddRNAi) technique and quantified the levels of D1 mRNA before and after Trf2 knock-down. For this, we designed UAS-Tris flies with inducible expression of double-stranded RNA homologous to the Trf2 mRNA. To drive UAS-Tris transgene in salivary glands we used sgs-Gal4 strain. The comparison of the levels of mRNA synthesis by qRT-PCR in salivary glands of UAS-Tris/+ larvae with UAS-Tris/+; sgs-Gal4/+ larvae demonstrated a decrease in D1 gene expression confirming that Trf2 may control D1 expression (Figure 5A). Similar results were obtained when we examine lawcp1/l(1)EF520 and lawcp1/Df(1)RA2 mutant flies with decreased expression of Trf2 (Figure 5B,C). Mutants with downregulated Trf2 expression had a drop in D1 gene expression. Next, we performed RNAi-mediated silencing of Trf2 in Drosophila S2 cells as described in Materials and Methods and confirmed our results (Figure 5D).
In the next experiments, we performed RNAi-mediated silencing of Trf2 and examined Drosophila S2 cells with depleted TRF2. We revealed an increased frequency of the micronuclei formation and the defective reformation of the nuclear envelopes (Figure 6). These defects were similar to those described for D1 mutant cells [7].
Together our results suggest that mutant Trf2 phenotype associated with genomic disbalance could be mediated in particular by decreased D1 gene expression.
3. Discussion
Here, in genetic experiments, we have demonstrated for the first time that the decreased expression of Trf2 could facilitate the linkage of two X chromosomes to the centromere. Attached X in Drosophila is a special case of the centric fusion (also referred to as Robertsonian translocation), which involves two homologous acrocentric chromosomes. We were looking for individuals with attached X because in D. melanogaster only two—a large X and fourth dot chromosome—from four chromosomes are acrocentric and it is more suitable to search for attached sex chromosomes bearing marker mutations. In other words, we have documented the formation of isochromosomes that carry identical copies of the same arm joined through a single centromere. We do not rule out centric fusion between X and the fourth chromosomes in our experiments, but we have not directly tested this. Nevertheless, we observed the formation of micronuclei in S2 cells with depleted Trf2, which may be a consequence of aneuploidy and lagging chromosome formation in mitosis against the background of translocations between non-homologous chromosomes [37].
We also found a high frequency of X chromosome nondisjunction in females with different Trf2 alleles. By the strength of nondisjunction, we have divided them into two groups. In lines with a relatively rare appearance of females with nondisjunction of X chromosome (up to 26%), the levels of nondisjunction were the highest, reaching 29% (the first group: l(1)G0166, l(1)G0152, l(1)EF520; Figure 3). Vice versa: in lines with frequent appearance of females with X chromosome nondisjunction (40–48.15%) the levels of nondisjunction were lower, barely reaching 17% (the second group: l(1)G0425, l(1)G0356, l(1)G0424, l(1)G0295; Figure 3). However, in all mutant Trf2 alleles, the frequency of X chromosome nondisjunction significantly exceeded the control one calculated for line with balancer FM7a chromosome, on which Trf2 lethals were maintained (1.4%), and for the line with wild type Trf2 allele but with the same (as in Trf2 mutants) genetic background (1.5%). The rarity of females with a high frequency of X chromosome nondisjunction (from the first group) can be explained by the fact that in these lines the effect of Trf2 mutation on chromosome destabilization is probably stronger than in others, and many gametes die during female meiosis, dropping out of the analysis. Lower levels of nondisjunction in the remaining Trf2 alleles allows for a larger number of females producing exceptional progeny to survive increasing the possibility of their detection. It is important to note that X-attachments occurred in the second group of alleles. We believe that only weak Trf2 alleles are able to create optimal “boundary” conditions of genome destabilization, which allow exceptional progeny to survive, including those with centric fusion.
Previously, it was shown that segregation errors observed in Trf2 mutant meiosis could be a consequence of split chromocenters and improper chromatin condensation not only in Drosophila, but also in mice [34,35]. Moreover, immunostaining of Drosophila polytene chromosomes revealed that Trf2 resides in pericentromeric regions [38]. However, despite such attractive localization it is unlikely that general transcription factor Trf2 directly participates in pericentromeric heterochromatin organization. Indeed, a recent study demonstrated that Trf2 interacts with TFIIA-L paralog Moonshine and can be recruited to the heterochromatin to initiate a heterochromatin-dependent transcription within PIWI-interacting RNA (piRNA) clusters [39]. This small RNA pathway acts in reproductive cells to prevent active transcription and transpositions of different mobile elements through heterochromatin formation at their location—mostly in pericentromeric region [40]. Thus, the localization of Trf2 in pericentromeric heterochromatin could be explained by its transcriptional activity within piRNA clusters in gonads. Nevertheless, the disorganized chromocenters observed in somatic cells mutant for Trf2 suggests its indirect participation in the maintenance of high order chromatin structure in pericentromeric region.
It was recently reported that the multi-AT-hook satellite DNA-binding protein, D1, is responsible for packaging of pericentromeric satellite DNA from heterologous chromosomes into chromocenters [7,27]. The bioinformatics analysis of D1 5′-flanking regulatory regions allows us to conclude that Trf2 is the most preferred candidate for D1 transcription activation. The reported observation that Trf2 is enriched at the D1 gene promoter region supports this possibility [36]. Knock down experiments performed in vivo in Drosophila and in S2 cell culture as far as analyses of Trf2 mutants have clearly demonstrated that the depletion of Trf2 reduces D1 RNA levels (Figure 5).
Finally, we revealed that RNAi-mediated silencing of Trf2 in Drosophila S2 cells leads to prominent defect in nuclear architecture—the formation of micronuclei. This defect may by a consequence of incorrect chromosome segregation induced by different chromosome rearrangements (mostly translocations) [41]. On the other hand, segregation errors lead to discoordination in the reformation of the nuclear envelope [42]. Indeed, we observed Lamin-negative nuclei in Trf2 depleted cells (Figure 6) which indicates a problem with nuclear envelope reformation in cells with decreased Trf2 expression. Cells with loss of D1 function demonstrate similar anomaly [7] confirming our assumption that Trf2 is the prevalent general transcription factor upregulating D1 gene expression.
Recent findings revealed the involvement of Trf2 in specialized transcription programs during development [43]. It was postulated that Trf2 facilitated the evolution of the third germ layer (mesoderm) and bilateria [44]. Our data, for the first time, indicate that Trf2 may also be involved in transcription program responsible for the structuring of pericentromeric regions and may trigger new karyotypes formation in particular by promoting centric fusion. Insight into the molecular mechanisms of Trf2 function and its new targets in different tissues will contribute to our understanding of its phenomenon.
4. Materials and Methods
4.1. Fly Stocks, Rearing Conditions and Genetic Crosses
Flies were kept at 25 °C on standard Drosophila wheat meal–yeast–sugar–agar medium. Individual crosses were performed in standard glass vials with 2 to 3 males and 1 female per vial.
The following stocks of D. melanogaster with lethal mutations in Trf2 were obtained from Bloomington Drosophila Stock Centre: l(1)G0425/FM7a; l(1)G0356/FM7a; l(1)G0424/FM7a; l(1)G0295/FM7a; l(1)G0166/FM7a; l(1)G0152/FM7a; and l(1)EF520/FM7a. These flies carry lethal Trf2 mutations induced by p{lacW} transposon [45]. l(1)EF520 produces truncated Trf2 protein (Simonova, personal communication).
X-linked Trf2 lethal mutations are balanced by In(1)FM. This chromosome carries a dominant Bar (B) marker mutation (narrow eyes) and recessive allele of yellow (y) gene (yellow body). We crossed y+l(1)/In(1)FM, yB female with males that carried the X chromosome marked by the y1 mutation (y1/Y) individually in order to identify exceptional progeny and estimate the frequency of X chromosome nondisjunction and possible presence of attached-X chromosomes in parent females. The regular progeny were In(1)FM, yB/Y males with narrow eyes and yellow body and 2 classes of females including (1) In(1)FM, yB/yB+ (yellow body, kidney-shaped eyes) and (2) y+l(1)/yB+ (grey body and normal oval eyes).
When X chromosome nondisjunction occurred, males and females of exceptional classes (that always differ by phenotypes) were detected. These were X/0 males with normal oval eyes and yellow bodies and XX/Y females with grey bodies and kidney-shaped eyes. Males of the normal class hemizygous for the X-linked lethal allele—l(1)/Y—die. Exceptional Y/0 males also die and XX/X super-females had low viability and died at an early age. Therefore, the frequency of X chromosome nondisjunction (Qn) was calculated according to the formula Qn = 100% × 2(X0 + XXY)/(XX + 2XY + 2X0 + 2XXY), where X0 and XXY are the number of flies of exceptional classes; XX and XY are the number of flies of normal classes [46]. The sum of exceptional classes in the numerator was multiplied by 2 in order to take into account lethal classes with the XX/X and Y/0 genotypes. The number of XY males in denominator was multiplied by 2 in order to take into account the class of lethal l(1)/Y males.
To calculate the frequency of parental females with nondisjunction (Qf), the following formula was used: Qf = 100% × Nn/N, where Nn is the number of females with X chromosome nondisjunction, N is the total number of analyzed females.
To detect the influence of In(1)FM balancer chromosome on the frequency of X chromosome nondisjunction and compare it with the frequency of nondisjunction Qn calculated for tested alleles a control experiment was performed. For this we crossed In(1)FM, B/In(1)FM, B females with y1/Y males. In(1)FM, B/y1 females of regular class must have kidney-shaped eyes in the F1 progeny caused by a combination of one copy of the Bar mutant allele with one copy of the wild-type allele of this gene. In(1)FM, B/Y males of regular class must have narrow eyes caused by the presence of one copy of the Bar mutant allele. Exceptional In(1)FM, B/In(1)FM, B/Y females must have narrow eyes caused by two copies of the Bar mutant allele, while y1/0 males must have normal oval eyes and yellow bodies.
To determine the influence of p{lacW} transposon on X chromosome nondisjunction in Trf2 alleles and to take into account the genetic background other control experiment was performed. We detected the frequency of sex chromosome nondisjunction in l(1)G0071 line with lethal mutation caused by the integration of the p{lacW} transposon not to Trf2 gene region.
If the parent female produced only exceptional progeny, it was concluded that she had attached X.
The lawcp1 mutation was described earlier [29]. Df(1)RA2/FM4, y31dsc8dmB (RA2) flies contain a deletion of [7D10;8A4-5] cytological region which overlaps Trf2 locus [7E]. l(1)EF520/lawcp1 and Df(1)RA2/lawcp1 heterozygous flies were used in RT-PCR experiments.
The sgs-GAL4 line was a gift from Dr. Vladik Mogila (Institute of Gene Biology Russian Academy of Science).
4.2. Drosophila Melanogaster Schneider 2 (S2) Cell Culture
Drosophila S2 cells (ATTC, Rockville, MD, USA) were cultured in Schneider’s Drosophila medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific, Waltham, MA, USA) at 25 °C.
4.3. RNA Interference (RNAi) in Cultured Drosophila S2 Cells
RNAi was carried out according to [47]. Cells were pelleted and then resuspended in Schneider’s Drosophila medium at a concentration of 1 × 106 cells/mL. 1 mL of cells suspension per well was seeded in a 6-well culture dish (Corning Costar, Cambridge, MA, USA). To perform RNAi, 15 µg of double-stranded RNA (dsRNA) were added to S2 cell culture. After incubation for 1 h at 25 °C, 2 mL of Schneider’s Drosophila medium supplemented with 10% fetal bovine serum were added to each well. Control cells were treated with 15 µg of dsRNA corresponding to eGFP gene fragment that has no homology to Drosophila genes. Cells were grown for 72 h at 25 °C and then harvested for RNA extraction. To minimize off-target effects sequence specific for TRF2 RNAi was chosen by using a web-based tool SnapDragon (https://www.flyrnai.org/snapdragon) [48].
4.4. Production of dsRNA for RNA Interference in Cultured Drosophila S2 Cells
Sequences used as templates for dsRNA synthesis in in vitro transcription reactions were amplified by polymerase chain reaction (PCR). The primers used for PCR harbored T7 RNA polymerase promoter sequence (TAATACGACTCACTATAGGGAGA) at their 5′-termini followed by gene-specific sequence. The PCR products were purified by using QIAquick Gel Extraction Kit (Qiagen, Valencia, CA, USA). To produce dsRNA corresponding to the genes of interest in vitro transcription was performed by using Invitrogen MEGAscript™ T7 Transcription Kit (Thermo Fisher Scientific, Waltham, MA, USA) in accordance with the manufacturer’s instructions. After the completion of the reactions, dsRNA was precipitated by adding 1/10 volume of 3 M ammonium acetate and 2.5 vol of 100% ethanol, and incubating for 30 min at −70 °C. The dsRNA pellets were air-dried and resuspended to a final concentration of 3 µg/mL and analyzed by 1% agarose gel electrophoresis to ensure the size and integrity of dsRNA. For the amplification by PCR of Trf2 gene open reading frame fragment, cDNA reversely transcribed from total RNA extracted from S2 cells was used. For the amplification of eGFP open reading frame fragment pEGFP-C1 vector from Clontech (Clontech, Mountain View, CA, USA) was used. The sense and antisense gene-specific sequences were as follows: Trf2 sense 5′-CTCGTTTCCTCAACTTTCGC-3′, Trf2 antisense 5′-CATCTGTTTCAGACGAGGCA-3′; eGFP sense 5′-TAAACGGCCACAAGTTC-3′, eGFP antisense 5′-GTGTTCTGCTGGTAGTGG-3′.
4.5. RNA extraction and cDNA Preparation
Total RNA from adult flies, salivary glands, and S2-cultured cells was extracted using RNAzol RT reagent (MRC, Cincinnati, OH, USA) according to the manufacturer’s specifications. To remove genomic DNA contamination RNA samples were treated with TURBO DNase (TURBO DNA-free kit, Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocol. cDNA was synthesized from 1 µg of total RNA using MMLV RT kit with random priming (Evrogen, Moscow, Russia).
4.6. Real-Time Quantitative Reverse Transcription PCR (qRT-PCR)
The qRT-PCR analyses were carried out using qPCRmix-HS LowROX and qPCRmix-HS SYBR+LowROX mixes (Evrogen, Moscow, Russia) according to the manufacturer’s protocol on ABI Prism 7500 Sequence Detection System (Applied Biosystems, Foster City, CA, USA). Differences in gene expression levels were determined using the relative quantification (2−ΔΔCt) method [49]. Rpl32 gene was used as the endogenous control. The levels of mRNA expression of the Rpl32 and Trf2 genes were measured using Taqman probe-based chemistry. The levels of mRNA of D1 gene was measured SYBR-based chemistry with dissociation curve analysis. By optimization of SYBR Green method, its performance and quality could be comparable to TaqMan method [50,51]. The following primer pairs and probes were used for amplification: for Rpl32 gene primers Rpl32 sense 5′-CCAGCATACAGGCCCAAGATC-3′, Rpl32 antisense 5′-ACGCACTCTGTTGTCGATACC-3′, TaqMan probe—FAM- CGCACCAAGCACTTCATCCGCCAC-BHQ1; for Trf2 gene primers Trf2 sense 5′-GCTTGCCGCATTTAAACTAAACTC-3′, Trf2 antisense 5′- TTTGGCTCTTGATTTTGTTGTTGC-3′, Trf2 TaqMan probe FAM- AGCCTGGACACCGAAGCGAAAAGC–BHQ1; for D1 gene primers D1 sense 5′-AGAAGCACGAGGACAATGAC-3′, D1 antisense 5′-TCTTTGGACACCTTGCCAG-3′. All reactions were carried out in triplicate. The significance of differences in gene expression levels between samples was evaluated by using REST software (Qiagen, Valencia, CA, USA) based on pair wise fixed reallocation randomization test [52]. P-value of less than 0.05 was considered significant.
4.7. DNA-Directed RNA-Interference
In vivo RNAi-mediated knockdown of Trf2 by using DNA-directed RNA-interference (ddRNAi) was accomplished using the Gal4/UAS system [53,54]. To create Tris plasmid for knock-down of Trf2 a 577 bp and 525 bp cDNA fragments corresponding to Trf2 open reading frame were amplified by PCR using primers harboring restriction sites at their 5′-termini and cloned with head-to-head orientation into the Drosophila transformation pUAST vector [54] to form an inverted repeat. This inverted repeat expressed under control of the UAS element [54] as a stem-loop sequence that trigger RNAi-mediated knock-down of Trf2 gene. The following primer were used for amplification by PCR 577bp and 525bp Trf2 cDNA fragments, respectively, (restriction sites are underlined): Trf2RI sense 5′-CCGAATTCCTCGTTTCCTCAACTTTCGC-3′, Trf2XhoI sense 5′-CCCTCGAGCATCTGTTTCAGACGAGGCA-3′, Trf2XbaI antisense 5′-CCTCTAGACTCGTTTCCTCAACTTTCGC-3′, Trf2XhoI antisense 5′- CCCTCGAGGGTGCTGATGTTTGGTCATTCG-5′. Structure of Tris construct was verified by PCR amplification, DNA sequencing and restriction mapping.
The construct was injected into w1118 preblastoderm embryos as described elsewhere [55]. Using inverse PCR, we found only one copy of UAS-Tris transgenic construct that was located 539 bp upstream of CG4781 5′-termini on chromosome 2R (Figure S2). To corroborate that observed phenotype after Trf2 knock-down was caused only by Trf2 itself, we performed rescue experiment using Drosophila line overexpressing Trf2 protein as described in our earlier work [56].
We used sgs-GAL4 flies as a driver to induce the expression of the UAS-Tris construct in salivary glands [57].
4.8. Computational Analysis of Promoters Regions
Berkeley Drosophila Genome Project (BDGP) neural network promoter prediction software (https://www.fruitfly.org/seq_tools/promoter.html) [58] with default parameters was used for D1 promoter and TSS prediction. Elements Navigation Tool (ElemeNT) (http://lifefaculty.biu.ac.il/gershon-tamar/index.php/element-description/elementv2) [59] with default parameters was used for analysis of core promoter elements. For promoter prediction and analysis, we used sequence from +200 of NM_169261.3 and NM_079562.3 5′-termini to −300 bp of NM_169262.2 5′-termini. All sequences were obtained from NCBI.
4.9. Chromosome Cytology, Immunofluorescence Staining and Microscopy
To analyze attached-X chromosomes in metaphases, brains from third-instar larvae were dissected in saline (NaCl 0.7%) and incubated for 1 h in saline with colcemid (Calbiochem, La Jolla, CA, USA) (0.1 µg/mL). Brains were then treated for 8 min with hypotonic solution (0.5% Na citrate), fixed in 4% paraformaldehyde in 45% acetic acid (1:1) for 10 min, and squashed in 45% acetic acid under a 20 × 20 mm coverslip. Chromosome squashes were frozen in liquid nitrogen. After flipping off the coverslip, slides were washed with PBS, incubated for 5 min in SytoxGreen (1:500, Thermo Fisher Scientific, Waltham, MA, USA), washed with cold PBS, and then mounted in Vectashield H-1000 mounting medium (Vector Laboratories, Burlingame, CA, USA) to reduce fluorescence fading.
S2 cells were fixed in 4% paraformaldehyde for 10 min, washed 3 times in PBS and then permeabilized by incubating with 0.2% Triton X-100 in PBS for 5 min at room temperature. After blocking, cells were incubated overnight with the primary mouse monoclonal antibody anti-Lamin Dm (ADL84, Developmental Studies Hybridoma Bank, Iowa City, Iowa, USA) diluted 1:40 in PBS. Secondary antimouse antibodies (1:100) were conjugated to Cy5 (Thermo Fisher Scientific, Waltham, MA, USA). DNA was stained with SytoxGreen (1:500, Thermo Fisher Scientific, Waltham, MA, USA). The preparations were mounted in Vectashield mounting medium (H-1000, Vector laboratories, Burlingame, CA, USA). The resulting immunofluorescence staining were examined using Leica TCS SP5 confocal microscope. Images were processed using Leica LAS X Lite and Adobe Photoshop software.
Supplementary Materials
Supplementary materials can be found at https://0-www-mdpi-com.brum.beds.ac.uk/1422-0067/21/22/8871/s1.
Author Contributions
Conceptualization, O.B.S. and R.O.C.; methodology, R.O.C. and J.E.V.; software, R.O.C.; validation, O.B.S., R.O.C., and J.E.V.; formal analysis, R.O.C. and J.E.V.; investigation, R.O.C. and J.E.V.; resources, O.B.S., R.O.C., and J.E.V.; data curation, O.B.S.; writing—original draft preparation, O.B.S., R.O.C., and J.E.V.; writing—review and editing, O.B.S., R.O.C., and J.E.V.; visualization, O.B.S., R.O.C., and J.E.V.; supervision, O.B.S.; project administration, O.B.S. ; funding acquisition, O.B.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by RFBR (project No. 20-04-00272a) and was partly conducted in the frame of IDB RAS government program of basic research in 2020 No. 0108-2019-0001. The APC was funded by RFBR (project No. 20-04-00272a) and by IDB RAS government program of basic research in 2020 No. 0108-2019-0001.
Acknowledgments
We thank the Bloomington Stock Center for providing fly stocks. The research was completed using equipment of the Core Centrum of Institute of Developmental Biology RAS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mack, H.; Swisshelm, K. Robertsonian translocations. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 301–305. ISBN 978-0-08-096156-9. [Google Scholar]
- Stankiewicz, P.; Lupski, J.R. Genome architecture, rearrangements and genomic disorders. Trends Genet. 2002, 18, 74–82. [Google Scholar] [CrossRef]
- Garagna, S.; Page, J.; Fernandez-Donoso, R.; Zuccotti, M.; Searle, J.B. The Robertsonian phenomenon in the house mouse: Mutation, meiosis and speciation. Chromosoma 2014, 123, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Rieger, R. A new mechanism for altering chromosome number during karyotype evolution. Theor. Appl. Genet. 1985, 70, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Rieger, R.; Fuchs, J. Alteration of basic chromosome number by fusion-fission cycles. Genome 1995, 38, 1289–1292. [Google Scholar] [CrossRef]
- Jones, K. Robertsonian fusion and centric fission in karyotype evolution of higher plants. Bot. Rev 1998, 64, 273–289. [Google Scholar] [CrossRef]
- Jagannathan, M.; Cummings, R.; Yamashita, Y.M. A conserved function for pericentromeric satellite DNA. ELife 2018, 7, e34122. [Google Scholar] [CrossRef]
- Romanenko, S.A.; Lyapunova, E.A.; Saidov, A.S.; O’Brien, P.C.M.; Serdyukova, N.A.; Ferguson-Smith, M.A.; Graphodatsky, A.S.; Bakloushinskaya, I. Chromosome translocations as a driver of diversification in mole voles ellobius (rodentia, mammalia). Int. J. Mol. Sci. 2019, 20, 4466. [Google Scholar] [CrossRef] [Green Version]
- Sember, A.; de Oliveira, E.A.; Ráb, P.; Bertollo, L.A.C.; de Freitas, N.L.; Viana, P.F.; Yano, C.F.; Hatanaka, T.; Marinho, M.M.F.; de Moraes, R.L.R.; et al. Centric fusions behind the karyotype evolution of neotropical nannostomus pencilfishes (characiforme, lebiasinidae): First insights from a molecular cytogenetic perspective. Genes 2020, 11, 91. [Google Scholar] [CrossRef] [Green Version]
- Matveevsky, S.; Kolomiets, O.; Bogdanov, A.; Alpeeva, E.; Bakloushinskaya, I. Meiotic chromosome contacts as a plausible prelude for robertsonian translocations. Genes 2020, 11, 386. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; O’Brien, P.C.; Milne, B.S.; Graphodatsky, A.S.; Solanky, N.; Trifonov, V.; Rens, W.; Sargan, D.; Ferguson-Smith, M.A. A complete comparative chromosome map for the dog, red fox, and human and its integration with canine genetic maps. Genomics 1999, 62, 189–202. [Google Scholar] [CrossRef]
- Ferguson-Smith, M.A.; Trifonov, V. Mammalian karyotype evolution. Nat. Rev. Genet. 2007, 8, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Kirsch-Volders, M.; Cundari, E.; Verdoodt, B. Towards a unifying model for the metaphase/anaphase transition. Mutagenesis 1998, 13, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.E.; Miller, D.E.; Miller, A.L.; Hawley, R.S. Female meiosis: Synapsis, recombination, and segregation in drosophila melanogaster. Genetics 2018, 208, 875–908. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.S.; Holland, A.J. The impact of mitotic errors on cell proliferation and tumorigenesis. Genes Dev. 2018, 32, 620–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, L.; Voisin, M.; Tatout, C.; Probst, A.V. Structure and function of centromeric and pericentromeric heterochromatin in arabidopsis thaliana. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Zowalaty, A.E.; Baumann, C.; Li, R.; Chen, W.; De La Fuente, R.; Ye, X. Seipin deficiency increases chromocenter fragmentation and disrupts acrosome formation leading to male infertility. Cell Death Dis. 2015, 6, e1817. [Google Scholar] [CrossRef] [Green Version]
- Belli, M.; Vigone, G.; Merico, V.; Redi, C.A.; Garagna, S.; Zuccotti, M. Time-lapse dynamics of the mouse oocyte chromatin organisation during meiotic resumption. BioMed Res. Int. 2014, 2014, 207357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalenskaya, I.A.; Zalensky, A.O. Non-random positioning of chromosomes in human sperm nuclei. Chromosome Res. 2004, 12, 163–173. [Google Scholar] [CrossRef]
- Funabiki, H.; Hagan, I.; Uzawa, S.; Yanagida, M. Cell cycle-dependent specific positioning and clustering of centromeres and telomeres in fission yeast. J. Cell Biol. 1993, 121, 961–976. [Google Scholar] [CrossRef] [Green Version]
- Probst, A.V.; Okamoto, I.; Casanova, M.; El Marjou, F.; Le Baccon, P.; Almouzni, G. A strand-specific burst in transcription of pericentric satellites is required for chromocenter formation and early mouse development. Dev. Cell 2010, 19, 625–638. [Google Scholar] [CrossRef] [Green Version]
- Chubykin, V.L. The structure of the chromocenter in the oocytes, the initiation of homologous pairing and the regulation of the formation of crossing over in drosophila. Tsitologiia 1995, 37, 481–490. [Google Scholar] [PubMed]
- Zuccotti, M.; Garagna, S.; Merico, V.; Monti, M.; Alberto Redi, C. Chromatin organisation and nuclear architecture in growing mouse oocytes. Mol. Cell. Endocrinol. 2005, 234, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Chubykin, V.L. Role of the chromocenter in nonrandom meiotic segregation of nonhomologous chromosomes in drosophila melanogaster females. Russ. J. Genet. 2001, 37, 205–212. [Google Scholar] [CrossRef]
- Chubykin, V.L. Genetic control of the formation and reorganization of chromocenter in drosophila. Russ. J. Genet. 2001, 37, 888–893. [Google Scholar] [CrossRef]
- Fedorova, S.; Nokkala, S.; Chubykin, V.; Omelyanchuk, L. The isolation of a mutation causing abnormal cytokinesis in male and split chromocenter in female meiosis in drosophila melanogaster. Hereditas 2001, 134, 125–134. [Google Scholar] [CrossRef]
- Jagannathan, M.; Cummings, R.; Yamashita, Y.M. The modular mechanism of chromocenter formation in drosophila. Elife 2019, 8. [Google Scholar] [CrossRef]
- Rabenstein, M.D.; Zhou, S.; Lis, J.T.; Tjian, R. TATA box-binding protein (TBP)-related factor 2 (TRF2), a third member of the TBP family. Proc. Natl. Acad. Sci. USA 1999, 96, 4791–4796. [Google Scholar] [CrossRef] [Green Version]
- Simonova, O.B.; Kuzin, B.A.; Georgiev, P.G.; Gerasimova, T.I. Novel regulatory mutation in drosophila melanogaster. Russ. J. Genet. 1992, 28, 164–167. [Google Scholar]
- Vorontsova, J.E.; Modestova, E.A.; Burdina, N.V.; Korochkin, L.I.; Simonova, O.B. Restoring viability of lethal mutants for the leg-arista-wing-complex gene in rescue experiments with transgenic constructs that express the trf2 gene domains in drosophila melanogaster. Dokl. Biol. Sci. 2007, 417, 429–431. [Google Scholar] [CrossRef]
- Simonova, O.B.; Petruk, S.F.; Dzhagaeva, I.V.; Korochkin, L.I. Role of the lawc(p1) mutation in the regulation of the white locus expression in drosophila. Genetika 1998, 34, 349–354. [Google Scholar]
- Wu, C.T. Transvection, nuclear structure, and chromatin proteins. J. Cell Biol. 1993, 120, 587–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopytova, D.V.; Krasnov, A.N.; Kopantceva, M.R.; Nabirochkina, E.N.; Nikolenko, J.V.; Maksimenko, O.; Kurshakova, M.M.; Lebedeva, L.A.; Yerokhin, M.M.; Simonova, O.B.; et al. Two isoforms of drosophila TRF2 are involved in embryonic development, premeiotic chromatin condensation, and proper differentiation of germ cells of both sexes. Mol. Cell. Biol. 2006, 26, 7492–7505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martianov, I.; Brancorsini, S.; Gansmuller, A.; Parvinen, M.; Davidson, I.; Sassone-Corsi, P. Distinct functions of TBP and TLF/TRF2 during spermatogenesis: Requirement of TLF for heterochromatic chromocenter formation in haploid round spermatids. Development 2002, 129, 945–955. [Google Scholar] [PubMed]
- Vorontsova, Y.E.; Cherezov, R.O.; Simonova, O.B. Effect of mutations in lawc/Trf2 gene on development of chromocenter and chromosome disjunction in drosophila melanogaster. Genetika 2013, 49, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Kedmi, A.; Zehavi, Y.; Glick, Y.; Orenstein, Y.; Ideses, D.; Wachtel, C.; Doniger, T.; Waldman Ben-Asher, H.; Muster, N.; Thompson, J.; et al. Drosophila TRF2 is a preferential core promoter regulator. Genes Dev. 2014, 28, 2163–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, P.J.; Greenman, C.D.; Fu, B.; Yang, F.; Bignell, G.R.; Mudie, L.J.; Pleasance, E.D.; Lau, K.W.; Beare, D.; Stebbings, L.A.; et al. Massive genomic rearrangement acquired in a single catastrophic event during cancer development. Cell 2011, 144, 27–40. [Google Scholar] [CrossRef]
- Vorontsova, J.; Cherezov, R.; Simonova, O. The effect of TBP-related factor 2 on chromocenter formation and chromosome segregation in drosophila melanogaster. Chromosom. Abnorm. A Hallmark Manif. Genom. Instab. 2017. [Google Scholar] [CrossRef] [Green Version]
- Andersen, P.R.; Tirian, L.; Vunjak, M.; Brennecke, J. A heterochromatin-dependent transcription machinery drives piRNA expression. Nature 2017, 549, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Siomi, M.C.; Sato, K.; Pezic, D.; Aravin, A.A. PIWI-interacting small RNAs: The vanguard of genome defence. Nat. Rev. Mol. Cell Biol. 2011, 12, 246–258. [Google Scholar] [CrossRef]
- Barra, V.; Fachinetti, D. The dark side of centromeres: Types, causes and consequences of structural abnormalities implicating centromeric DNA. Nat. Commun. 2018, 9, 4340. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Pellman, D. The coordination of nuclear envelope assembly and chromosome segregation in metazoans. Nucleus 2020, 11, 35–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehavi, Y.; Kedmi, A.; Ideses, D.; Juven-Gershon, T. TRF2: TRansForming the view of general transcription factors. Transcription 2015, 6, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duttke, S.H.C.; Doolittle, R.F.; Wang, Y.-L.; Kadonaga, J.T. TRF2 and the evolution of the bilateria. Genes Dev. 2014, 28, 2071–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, A.; Schöttler, P.; Werner, M.; Beinert, N.; Dowe, G.; Burkert, P.; Mourkioti, F.; Dentzer, L.; He, Y.; Deak, P.; et al. Mapping and identification of essential gene functions on the X chromosome of drosophila. EMBO Rep. 2002, 3, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Zitron, A.E.; Hawley, R.S. The genetic analysis of distributive segregation in drosophila melanogaster. I. isolation and characterization of aberrant X segregation (Axs), a mutation defective in chromosome partner choice. Genetics 1989, 122, 801–821. [Google Scholar]
- Clemens, J.C.; Worby, C.A.; Simonson-Leff, N.; Muda, M.; Maehama, T.; Hemmings, B.A.; Dixon, J.E. Use of double-stranded RNA interference in Drosophila cell lines to dissect signal transduction pathways. Proc. Natl. Acad. Sci. USA 2000, 97, 6499–6503. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Comjean, A.; Roesel, C.; Vinayagam, A.; Flockhart, I.; Zirin, J.; Perkins, L.; Perrimon, N.; Mohr, S.E. FlyRNAi.org-the database of the drosophila RNAi screening center and transgenic RNAi project: 2017 update. Nucleic Acids Res. 2017, 45, D672–D678. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Arikawa, E.; Sun, Y.; Wang, J.; Zhou, Q.; Ning, B.; Dial, S.L.; Guo, L.; Yang, J. Cross-platform comparison of SYBR green real-time PCR with TaqMan PCR, microarrays and other gene expression measurement technologies evaluated in the microarray quality control (MAQC) study. BMC Genom. 2008, 9, 328. [Google Scholar] [CrossRef] [Green Version]
- Tajadini, M.; Panjehpour, M.; Javanmard, S.H. Comparison of SYBR green and TaqMan methods in quantitative real-time polymerase chain reaction analysis of four adenosine receptor subtypes. Adv. BioMed Res. 2014, 3. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.A.; Giniger, E.; Maniatis, T.; Ptashne, M. GAL4 activates transcription in drosophila. Nature 1988, 332, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [PubMed]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherezov, R.O.; Vorontsova, I.E.; Mertsalov, I.B.; Kulikova, D.A.; Simonova, O.B. Influence of hairpin RNA, specific to the gene lawc, on expression of overlapping genes of the lawc/Trf2 complex in D. melanogaster. Izv. Akad. Nauk. Ser. Biol. 2013, 2, 133–137. [Google Scholar]
- Phelps, C.B.; Brand, A.H. Ectopic gene expression in drosophila using GAL4 system. Methods 1998, 14, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Reese, M.G. Application of a time-delay neural network to promoter annotation in the drosophila melanogaster genome. Comput. Chem. 2001, 26, 51–56. [Google Scholar] [CrossRef]
- Sloutskin, A.; Danino, Y.M.; Orenstein, Y.; Zehavi, Y.; Doniger, T.; Shamir, R.; Juven-Gershon, T. ElemeNT: A computational tool for detecting core promoter elements. Transcription 2015, 6, 41–50. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic representation of individual genetic crosses for identifying females with attached-X and females with X chromosome nondisjunction. (A) Normal chromosome segregation; (B) F1 progeny from a female with X chromosome nondisjunction; (C) F1 progeny from female carrying attached-X.
Figure 1.
Schematic representation of individual genetic crosses for identifying females with attached-X and females with X chromosome nondisjunction. (A) Normal chromosome segregation; (B) F1 progeny from a female with X chromosome nondisjunction; (C) F1 progeny from female carrying attached-X.

Figure 2.
Females with Trf2 lethal mutations have attached-X chromosomes. (A) Schemes of wild type female metaphase plate carrying normal X (upper panel) and attached-X and Y (lower panel). (B–D) Metaphase plates of larval neuroblasts: (B) wild type female; (C) l(1)G0425^FM7a/Y; (D) l(1)G0356^FM7a/Y. Arrows point to normal X chromosomes, arrowheads point to attached-X chromosomes.
Figure 2.
Females with Trf2 lethal mutations have attached-X chromosomes. (A) Schemes of wild type female metaphase plate carrying normal X (upper panel) and attached-X and Y (lower panel). (B–D) Metaphase plates of larval neuroblasts: (B) wild type female; (C) l(1)G0425^FM7a/Y; (D) l(1)G0356^FM7a/Y. Arrows point to normal X chromosomes, arrowheads point to attached-X chromosomes.

Figure 3.
Levels of X chromosome nondisjunction in females with different Trf2 lethal mutations. (A) Levels of X chromosome nondisjunction. (B) The frequency of females with X chromosome nondisjunction. Below are alleles of Trf2: (l(1)G0425, l(1)G0356, l(1)G0424, l(1)G0295, l(1)G0166, l(1)G0152, l(1)EF520), and two controls: balancer In(1)FM and l(1)G0071 with wild type Trf2 allele.
Figure 3.
Levels of X chromosome nondisjunction in females with different Trf2 lethal mutations. (A) Levels of X chromosome nondisjunction. (B) The frequency of females with X chromosome nondisjunction. Below are alleles of Trf2: (l(1)G0425, l(1)G0356, l(1)G0424, l(1)G0295, l(1)G0166, l(1)G0152, l(1)EF520), and two controls: balancer In(1)FM and l(1)G0071 with wild type Trf2 allele.

Figure 4.
Analysis of putative promoter regulatory elements in Drosophila melanogaster D1 gene promoter region. (Upper panel) schematic D1 gene structure, showing the position and approximate size of exons, introns, and open reading frame in different splice variants according to Flybase. The sequence chosen for bioinformatic analysis performed as described in materials and methods is underlined. Two different promoters involved in transcription regulation of D1 gene designated as P1 and P2. (Lower panel) schematic results of bioinformatic analysis of putative promoter regulatory elements in D1 promoter. TSS+1—transcription start site (TSS). −300—the distance upstream of D1—RC TSS. +200—the distance downstream of D1—RA and D1—RD TSS. BRE—the B recognition element.
Figure 4.
Analysis of putative promoter regulatory elements in Drosophila melanogaster D1 gene promoter region. (Upper panel) schematic D1 gene structure, showing the position and approximate size of exons, introns, and open reading frame in different splice variants according to Flybase. The sequence chosen for bioinformatic analysis performed as described in materials and methods is underlined. Two different promoters involved in transcription regulation of D1 gene designated as P1 and P2. (Lower panel) schematic results of bioinformatic analysis of putative promoter regulatory elements in D1 promoter. TSS+1—transcription start site (TSS). −300—the distance upstream of D1—RC TSS. +200—the distance downstream of D1—RA and D1—RD TSS. BRE—the B recognition element.

Figure 5.
Analysis of Trf2 and D1 genes mRNA levels in salivary glands of the third instar larvae, adult flies and S2 cells. The relative expression levels of Trf2 and D1 genes were examined using qRT-PCR as described in materials and methods. (A) Expression levels of Trf2 and D1 genes detected in salivary glands of: control UAS-Tris/+ larvae and after Trf2 knock-down by ddRNAi (sgs > Tris). (B) Expression levels of Trf2 and D1 genes in lawcp1/EF520 adult flies compared to control EF520/+ flies. (C) Expression levels of Trf2 and D1 genes in lawcp1/Df(1)RA2 adult flies compared to control Df(1)RA2/+ flies; (D) expression levels of Trf2 and D1 genes in the S2 cells after RNAi-mediated silencing of Trf2 (S2-TRFi) compared to control (S2 cells treated with dsRNA corresponding to eGFP gene). Data are presented as the mean ± SD of three independent experiments. * p < 0.05, compared to control.
Figure 5.
Analysis of Trf2 and D1 genes mRNA levels in salivary glands of the third instar larvae, adult flies and S2 cells. The relative expression levels of Trf2 and D1 genes were examined using qRT-PCR as described in materials and methods. (A) Expression levels of Trf2 and D1 genes detected in salivary glands of: control UAS-Tris/+ larvae and after Trf2 knock-down by ddRNAi (sgs > Tris). (B) Expression levels of Trf2 and D1 genes in lawcp1/EF520 adult flies compared to control EF520/+ flies. (C) Expression levels of Trf2 and D1 genes in lawcp1/Df(1)RA2 adult flies compared to control Df(1)RA2/+ flies; (D) expression levels of Trf2 and D1 genes in the S2 cells after RNAi-mediated silencing of Trf2 (S2-TRFi) compared to control (S2 cells treated with dsRNA corresponding to eGFP gene). Data are presented as the mean ± SD of three independent experiments. * p < 0.05, compared to control.

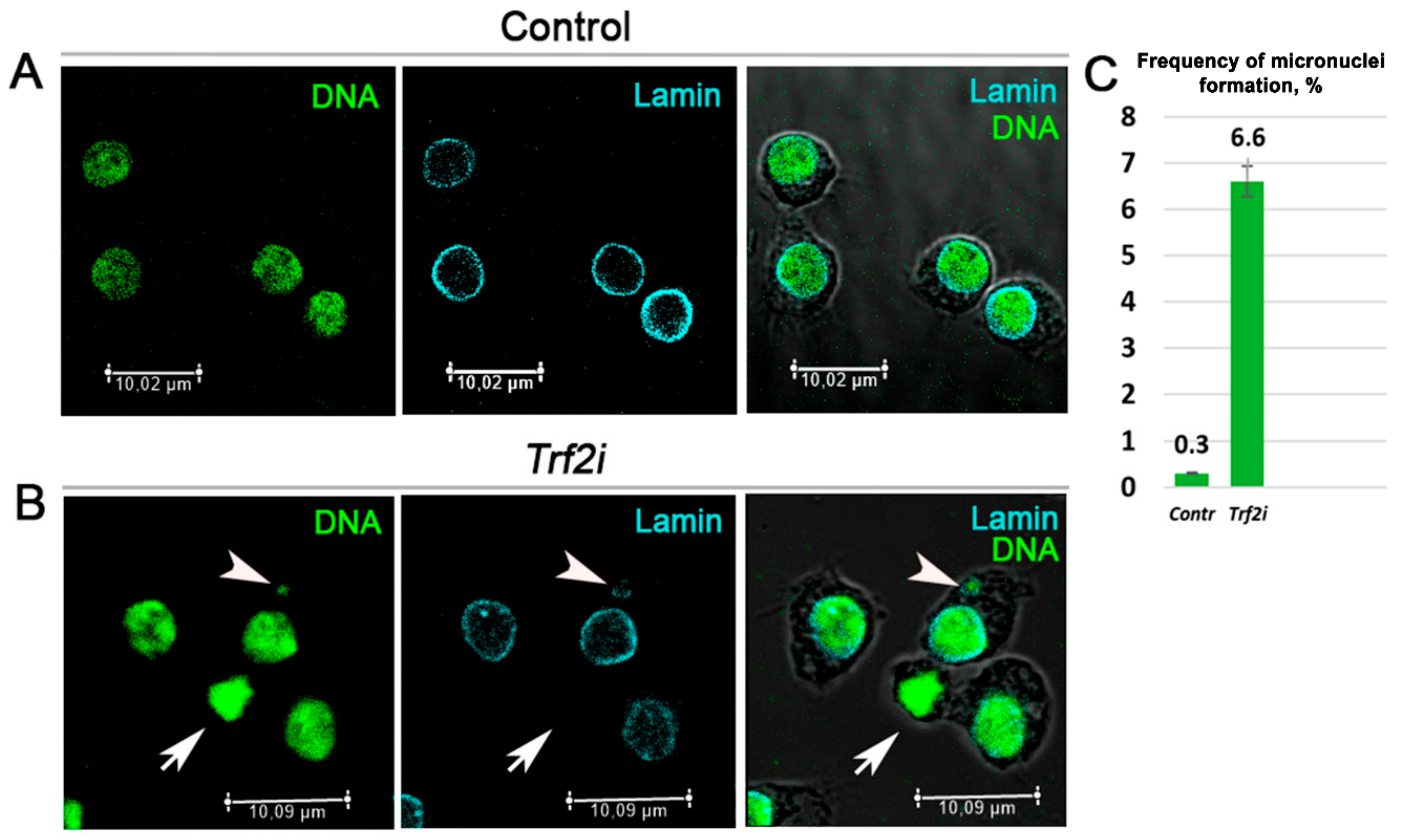
Figure 6.
RNAi-mediated depletion of Trf2 leads to micronuclei formation and disturbs nuclear envelope. Confocal immunofluorescence images of Drosophila S2 cells (A) before and (B) after RNAi-mediated silencing of Trf2. Cells were stained for SytoxGreen to highlight DNA (green) and anti-Lamin Dm to visualized nuclear envelope (cian). The third column represents merged images. Arrowheads pointe to micronucleus, arrows indicate Lamin negative nuclear envelope. (C) Quantification of cells with micronuclei from control (Contr, n = 350) and Trf2 dsRNA transfected cells (Trf2i, n = 229) from three independent experiments. A p-value from Student’s t-test was 0.0047. Error bars: SD.
Figure 6.
RNAi-mediated depletion of Trf2 leads to micronuclei formation and disturbs nuclear envelope. Confocal immunofluorescence images of Drosophila S2 cells (A) before and (B) after RNAi-mediated silencing of Trf2. Cells were stained for SytoxGreen to highlight DNA (green) and anti-Lamin Dm to visualized nuclear envelope (cian). The third column represents merged images. Arrowheads pointe to micronucleus, arrows indicate Lamin negative nuclear envelope. (C) Quantification of cells with micronuclei from control (Contr, n = 350) and Trf2 dsRNA transfected cells (Trf2i, n = 229) from three independent experiments. A p-value from Student’s t-test was 0.0047. Error bars: SD.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
X-chromosome nondisjunction and existence of attached-X chromosomes in females with lethal Trf2 mutations. The first column indicates Trf2 alleles balanced on In(1)FM and control stocks. The second column indicates the presence (“+”) or absence (“−”) of females with attached-X chromosomes. The third column indicates the frequency of X-chromosome nondisjunction Qn. The number (N) of analyzed flies in the progeny of females with X-chromosome nondisjunction is given in brackets. The fourth column indicates the frequency of parental females with X-chromosome nondisjunction Qf. The fifth column indicates the number of parental females analyzed.
Table 1.
X-chromosome nondisjunction and existence of attached-X chromosomes in females with lethal Trf2 mutations. The first column indicates Trf2 alleles balanced on In(1)FM and control stocks. The second column indicates the presence (“+”) or absence (“−”) of females with attached-X chromosomes. The third column indicates the frequency of X-chromosome nondisjunction Qn. The number (N) of analyzed flies in the progeny of females with X-chromosome nondisjunction is given in brackets. The fourth column indicates the frequency of parental females with X-chromosome nondisjunction Qf. The fifth column indicates the number of parental females analyzed.
| Stocks | Attached X | Qn, % (N) | Qf, % | Total |
|---|---|---|---|---|
| l(1)G0425/In(1)FM | + | 7.17 (779) | 40 | 25 |
| l(1)G0356/In(1)FM | + | 11.59 (845) | 46.15 | 26 |
| l(1)G0424/In(1)FM | − | 17.39 (777) | 40 | 25 |
| l(1)G0295/In(1)FM | − | 14.51 (1201) | 48.15 | 27 |
| l(1)G0166/In(1)FM | − | 29.6 (236) | 20 | 30 |
| l(1)G0152/In(1)FM | − | 28.25 (370) | 26.66 | 30 |
| l(1)EF520/In(1)FM | − | 29.12 (88) | 6.67 | 30 |
| Control stocks | ||||
| In(1)FM | − | 1.4 (2445) | 1.54 | 65 |
| l(1)G0071/In(1)FM | − | 1.5 (650) | 1.43 | 70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cherezov, R.O.; Vorontsova, J.E.; Simonova, O.B. TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation. Int. J. Mol. Sci. 2020, 21, 8871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228871
AMA Style
Cherezov RO, Vorontsova JE, Simonova OB. TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation. International Journal of Molecular Sciences. 2020; 21(22):8871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228871
Chicago/Turabian StyleCherezov, Roman O., Julia E. Vorontsova, and Olga B. Simonova. 2020. "TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation" International Journal of Molecular Sciences 21, no. 22: 8871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228871
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.