Molecular Regulation of Copper Homeostasis in the Male Gonad during the Process of Spermatogenesis

 , ,
, ,
Abstract
:1. Introduction
2. An Outline of Systemic Copper Metabolism in Mammals
3. Cellular Copper Uptake, Transport, and Utilization Are Orchestrated by a Closely Integrated Network of Proteins
4. Dysfunction of Spermatogenesis in Both Copper Overload and Deficiency
5. High-Affinity Copper Proteins from the CTR Family Facilitate Copper Transport to Both Sertoli and Germinal Cells
6. Complex Regulation of the Sod1 Gene Encoding Cu,Zn-Superoxide Dismutase (SOD1) in Male Gonads
7. Role of ATP7A, Copper Transporting ATPase in the Protection of Premeiotic and Meiotic Germ Cells from Copper Toxicity
8. Role of the ATP7B Copper-Transporting ATPase in the Regulation of Copper Concentration in Postmeiotic Germ Cells
9. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Tümer, Z.; Møller, L.B. Menkes disease. Eur. J. Hum. Genet. 2010, 18, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldhuis, N.A.; Gaeth, A.P.; Pearson, R.B.; Gabriel, K.; Camakaris, J. The multi-layered regulation of copper translocating P-type ATPases. Biometals 2009, 22, 177–190. [Google Scholar] [CrossRef]
- Lutsenko, S.; Barnes, N.L.; Bartee, M.Y.; Dmitriev, O.Y. Function and regulation of human copper-transporting ATPases. Physiol. Rev. 2007, 87, 1011–1046. [Google Scholar] [CrossRef] [PubMed]
- Kodama, H.; Fujisawa, C.; Bhadhprasit, W. Inherited copper transport disorders: Biochemical mechanisms, diagnosis, and treatment. Curr. Drug Metab. 2012, 13, 237–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vashchenko, G.; MacGillivray, R.T.A. Multi-copper oxidases and human iron metabolism. Nutrients 2013, 5, 2289–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambling, L.; Kennedy, C.; McArdle, H.J. Iron and copper in fetal development. Semin. Cell Dev. Biol. 2011, 22, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Van den Berghe, P.V.E.; Klomp, L.W.J. Posttranslational regulation of copper transporters. J. Biol. Inorg. Chem. 2010, 15, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnlund, J.R. Human whole-body copper metabolism. Am. J. Clin. Nutr. 1998, 67, 960S–964S. [Google Scholar] [CrossRef] [Green Version]
- Bartee, M.Y.; Lutsenko, S. Hepatic copper-transporting ATPase ATP7B: Function and inactivation at the molecular and cellular level. Biometals 2007, 20, 627–637. [Google Scholar] [CrossRef]
- Lenartowicz, M.; Wieczerzak, K.; Krzeptowski, W.; Dobosz, P.; Grzmil, P.; Starzynski, R.; Lipinski, P. Developmental changes in the expression of the Atp7a gene in the liver of mice during the postnatal period. J. Exp. Zool. A Ecol. Genet. Physiol. 2010, 313, 209–217. [Google Scholar]
- Linz, R.; Barnes, N.L.; Zimnicka, A.M.; Kaplan, J.H.; Eipper, B.; Lutsenko, S. Intracellular targeting of copper-transporting ATPase ATP7A in a normal and Atp7b-/- kidney. Am. J. Physiol. Ren. Physiol. 2008, 294, F53–F61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenartowicz, M.; Windak, R.; Tylko, G.; Kowal, M.; Styrna, J. Effects of copper supplementation on the structure and content of elements in kidneys of mosaic mutant mice. Biol. Trace Elem. Res. 2010, 136, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Mercer, J.F.; Livingston, J.; Hall, B.; Paynter, J.A.; Begy, C.; Chandrasekharappa, S.; Lockhart, P.; Grimes, A.; Bhave, M.; Siemieniak, D. Isolation of a partial candidate gene for Menkes disease by positional cloning. Nat. Genet. 1993, 3, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Chelly, J.; Tumer, Z.; Tonnesen, T.; Petterson, A.; Ishikawa-Brush, Y.; Tommerup, N.; Horn, N.; Monaco, A.P. Isolation of a candidate gene for Menkes disease that encodes a potential heavy metal binding protein. Nat. Genet. 1993, 3, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Vulpe, C.; Levinson, B.; Whitney, S.; Packman, S.; Gitschier, J. Isolation of a candidate gene for Menkes disease and evidence that it encodes a copper-transporting ATPase. Nat. Genet. 1993, 3, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Bull, P.C.; Thomas, G.R.; Rommens, J.M.; Forbes, J.R.; Cox, D.W. The Wilson disease gene is a putative copper transporting P-type ATPase similar to the Menkes gene. Nat. Genet. 1993, 5, 327–337. [Google Scholar] [CrossRef]
- Gupta, A.; Lutsenko, S. Human copper transporters: Mechanism, role in human diseases and therapeutic potential. Future Med. Chem. 2009, 1, 1125–1142. [Google Scholar] [CrossRef] [Green Version]
- Nevitt, T.; Ohrvik, H.; Thiele, D.J. Charting the travels of copper in eukaryotes from yeast to mammals. Biochim. Biophys. Acta 2012, 1823, 1580–1593. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.-M.; Gybina, A.A.; Pyatskowit, J.W.; Gitschier, J.; Prohaska, J.R. Copper transport protein (Ctr1) levels in mice are tissue specific and dependent on copper status. J. Nutr. 2006, 136, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Öhrvik, H.; Nose, Y.; Wood, L.K.; Kim, B.-E.; Gleber, S.-C.; Ralle, M.; Thiele, D.J. Ctr2 regulates biogenesis of a cleaved form of mammalian Ctr1 metal transporter lacking the copper- and cisplatin-binding ecto-domain. Proc. Natl. Acad. Sci. USA 2013, 110, E4279–E4288. [Google Scholar] [CrossRef] [Green Version]
- Aller, S.G.; Unger, V.M. Projection structure of the human copper transporter CTR1 at 6-A resolution reveals a compact trimer with a novel channel-like architecture. Proc. Natl. Acad. Sci. USA 2006, 103, 3627–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Smith, K.; Lee, J.; Thiele, D.J.; Petris, M.J. Identification of methionine-rich clusters that regulate copper- stimulated endocytosis of the human Ctr1 copper transporter. J. Biol. Chem. 2004, 279, 17428–17433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wee, N.K.Y.; Weinstein, D.C.; Fraser, S.T.; Assinder, S.J. The mammalian copper transporters CTR1 and CTR2 and their roles in development and disease. Int. J. Biochem. Cell Biol. 2013, 45, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Lutsenko, S.; Tsivkovskii, R.; Walker, J.M. Functional properties of the human copper-transporting ATPase ATP7B (the Wilson’s disease protein) and regulation by metallochaperone Atox1. Ann. N. Y. Acad. Sci. 2003, 986, 204–211. [Google Scholar] [CrossRef]
- Barry, A.N.; Shinde, U.; Lutsenko, S. Structural organization of human Cu-transporting ATPases: Learning from building blocks. J. Biol. Inorg. Chem. 2010, 15, 47–59. [Google Scholar] [CrossRef]
- Rae, T.D.; Torres, A.S.; Pufahl, R.A.; O’Halloran, T.V. Mechanism of Cu,Zn-superoxide dismutase activation by the human metallochaperone hCCS. J. Biol. Chem. 2001, 276, 5166–5176. [Google Scholar] [CrossRef] [Green Version]
- Prohaska, J.R.; Geissler, J.; Brokate, B.; Broderius, M. Copper, zinc-superoxide dismutase protein but not mRNA is lower in copper-deficient mice and mice lacking the copper chaperone for superoxide dismutase. Exp. Biol. Med. 2003, 228, 959–966. [Google Scholar] [CrossRef]
- Wong, P.C.; Waggoner, D.; Subramaniam, J.R.; Tessarollo, L.; Bartnikas, T.B.; Culotta, V.C.; Price, D.L.; Rothstein, J.; Gitlin, J.D. Copper chaperone for superoxide dismutase is essential to activate mammalian Cu/Zn superoxide dismutase. Proc. Natl. Acad. Sci. USA 2000, 97, 2886–2891. [Google Scholar] [CrossRef] [Green Version]
- Casareno, R.L.; Waggoner, D.; Gitlin, J.D. The copper chaperone CCS directly interacts with copper/zinc superoxide dismutase. J. Biol. Chem. 1998, 273, 23625–23628. [Google Scholar] [CrossRef] [Green Version]
- Kako, K.; Tsumori, K.; Ohmasa, Y.; Takahashi, Y.; Munekata, E. The expression of Cox17p in rodent tissues and cells. Eur. J. Biochem. 2000, 267, 6699–6707. [Google Scholar] [CrossRef] [Green Version]
- Prohaska, J.R.; Gybina, A.A. Intracellular copper transport in mammals. J. Nutr. 2004, 134, 1003–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linz, R.; Lutsenko, S. Copper-transporting ATPases ATP7A and ATP7B: Cousins, not twins. J. Bioenerg. Biomembr. 2007, 39, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Lenartowicz, M.; Kennedy, C.; Hayes, H.; McArdle, H.J. Transcriptional regulation of copper metabolism genes in the liver of fetal and neonatal control and iron-deficient rats. Biometals 2015, 28, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenartowicz, M.; Starzynski, R.; Wieczerzak, K.; Krzeptowski, W.; Lipinski, P.; Styrna, J. Alterations in the expression of the Atp7a gene in the early postnatal development of the mosaic mutant mice (Atp7a mo-ms)—An animal model for Menkes disease. Gene Expr. Patterns 2011, 11, 41–47. [Google Scholar] [CrossRef]
- Lutsenko, S.; Gupta, A.; Burkhead, J.L.; Zuzel, V. Cellular multitasking: The dual role of human Cu-ATPases in cofactor delivery and intracellular copper balance. Arch. Biochem. Biophys. 2008, 476, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.D.P.; Cox, D.W. Expression in mouse kidney of membrane copper transporters Atp7a and Atp7b. Nephron 2002, 92, 629–634. [Google Scholar] [CrossRef]
- Bull, P.C.; Cox, D.W. Long range restriction mapping of 13q14.3 focused on the Wilson disease region. Genomics 1993, 16, 593–598. [Google Scholar] [CrossRef]
- La Fontaine, S.; Mercer, J.F.B. Trafficking of the copper-ATPases, ATP7A and ATP7B: Role in copper homeostasis. Arch. Biochem. Biophys. 2007, 463, 149–167. [Google Scholar] [CrossRef]
- Polishchuk, E.V.; Concilli, M.; Iacobacci, S.; Chesi, G.; Pastore, N.; Piccolo, P.; Paladino, S.; Baldantoni, D.; van IJzendoorn, S.C.D.; Chan, J.; et al. Wilson disease protein ATP7B utilizes lysosomal exocytosis to maintain copper homeostasis. Dev. Cell 2014, 29, 686–700. [Google Scholar] [CrossRef] [Green Version]
- Hardman, B.; Manuelpillai, U.; Wallace, E.M.; van de Waasenburg, S.; Cater, M.; Mercer, J.F.B.; Ackland, M.L. Expression and localization of menkes and Wilson copper transporting ATPases in human placenta. Placenta 2004, 25, 512–517. [Google Scholar] [CrossRef] [Green Version]
- Hardman, B.; Luff, S.; Ackland, M.L. Differential intracellular localisation of the Menkes and Wilson copper transporting ATPases in the third trimester human placenta. Placenta 2011, 32, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Michalczyk, A.A.; Rieger, J.; Allen, K.J.; Mercer, J.F.; Ackland, M.L. Defective localization of the Wilson disease protein (ATP7B) in the mammary gland of the toxic milk mouse and the effects of copper supplementation. Biochem. J. 2000, 352 Pt 2, 565–571. [Google Scholar] [CrossRef]
- Lutsenko, S. Modifying factors and phenotypic diversity in Wilson’s disease. Ann. N. Y. Acad. Sci. 2014, 1315, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, J.F.B.; Barnes, N.; Stevenson, J.; Strausak, D.; Llanos, R.M. Copper-induced trafficking of the cU-ATPases: A key mechanism for copper homeostasis. Biometals 2003, 16, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Petris, M.J.; Strausak, D.; Mercer, J.F. The Menkes copper transporter is required for the activation of tyrosinase. Hum. Mol. Genet. 2000, 9, 2845–2851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Itoh, S.; Jeney, V.; Ushio-Fukai, M.; Fukai, T. Essential role for the Menkes ATPase in activation of extracellular superoxide dismutase: Implication for vascular oxidative stress. FASEB J. 2006, 20, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Kang, Z.; Qiao, N.; Liu, G.; Huang, K.; Wang, X.; Pang, C.; Zeng, Q.; Tang, Z.; Li, Y. Chronic Copper Exposure Induces Hypospermatogenesis in Mice by Increasing Apoptosis Without Affecting Testosterone Secretion. Biol. Trace Elem. Res. 2020, 195, 472–480. [Google Scholar] [CrossRef]
- Kowal, M.; Lenartowicz, M.; Pecio, A.; Golas, A.; Blaszkiewicz, T.; Styrna, J. Copper metabolism disorders affect testes structure and gamete quality in male mice. Syst. Biol. Reprod. Med. 2010, 56, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Y.; Yang, X.; Sun, X.D.; Zhuang, C.C.; Xu, F.B.; Li, Y.F. Suppressive Effects of Copper Sulfate Accumulation on the Spermatogenesis of Rats. Biol. Trace Elem. Res. 2016, 174, 356–361. [Google Scholar] [CrossRef]
- Tvrda, E.; Peer, R.; Sikka, S.C.; Agarwal, A. Iron and copper in male reproduction: A double-edged sword. J. Assist. Reprod. Genet. 2015, 32, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Kheirandish, R.; Askari, N.; Babaei, H. Zinc therapy improves deleterious effects of chronic copper administration on mice testes: Histopathological evaluation. Andrologia 2014, 46, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, A.; Sarkar, M.; Biswas, N.M. Dose-dependent effect of copper chloride on male reproductive function in immature rats. Kathmandu Univ. Med. J. (KUMJ) 2005, 3, 392–400. [Google Scholar]
- Akinloye, O.; Abbiyesuku, F.M.; Oguntibeju, O.O.; Arowojolu, A.O.; Truter, E.J. The impact of blood and seminal plasma zinc and copper concentrations on spermogram and hormonal changes in infertile Nigerian men. Reprod. Biol. 2011, 11, 83–98. [Google Scholar] [CrossRef]
- Aydemir, B.; Kiziler, A.R.; Onaran, I.; Alici, B.; Ozkara, H.; Akyolcu, M.C. Impact of Cu and Fe concentrations on oxidative damage in male infertility. Biol. Trace Elem. Res. 2006, 112, 193–203. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Nath, S.; Massanyi, P.; Stawarz, R.; Kacaniova, M.; Kolesarova, A. Copper-induced changes in reproductive functions: In vivo and in vitro effects. Physiol. Res. 2016, 65, 11–22. [Google Scholar] [CrossRef]
- Kasperczyk, A.; Dobrakowski, M.; Czuba, Z.P.; Kapka-Skrzypczak, L.; Kasperczyk, S. Environmental exposure to zinc and copper influences sperm quality in fertile males. Ann. Agric. Environ. Med. 2016, 23, 138–143. [Google Scholar] [CrossRef]
- Roblero, L.; Guadarrama, A.; Lopez, T.; Zegers-Hochschild, F. Effect of copper ion on the motility, viability, acrosome reaction and fertilizing capacity of human spermatozoa in vitro. Reprod. Fertil. Dev. 1996, 8, 871–874. [Google Scholar] [CrossRef]
- Aupperle, H.; Schoon, H.A.; Frank, A. Experimental copper deficiency, chromium deficiency and additional molybdenum supplementation in goats--pathological findings. Acta Vet. Scand. 2001, 42, 311–321. [Google Scholar] [CrossRef]
- Lyubimov, A.V.; Smith, J.A.; Rousselle, S.D.; Mercieca, M.D.; Tomaszewski, J.E.; Smith, A.C.; Levine, B.S. The effects of tetrathiomolybdate (TTM, NSC-714598) and copper supplementation on fertility and early embryonic development in rats. Reprod. Toxicol. 2004, 19, 223–233. [Google Scholar] [CrossRef]
- Van Niekerk, F.E.; Van Niekerk, C.H. The influence of experimentally induced copper deficiency on the fertility of rams. I. Semen parameters and peripheral plasma androgen concentration. J. S. Afr. Vet. Assoc. 1989, 60, 28–31. [Google Scholar]
- Van Niekerk, F.E.; Van Niekerk, C.H. The influence of experimentally induced copper deficiency on the fertility of rams. II. Macro- and microscopic changes in the testes. J. S. Afr. Vet. Assoc. 1989, 60, 32–35. [Google Scholar]
- Yang, H.; Brosel, S.; Acin-Perez, R.; Slavkovich, V.; Nishino, I.; Khan, R.; Goldberg, I.J.; Graziano, J.; Manfredi, G.; Schon, E.A. Analysis of mouse models of cytochrome c oxidase deficiency owing to mutations in Sco2. Hum. Mol. Genet. 2010, 19, 170–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.Y.; Mruk, D.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaffari, R.; Di Bona, K.R.; Riley, C.L.; Richburg, J.H. Copper transporter 1 (CTR1) expression by mouse testicular germ cells, but not Sertoli cells, is essential for functional spermatogenesis. PLoS ONE 2019, 14, e0215522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogórek, M.; Lenartowicz, M.; Starzyński, R.; Jończy, A.; Staroń, R.; Doniec, A.; Krzeptowski, W.; Bednarz, A.; Pierzchała, O.; Lipiński, P.; et al. Atp7a and Atp7b regulate copper homeostasis in developing male germ cells in mice. Metallomics 2017, 9, 1288–1303. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, J.; Ioannoni, R.; Normant, V.; Labbe, S. A role for the transcription factor Mca1 in activating the meiosis- specific copper transporter Mfc1. PLoS ONE 2018, 13, e0201861. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, J.; Thiele, D.J.; Labbe, S.; Puig, S. Dissection of the relative contribution of the Schizosaccharomyces pombe Ctr4 and Ctr5 proteins to the copper transport and cell surface delivery functions. Microbiology 2011, 157, 1021–1031. [Google Scholar] [CrossRef] [Green Version]
- Plante, S.; Ioannoni, R.; Beaudoin, J.; Labbe, S. Characterization of Schizosaccharomyces pombe copper transporter proteins in meiotic and sporulating cells. J. Biol. Chem. 2014, 289, 10168–10181. [Google Scholar] [CrossRef] [Green Version]
- Beaudoin, J.; Ioannoni, R.; Labbe, S. Mfc1 is a novel copper transporter during meiosis. Commun. Integr. Biol. 2012, 5, 118–121. [Google Scholar] [CrossRef]
- Beaudoin, J.; Ekici, S.; Daldal, F.; Ait-Mohand, S.; Guerin, B.; Labbe, S. Copper transport and regulation in Schizosaccharomyces pombe. Biochem. Soc. Trans. 2013, 41, 1679–1686. [Google Scholar] [CrossRef] [Green Version]
- Steiger, D.; Fetchko, M.; Vardanyan, A.; Atanesyan, L.; Steiner, K.; Turski, M.L.; Thiele, D.J.; Georgiev, O.; Schaffner, W. The Drosophila copper transporter Ctr1C functions in male fertility. J. Biol. Chem. 2010, 285, 17089–17097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skandhan, K.P. Review on copper in male reproduction and contraception. Rev. Française Gynécologie D’obstétrique 1992, 87, 594–598. [Google Scholar]
- Celino, F.T.; Yamaguchi, S.; Miura, C.; Ohta, T.; Tozawa, Y.; Iwai, T.; Miura, T. Tolerance of spermatogonia to oxidative stress is due to high levels of Zn and Cu/Zn superoxide dismutase. PLoS ONE 2011, 6, e16938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Trist, B.; Hilton, J.B.; Crouch, P.J.; Hare, D.J.; Double, K.L. Superoxide dismutase 1 in health and disease: How a front-line antioxidant becomes neurotoxic. Angew. Chem. Int. Ed. Engl. 2020. [Google Scholar] [CrossRef]
- Lopes, I.M.D.; de Oliveira, I.M.; Bargi-Souza, P.; Cavallin, M.D.; Kolc, C.S.M.; Khalil, N.M.; Quinaia, S.P.; Romano, M.A.; Romano, R.M. Effects of Silver Nanoparticle Exposure to the Testicular Antioxidant System during the Prepubertal Rat Stage. Chem. Res. Toxicol. 2019, 32, 986–994. [Google Scholar] [CrossRef]
- Gu, W.; Morales, C.; Hecht, N.B. In male mouse germ cells, copper-zinc superoxide dismutase utilizes alternative promoters that produce multiple transcripts with different translation potential. J. Biol. Chem. 1995, 270, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.; Hecht, N.R. Translation of a testis-specific Cu/Zn superoxide dismutase (SOD-1) mRNA is regulated by a 65-kilodalton protein which binds to its 5’ untranslated region. Mol. Cell. Biol. 1996, 16, 4535–4543. [Google Scholar] [CrossRef] [Green Version]
- Ogórek, M.; Herman, S.; Pierzchała, O.; Bednarz, A.; Rajfur, Z.; Baster, Z.; Grzmil, P.; Starzyński, R.R.; Szudzik, M.; Jończy, A.; et al. Molecular machinery providing copper bioavailability for spermatozoa along the epididymial tubule in mouse. Biol. Reprod. 2019, 100, 1505–1520. [Google Scholar] [CrossRef]
- Aitken, R.J.; Buckingham, D.W.; Carreras, A.; Irvine, D.S. Superoxide dismutase in human sperm suspensions: Relationship with cellular composition, oxidative stress, and sperm function. Free Radic. Biol. Med. 1996, 21, 495–504. [Google Scholar] [CrossRef]
- Buffone, M.G.; Calamera, J.C.; Brugo-Olmedo, S.; De Vincentiis, S.; Calamera, M.M.; Storey, B.T.; Doncel, G.F.; Alvarez, J.G. Superoxide dismutase content in sperm correlates with motility recovery after thawing of cryopreserved human spermatozoa. Fertil. Steril. 2012, 97, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Koziorowska-Gilun, M.; Koziorowski, M.; Fraser, L.; Strzezek, J. Antioxidant defence system of boar cauda epididymidal spermatozoa and reproductive tract fluids. Reprod. Domest. Anim. 2011, 46, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Jeon, S.; Song, Y.-J.; Yi, L.S.H. Proteomic analysis of boar spermatozoa and quantity changes of superoxide dismutase 1, glutathione peroxidase, and peroxiredoxin 5 during epididymal maturation. Anim. Reprod. Sci. 2012, 135, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Grant, K.E.; de Oliveira, R.V.; Hennington, B.S.; Govindaraju, A.; Perkins, A.; Stokes, J.; Rowe, D.; Topper, E.; Kaya, A.; Moura, A.; et al. Sperm superoxide dismutase is associated with bull fertility. Reprod. Fertil. Dev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Antinone, S.E.; Ghadge, G.D.; Ostrow, L.W.; Roos, R.P.; Green, W.N. S-acylation of SOD1, CCS, and a stable SOD1-CCS heterodimer in human spinal cords from ALS and non-ALS subjects. Sci. Rep. 2017, 7, 41141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoto, L.G.; Arregui, L.; Sanchez, N.M.; Gomendio, M.; Roldan, E.R.S. Postnatal testicular development in mouse species with different levels of sperm competition. Reproduction 2012, 143, 333–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, M.C. The relationship of copper to DNA damage and damage prevention in humans. Mutat. Res. 2012, 733, 83–91. [Google Scholar] [CrossRef]
- Lenartowicz, M.; Krzeptowski, W.; Lipinski, P.; Grzmil, P.; Starzynski, R.; Pierzchala, O.; Moller, L.B. Mottled Mice and Non-Mammalian Models of Menkes Disease. Front. Mol. Neurosci. 2015, 8, 72. [Google Scholar] [CrossRef] [Green Version]
- La Fontaine, S.; Firth, S.D.; Lockhart, P.J.; Brooks, H.; Camakaris, J.; Mercer, J.F. Intracellular localization and loss of copper responsiveness of Mnk, the murine homologue of the Menkes protein, in cells from blotchy (Mo blo) and brindled (Mo br) mouse mutants. Hum. Mol. Genet. 1999, 8, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Niciu, M.J.; Ma, X.-M.; El Meskini, R.; Pachter, J.S.; Mains, R.E.; Eipper, B.A. Altered ATP7A expression and other compensatory responses in a murine model of Menkes disease. Neurobiol. Dis. 2007, 27, 278–291. [Google Scholar] [CrossRef] [Green Version]
- Kodama, H.; Abe, T.; Takama, M.; Takahashi, I.; Kodama, M.; Nishimura, M. Histochemical localization of copper in the intestine and kidney of macular mice: Light and electron microscopic study. J. Histochem. Cytochem. 1993, 41, 1529–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Styrna, J. Analysis of causes of lethality in mice with the Ms (Mosaic) gene. Genet. Pol. 1977, 18, 61–79. [Google Scholar]
- Lenartowicz, M.; Sasuła, K. Altered copper metabolism in the Mosaic mutant mice. Nutr. Res. 2000, 20, 1467–1471. [Google Scholar] [CrossRef]
- Lenartowicz, M.; Kowal, M.; Buda-Lewandowska, D.; Styrna, J. Pathological structure of the kidney from adult mice with mosaic mutation. J. Inherit. Metab. Dis. 2002, 25, 647–659. [Google Scholar] [CrossRef]
- Kotula-Balak, M.; Lenartowicz, M.; Kowal, M.; Styrna, J.; Bilinska, B. Testicular morphology and expression of aromatase in testes of mice with the mosaic mutation (Atp7a mo-ms). Theriogenology 2007, 67, 423–434. [Google Scholar] [CrossRef]
- Rodriguez, I.; Ody, C.; Araki, K.; Garcia, I.; Vassalli, P. An early and massive wave of germinal cell apoptosis is required for the development of functional spermatogenesis. EMBO J. 1997, 16, 2262–2270. [Google Scholar] [CrossRef] [Green Version]
- Patel, B.N.; Dunn, R.J.; David, S. Alternative RNA splicing generates a glycosylphosphatidylinositol- anchored form of ceruloplasmin in mammalian brain. J. Biol. Chem. 2000, 275, 4305–4310. [Google Scholar] [CrossRef] [Green Version]
- Ramos, D.; Mar, D.; Ishida, M.; Vargas, R.; Gaite, M.; Montgomery, A.; Linder, M.C. Mechanism of Copper Uptake from Blood Plasma Ceruloplasmin by Mammalian Cells. PLoS ONE 2016, 11, e0149516. [Google Scholar] [CrossRef] [Green Version]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out Ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [Green Version]
- De Domenico, I.; Ward, D.M.; di Patti, M.C.B.; Jeong, S.Y.; David, S.; Musci, G.; Kaplan, J. Ferroxidase activity is required for the stability of cell surface ferroportin in cells expressing GPI-ceruloplasmin. EMBO J. 2007, 26, 2823–2831. [Google Scholar] [CrossRef]
- Fortna, R.R.; Watson, H.A.; Nyquist, S.E. Glycosyl phosphatidylinositol-anchored ceruloplasmin is expressed by rat Sertoli cells and is concentrated in detergent-insoluble membrane fractions. Biol. Reprod. 1999, 61, 1042–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leichtmann-Bardoogo, Y.; Cohen, L.A.; Weiss, A.; Marohn, B.; Schubert, S.; Meinhardt, A.; Meyron-Holtz, E.G. Compartmentalization and regulation of iron metabolism proteins protect male germ cells from iron overload. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1519–E1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
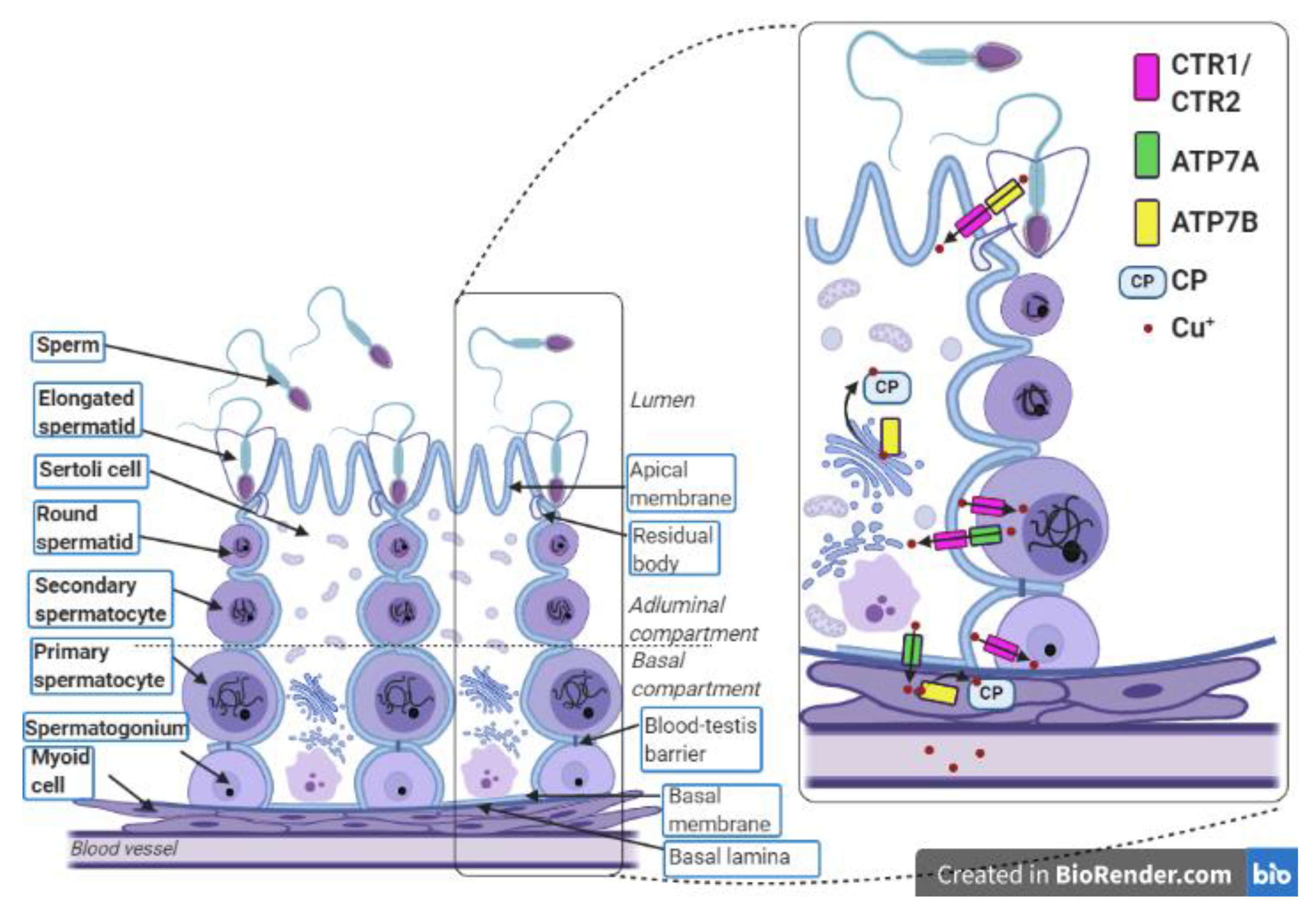{kind=link}
|
Cell Type | Germ Cells | Somatic Cells | ||||
|---|---|---|---|---|---|---|
| Protein |  |  |  |  |  | |
| CTR1 | Cu transport to the cell | Cu transport to the cell | Not found | Cu transport to the cell | Cu transport to the cell? | |
| ATP7A | Not found | Cellular Cu efflux and protection of cells from copper overload | Not found | Cellular Cu efflux and protection of cells from copper overload | Not found | |
| ATP7B | Not found | Not found | Cellular Cu efflux and protection of cells from copper overload | CP metalation | CP metalation | |
| SOD1 | Antioxidant protection | Not found | Antioxidant protection | Antioxidant protection | Not found | |
| CP | Not found | Not found | Not found | Oxidation of Fe++ ions? Cu+ ions sequestering? | Oxidation of Fe++ ions | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herman, S.; Lipiński, P.; Ogórek, M.; Starzyński, R.; Grzmil, P.; Bednarz, A.; Lenartowicz, M. Molecular Regulation of Copper Homeostasis in the Male Gonad during the Process of Spermatogenesis. Int. J. Mol. Sci. 2020, 21, 9053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239053
Herman S, Lipiński P, Ogórek M, Starzyński R, Grzmil P, Bednarz A, Lenartowicz M. Molecular Regulation of Copper Homeostasis in the Male Gonad during the Process of Spermatogenesis. International Journal of Molecular Sciences. 2020; 21(23):9053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239053
Chicago/Turabian StyleHerman, Sylwia, Paweł Lipiński, Mateusz Ogórek, Rafał Starzyński, Paweł Grzmil, Aleksandra Bednarz, and Małgorzata Lenartowicz. 2020. "Molecular Regulation of Copper Homeostasis in the Male Gonad during the Process of Spermatogenesis" International Journal of Molecular Sciences 21, no. 23: 9053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239053