Expression of Connexins 37, 43 and 45 in Developing Human Spinal Cord and Ganglia

 ,
,  , ,
, ,
Abstract
:1. Introduction
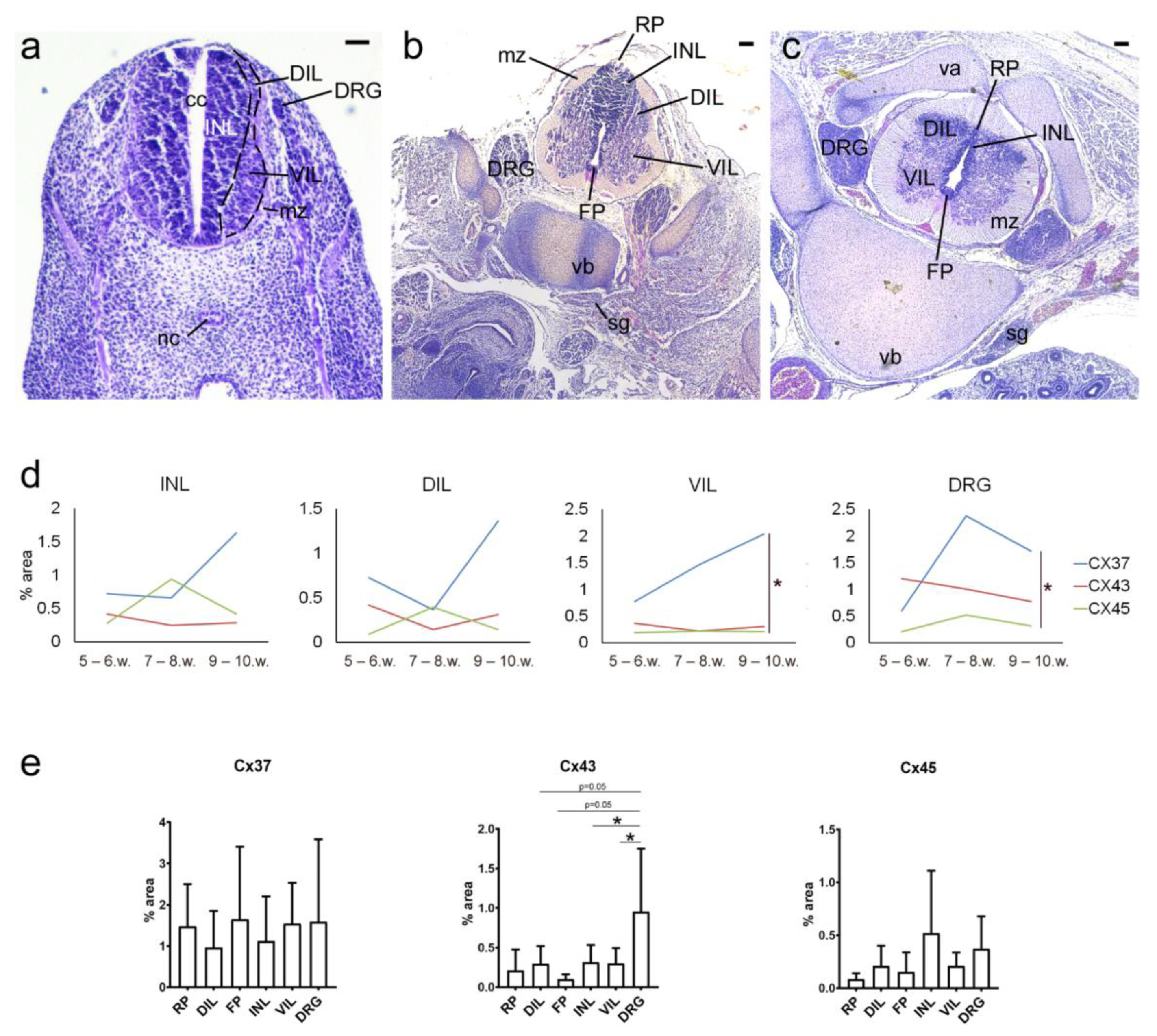
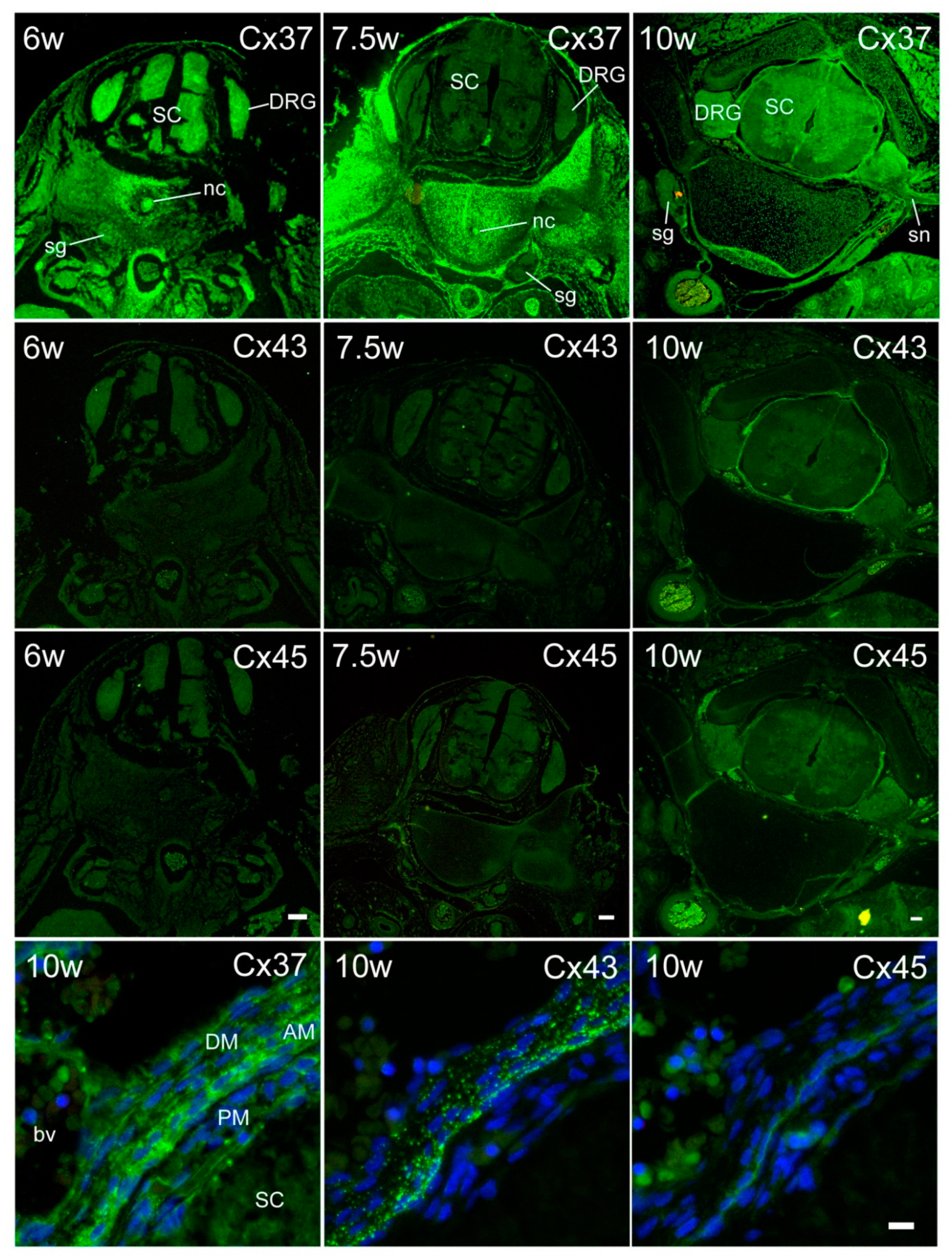
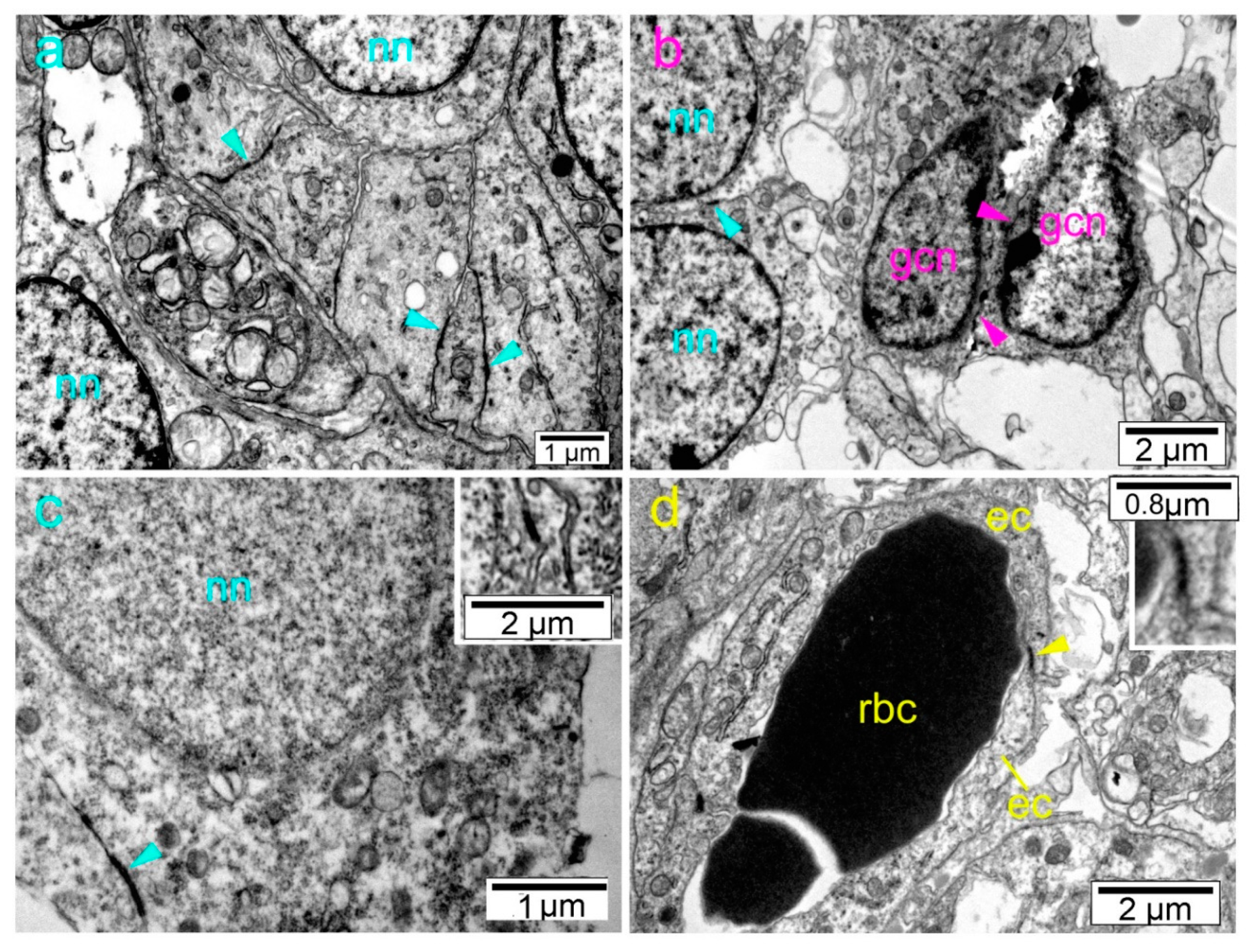
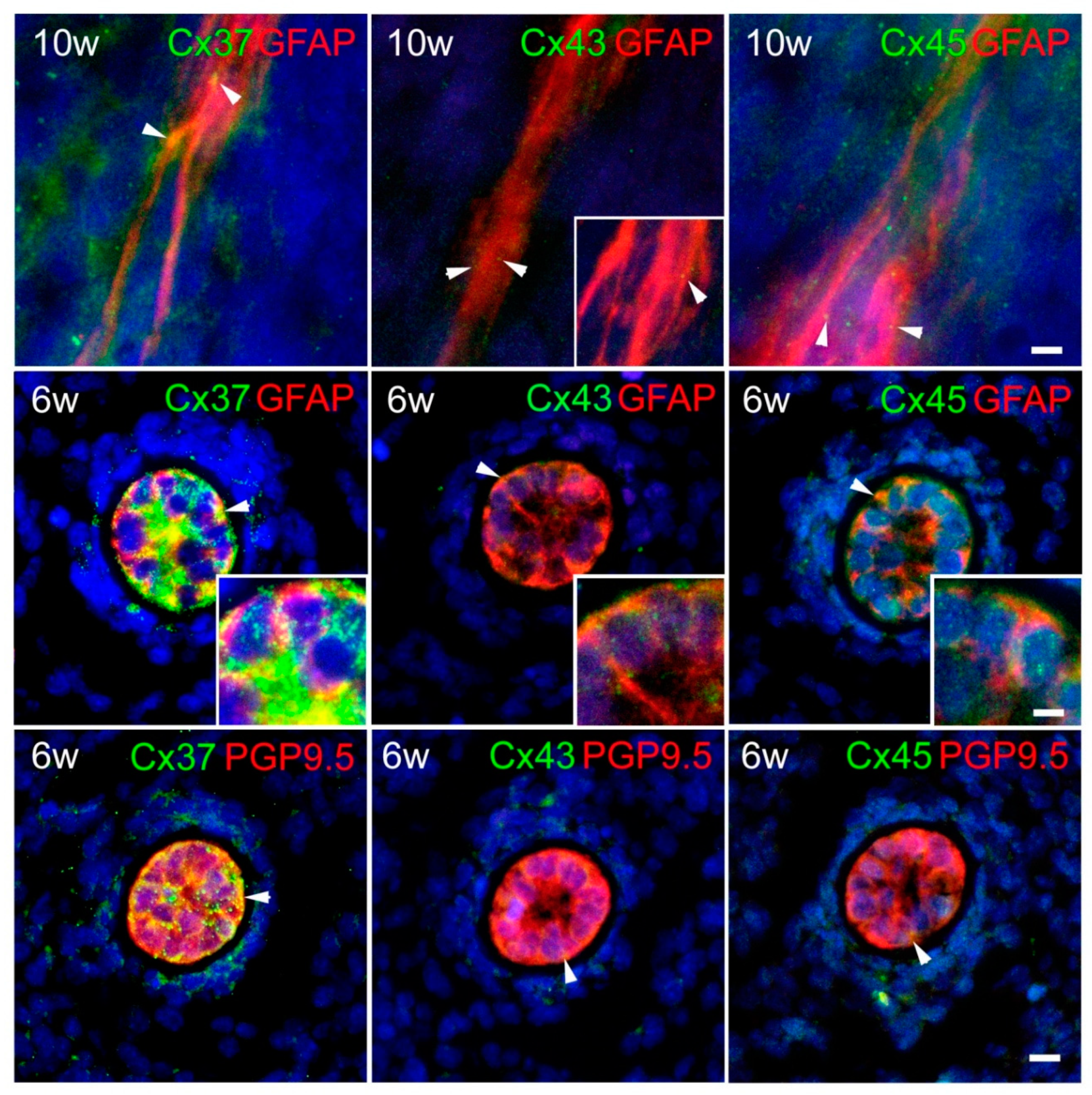
2. Results
3. Discussion
4. Materials and Methods
4.1. Tissue Procurement and Processing
4.2. Immunohistochemistry Procedure
4.3. Transmission Electron Microscopy
4.4. Data Acquisition and Analysis
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SC | spinal cord |
| DRG | dorsal root ganglia |
| Cx | connexin |
| CNS | central nervous system |
| Shh | Sonic hedgehog |
| BMPs | bone morphogenetic proteins |
| INL | inner layer |
| DIL | dorsal part of the intermediate layer |
| VIL | ventral part of the intermediate layer |
| RP | roof plate |
| FP | floor plate |
| PGP9.5 | protein gene product 9.5 |
| PM | pia mater |
| AM | arachnoid |
| DM | dura mater |
| GFAP | glial fibrillary acidic protein |
| ATP | adenosine triphosphate |
| AP | alar plate |
| BP | basal plate |
| TEM | transmission electron microscopy |
| PBS | phosphate buffer saline |
| sg | sympathetic ganglion |
References
- Engelhardt, M.S.J. Functional Neuroanatomy of the Spinal Cord. In Neurological Aspects of Spinal Cord Injury; Springer: Cham, Switzerland, 2017; pp. 19–60. [Google Scholar] [CrossRef]
- Jessell, T.M. Neuronal specification in the spinal cord: Inductive signals and transcriptional codes. Nat. Rev. Genet. 2000, 1, 20–29. [Google Scholar] [CrossRef]
- Noronha, M.J. Embryonic Development of the Spinal Cord and Associated Disorders. In Diseases of the Spinal Cord. Clinical Medicine and the Nervous System; Springer: London, UK, 1992; pp. 79–93. [Google Scholar] [CrossRef]
- Gilbert, S.F. Developmental Biology, 6th ed.; Sinauer Associates: Sunderland, MA, USA, 2000. [Google Scholar]
- Sadler, T.W. Langman’s Medical Embryology. In Medical Embryology, 12th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012. [Google Scholar]
- Vukojevic, K.; Skobic, H.; Saraga-Babic, M. Proliferation and differentiation of glial and neuronal progenitors in the development of human spinal ganglia. Differ. Res. Biol. Divers. 2009, 78, 91–98. [Google Scholar] [CrossRef]
- Kasemeier-Kulesa, J.C.; Kulesa, P.M.; Lefcort, F. Imaging neural crest cell dynamics during formation of dorsal root ganglia and sympathetic ganglia. Development 2005, 132, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.C.; Niu, T.; Alaynick, W.A. Molecular and cellular development of spinal cord locomotor circuitry. Front. Mol. Neurosci. 2015, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restovic, I.; Bocina, I.; Vukojevic, K.; Kero, D.; Filipovic, N.; Raonic, J.; Vucinic, J.; Vukmirovic, F.; Vuckovic, L.; Saraga-Babic, M. Time course and expression pattern of the neuronal markers in the developing human spinal cord. Int. J. Dev. Neurosci. 2019, 74, 1–10. [Google Scholar] [CrossRef]
- Muhr, J.; Graziano, E.; Wilson, S.; Jessell, T.M.; Edlund, T. Convergent inductive signals specify midbrain, hindbrain, and spinal cord identity in gastrula stage chick embryos. Neuron 1999, 23, 689–702. [Google Scholar] [CrossRef] [Green Version]
- Wilson, L.; Maden, M. The mechanisms of dorsoventral patterning in the vertebrate neural tube. Dev. Biol. 2005, 282, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Decrock, E.; De Bock, M.; Wang, N.; Bultynck, G.; Giaume, C.; Naus, C.C.; Green, C.R.; Leybaert, L. Connexin and pannexin signaling pathways, an architectural blueprint for CNS physiology and pathology? Cell. Mol. Life Sci. 2015, 72, 2823–2851. [Google Scholar] [CrossRef]
- Cina, C.; Bechberger, J.F.; Ozog, M.A.; Naus, C.C. Expression of connexins in embryonic mouse neocortical development. J. Comp. Neurol. 2007, 504, 298–313. [Google Scholar] [CrossRef]
- Elias, L.A.; Wang, D.D.; Kriegstein, A.R. Gap junction adhesion is necessary for radial migration in the neocortex. Nature 2007, 448, 901–907. [Google Scholar] [CrossRef]
- Belousov, A.B.; Fontes, J.D. Neuronal gap junctions: Making and breaking connections during development and injury. Trends Neurosci. 2013, 36, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotini, M.; Mayor, R. Connexins in migration during development and cancer. Dev. Biol. 2015, 401, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swayne, L.A.; Bennett, S.A. Connexins and pannexins in neuronal development and adult neurogenesis. BMC Cell Biol. 2016, 17 (Suppl. 1), 10. [Google Scholar] [CrossRef] [Green Version]
- Cina, C.; Maass, K.; Theis, M.; Willecke, K.; Bechberger, J.F.; Naus, C.C. Involvement of the cytoplasmic C-terminal domain of connexin43 in neuronal migration. J. Neurosci. 2009, 29, 2009–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohl, G.; Maxeiner, S.; Willecke, K. Expression and functions of neuronal gap junctions. Nat. Rev. Neurosci. 2005, 6, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Vinken, M. Introduction: Connexins, pannexins and their channels as gatekeepers of organ physiology. Cell. Mol. Life Sci. 2015, 72, 2775–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, R.J.; Lall, V.K.; Maxeiner, S.; Willecke, K.; Deuchars, J.; King, A.E. Localization of neurones expressing the gap junction protein Connexin45 within the adult spinal dorsal horn: A study using Cx45-eGFP reporter mice. Brain Struct. Funct. 2013, 218, 751–765. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.S.; Axelsen, L.N.; Sorgen, P.L.; Verma, V.; Delmar, M.; Holstein-Rathlou, N.H. Gap junctions. Compr. Physiol. 2012, 2, 1981–2035. [Google Scholar] [CrossRef] [Green Version]
- Unwin, P.N.; Zampighi, G. Structure of the junction between communicating cells. Nature 1980, 283, 545–549. [Google Scholar] [CrossRef]
- Sohl, G.; Willecke, K. An update on connexin genes and their nomenclature in mouse and man. Cell Commun. Adhes. 2003, 10, 173–180. [Google Scholar] [CrossRef]
- Oyamada, M.; Oyamada, Y.; Takamatsu, T. Regulation of connexin expression. Biochim. Biophys. Acta 2005, 1719, 6–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, A.; Liu, S.; Li, Y. Gap Junctions in the Nervous System: Probing Functional Connections Using New Imaging Approaches. Front. Cell. Neurosci. 2018, 12, 320. [Google Scholar] [CrossRef] [PubMed]
- Rouach, N.; Avignone, E.; Meme, W.; Koulakoff, A.; Venance, L.; Blomstrand, F.; Giaume, C. Gap junctions and connexin expression in the normal and pathological central nervous system. Biol. Cell 2002, 94, 457–475. [Google Scholar] [CrossRef]
- Fukuda, T. Structural organization of the gap junction network in the cerebral cortex. Neuroscientist 2007, 13, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Lapato, A.S.; Tiwari-Woodruff, S.K. Connexins and pannexins: At the junction of neuro-glial homeostasis & disease. J. Neurosci. Res. 2018, 96, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.H.; De Vuyst, E.; Leybaert, L. The gap junction cellular internet: Connexin hemichannels enter the signalling limelight. Biochem. J. 2006, 397, 1–14. [Google Scholar] [CrossRef]
- Kelsell, D.P.; Dunlop, J.; Stevens, H.P.; Lench, N.J.; Liang, J.N.; Parry, G.; Mueller, R.F.; Leigh, I.M. Connexin 26 mutations in hereditary non-syndromic sensorineural deafness. Nature 1997, 387, 80–83. [Google Scholar] [CrossRef]
- Bergoffen, J.; Scherer, S.S.; Wang, S.; Scott, M.O.; Bone, L.J.; Paul, D.L.; Chen, K.; Lensch, M.W.; Chance, P.F.; Fischbeck, K.H. Connexin mutations in X-linked Charcot-Marie-Tooth disease. Science 1993, 262, 2039–2042. [Google Scholar] [CrossRef]
- Belousov, A.B.; Fontes, J.D.; Freitas-Andrade, M.; Naus, C.C. Gap junctions and hemichannels: Communicating cell death in neurodevelopment and disease. BMC Cell Biol. 2017, 18, 4. [Google Scholar] [CrossRef] [Green Version]
- Paznekas, W.A.; Boyadjiev, S.A.; Shapiro, R.E.; Daniels, O.; Wollnik, B.; Keegan, C.E.; Innis, J.W.; Dinulos, M.B.; Christian, C.; Hannibal, M.C.; et al. Connexin 43 (GJA1) mutations cause the pleiotropic phenotype of oculodentodigital dysplasia. Am. J. Hum. Genet. 2003, 72, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, R. Connexins in health and disease. Clin. Exp. Neuroimmunol. 2018, 9, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, S.H.; Folsom, T.D.; Reutiman, T.J.; Lee, S. Expression of astrocytic markers aquaporin 4 and connexin 43 is altered in brains of subjects with autism. Synapse 2008, 62, 501–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.Y.; Cui, Y.; Deng, F.; Feng, J.C. Connexin: A potential novel target for protecting the central nervous system? Neural Regen. Res. 2015, 10, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Guerra, M.M.; Henzi, R.; Ortloff, A.; Lichtin, N.; Vio, K.; Jimenez, A.J.; Dominguez-Pinos, M.D.; Gonzalez, C.; Jara, M.C.; Hinostroza, F.; et al. Cell Junction Pathology of Neural Stem Cells Is Associated With Ventricular Zone Disruption, Hydrocephalus, and Abnormal Neurogenesis. J. Neuropathol. Exp. Neurol. 2015, 74, 653–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbiani, G.; Reali, C.; Valentin-Kahan, A.; Rehermann, M.I.; Fagetti, J.; Falco, M.V.; Russo, R.E. Connexin Signaling Is Involved in the Reactivation of a Latent Stem Cell Niche after Spinal Cord Injury. J. Neurosci. 2020, 40, 2246–2258. [Google Scholar] [CrossRef]
- Russo, R.E.; Reali, C.; Radmilovich, M.; Fernandez, A.; Trujillo-Cenoz, O. Connexin 43 delimits functional domains of neurogenic precursors in the spinal cord. J. Neurosci. 2008, 28, 3298–3309. [Google Scholar] [CrossRef] [Green Version]
- Genet, N.; Bhatt, N.; Bourdieu, A.; Hirschi, K.K. Multifaceted Roles of Connexin 43 in Stem Cell Niches. Curr. Stem Cell Rep. 2018, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Recabal, A.; Elizondo-Vega, R.; Philippot, C.; Salgado, M.; Lopez, S.; Palma, A.; Tarifeno-Saldivia, E.; Timmermann, A.; Seifert, G.; Caprile, T.; et al. Connexin-43 Gap Junctions Are Responsible for the Hypothalamic Tanycyte-Coupled Network. Front. Cell. Neurosci. 2018, 12, 406. [Google Scholar] [CrossRef]
- Casse, F.; Richetin, K.; Toni, N. Astrocytes’ Contribution to Adult Neurogenesis in Physiology and Alzheimer’s Disease. Front. Cell. Neurosci. 2018, 12, 432. [Google Scholar] [CrossRef] [Green Version]
- Nagy, J.I.; Rash, J.E. Electrical Synapses: New Rules for Assembling an Old Structure Asymmetrically. Curr. Biol. 2017, 27, R1214–R1216. [Google Scholar] [CrossRef]
- Mayorquin, L.C.; Rodriguez, A.V.; Sutachan, J.J.; Albarracin, S.L. Connexin-Mediated Functional and Metabolic Coupling Between Astrocytes and Neurons. Front. Mol. Neurosci. 2018, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, X.H. Expression of nNOS, Pax3 and Cx43 proteins in early developing posterior horn of embryonic and fetal human spinal cord. Nan Fang Yi Ke Da Xue Xue Bao 2009, 29, 1651–1653. [Google Scholar] [PubMed]
- Chang, Q.; Gonzalez, M.; Pinter, M.J.; Balice-Gordon, R.J. Gap junctional coupling and patterns of connexin expression among neonatal rat lumbar spinal motor neurons. J. Neurosci. 1999, 19, 10813–10828. [Google Scholar] [CrossRef] [PubMed]
- Belliveau, D.J.; Bani-Yaghoub, M.; McGirr, B.; Naus, C.C.; Rushlow, W.J. Enhanced neurite outgrowth in PC12 cells mediated by connexin hemichannels and ATP. J. Biol. Chem. 2006, 281, 20920–20931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jourdeuil, K.; Taneyhill, L.A. Spatiotemporal expression pattern of Connexin 43 during early chick embryogenesis. Gene Expr. Patterns 2018, 27, 67–75. [Google Scholar] [CrossRef]
- Zhao, Y.; Xin, Y.; He, Z.; Hu, W. Function of Connexins in the Interaction between Glial and Vascular Cells in the Central Nervous System and Related Neurological Diseases. Neural Plast. 2018, 2018, 6323901. [Google Scholar] [CrossRef]
- Bautista, W.; Rash, J.E.; Vanderpool, K.G.; Yasumura, T.; Nagy, J.I. Re-evaluation of connexins associated with motoneurons in rodent spinal cord, sexually dimorphic motor nuclei and trigeminal motor nucleus. Eur. J. Neurosci. 2014, 39, 757–770. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.M.; McWhorter, A.R. Role of connexin37 and connexin40 in vascular development. Cell Commun. Adhes. 2003, 10, 379–385. [Google Scholar] [CrossRef]
- Hampl, R.; Bicikova, M.; Sosvorova, L. How hormones influence composition and physiological function of the brain-blood barrier. Physiol. Res. 2015, 64, S259–S264. [Google Scholar] [CrossRef]
- De Bock, M.; Culot, M.; Wang, N.; Bol, M.; Decrock, E.; De Vuyst, E.; da Costa, A.; Dauwe, I.; Vinken, M.; Simon, A.M.; et al. Connexin channels provide a target to manipulate brain endothelial calcium dynamics and blood-brain barrier permeability. J. Cereb. Blood Flow Metab. 2011, 31, 1942–1957. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Ozono, Y.; Mikuriya, S.; Kohro, Y.; Tozaki-Saitoh, H.; Iwatsuki, K.; Uneyama, H.; Ichikawa, R.; Salter, M.W.; Tsuda, M.; et al. Dorsal horn neurons release extracellular ATP in a VNUT-dependent manner that underlies neuropathic pain. Nat. Commun. 2016, 7, 12529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruangvoravat, C.P.; Lo, C.W. Connexin 43 expression in the mouse embryo: Localization of transcripts within developmentally significant domains. Dev. Dyn. 1992, 194, 261–281. [Google Scholar] [CrossRef]
- Bannerman, P.; Nichols, W.; Puhalla, S.; Oliver, T.; Berman, M.; Pleasure, D. Early migratory rat neural crest cells express functional gap junctions: Evidence that neural crest cell survival requires gap junction function. J. Neurosci. Res. 2000, 61, 605–615. [Google Scholar] [CrossRef]
- Tonkin, R.S.; Mao, Y.; O’Carroll, S.J.; Nicholson, L.F.; Green, C.R.; Gorrie, C.A.; Moalem-Taylor, G. Gap junction proteins and their role in spinal cord injury. Front. Mol. Neurosci. 2014, 7, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, M.V.; Zukin, R.S. Electrical coupling and neuronal synchronization in the Mammalian brain. Neuron 2004, 41, 495–511. [Google Scholar] [CrossRef] [Green Version]
- de Rivero Vaccari, J.C.; Corriveau, R.A.; Belousov, A.B. Gap junctions are required for NMDA receptor dependent cell death in developing neurons. J. Neurophysiol. 2007, 98, 2878–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, J.I.; Rash, J.E. Astrocyte and oligodendrocyte connexins of the glial syncytium in relation to astrocyte anatomical domains and spatial buffering. Cell Commun. Adhes. 2003, 10, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Lagos-Cabre, R.; Brenet, M.; Diaz, J.; Perez, R.D.; Perez, L.A.; Herrera-Molina, R.; Quest, A.F.G.; Leyton, L. Intracellular Ca(2+) Increases and Connexin 43 Hemichannel Opening Are Necessary but Not Sufficient for Thy-1-Induced Astrocyte Migration. Int. J. Mol. Sci. 2018, 19, 2179. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.R.; Roh, D.H.; Yoon, S.Y.; Kwon, S.G.; Choi, H.S.; Han, H.J.; Beitz, A.J.; Lee, J.H. Astrocyte sigma-1 receptors modulate connexin 43 expression leading to the induction of below-level mechanical allodynia in spinal cord injured mice. Neuropharmacology 2016, 111, 34–46. [Google Scholar] [CrossRef]
- Li, H.L.; Huang, Y.; Zhou, Y.L.; Teng, R.H.; Zhou, S.Z.; Lin, J.P.; Yang, Y.; Zhu, S.M.; Xu, H.; Yao, Y.X. C-X-C Motif Chemokine 10 Contributes to the Development of Neuropathic Pain by Increasing the Permeability of the Blood-Spinal Cord Barrier. Front. Immunol. 2020, 11, 477. [Google Scholar] [CrossRef]
- Beyer, E.C.; Gemel, J.; Martinez, A.; Berthoud, V.M.; Valiunas, V.; Moreno, A.P.; Brink, P.R. Heteromeric mixing of connexins: Compatibility of partners and functional consequences. Cell Commun. Adhes. 2001, 8, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dermietzel, R.; Gao, Y.; Scemes, E.; Vieira, D.; Urban, M.; Kremer, M.; Bennett, M.V.; Spray, D.C. Connexin43 null mice reveal that astrocytes express multiple connexins. Brain Res. Brain Res. Rev. 2000, 32, 45–56. [Google Scholar] [CrossRef]
- Elenes, S.; Martinez, A.D.; Delmar, M.; Beyer, E.C.; Moreno, A.P. Heterotypic docking of Cx43 and Cx45 connexons blocks fast voltage gating of Cx43. Biophys. J. 2001, 81, 1406–1418. [Google Scholar] [CrossRef] [Green Version]
- Martinez, A.D.; Hayrapetyan, V.; Moreno, A.P.; Beyer, E.C. Connexin43 and connexin45 form heteromeric gap junction channels in which individual components determine permeability and regulation. Circ. Res. 2002, 90, 1100–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterhager, E.; Kidder, G.M. Gap junction connexins in female reproductive organs: Implications for women’s reproductive health. Hum. Reprod. Update 2015, 21, 340–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, A.M.; Goodenough, D.A.; Li, E.; Paul, D.L. Female infertility in mice lacking connexin 37. Nature 1997, 385, 525–529. [Google Scholar] [CrossRef]
- Williams, J.R. The Declaration of Helsinki and public health. Bull. World Health Organ. 2008, 86, 650–652. [Google Scholar] [CrossRef]
- O’Rahilly, R.; Gardner, E. The timing and sequence of events in the development of the human nervous system during the embryonic period proper. Z. Anat. Entwickl. 1971, 134, 1–12. [Google Scholar] [CrossRef]
- Sheehan, D.C.; Hrapchak, B.B. Theory and Practice of Histotechnology; Mosby: Maryland Heights, MO, USA, 1980. [Google Scholar]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Weeks) | Crown-Rump Length (CRL) (mm) | Carnegie Stage | No. |
|---|---|---|---|
| 5 | 8 | 14 | 1 |
| 6 | 14 | 16 | 2 |
| 7 | 21 | 20 | 2 |
| 8 | 27 | 22 | 1 |
| 9 | 32 | - | 2 |
| 10 | 36 | - | 2 |
| Primary | Antibody | Code no. | Host | Dilution | Source |
| Anti-Cx37/GJA4 | ab181701 | Rabbit | 1:100 | Abcam, Cambridge, UK | |
| Anti-Connexin 43/GJA1 | ab87645 | Goat | 1:300 | ||
| Anti-Connexin 45/GJA7/Cx45 | ab135474 | Rabbit | 1:100 | ||
| Anti-GFAP | ab53554 | Goat | 1:100 | ||
| Anti-GFAP (2E1) | sc-33673 | Mouse | 1:50 | Santa Cruz Biotechnology Inc., Dallas, TX, USA | |
| PGP9.5 Monoclonal Antibody (BH7) | 480012 | Mouse | 1:500 | Invitrogen | |
| Secondary | AlexaFluor®488 Donkey Anti-Rabbit IgG | 711-545-152 | Donkey | 1:400 | Jackson Immuno Research Laboratories Inc., Baltimore, PA, USA |
| Rhodamine Red™-X (RRX) Donkey Anti-Goat IgG | 705-295-003 | ||||
| Rhodamine Red™-X (RRX) Donkey Anti-Mouse IgG | 715-295-151 | ||||
| AlexaFluor®488 Donkey Anti-Goat IgG | 705-545-003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurić, M.; Zeitler, J.; Vukojević, K.; Bočina, I.; Grobe, M.; Kretzschmar, G.; Saraga-Babić, M.; Filipović, N. Expression of Connexins 37, 43 and 45 in Developing Human Spinal Cord and Ganglia. Int. J. Mol. Sci. 2020, 21, 9356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249356
Jurić M, Zeitler J, Vukojević K, Bočina I, Grobe M, Kretzschmar G, Saraga-Babić M, Filipović N. Expression of Connexins 37, 43 and 45 in Developing Human Spinal Cord and Ganglia. International Journal of Molecular Sciences. 2020; 21(24):9356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249356
Chicago/Turabian StyleJurić, Marija, Julia Zeitler, Katarina Vukojević, Ivana Bočina, Maximilian Grobe, Genia Kretzschmar, Mirna Saraga-Babić, and Natalija Filipović. 2020. "Expression of Connexins 37, 43 and 45 in Developing Human Spinal Cord and Ganglia" International Journal of Molecular Sciences 21, no. 24: 9356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249356