Spadin Modulates Astrocytic Passive Conductance via Inhibition of TWIK-1/TREK-1 Heterodimeric Channels

 , ,
, ,  and
and 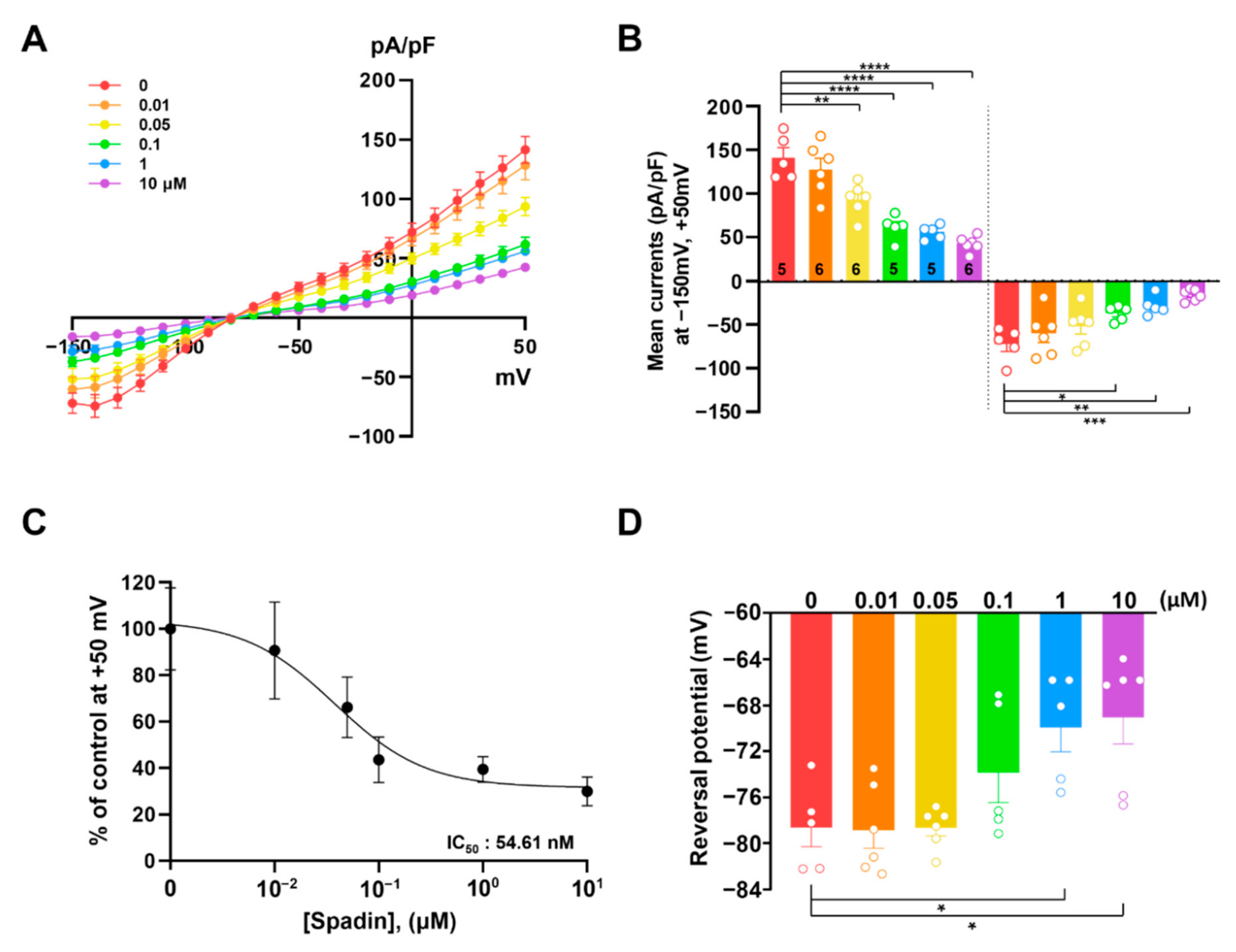{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Spadin Inhibits TREK-1 Mediated K+ Currents in Cultured Astrocytes
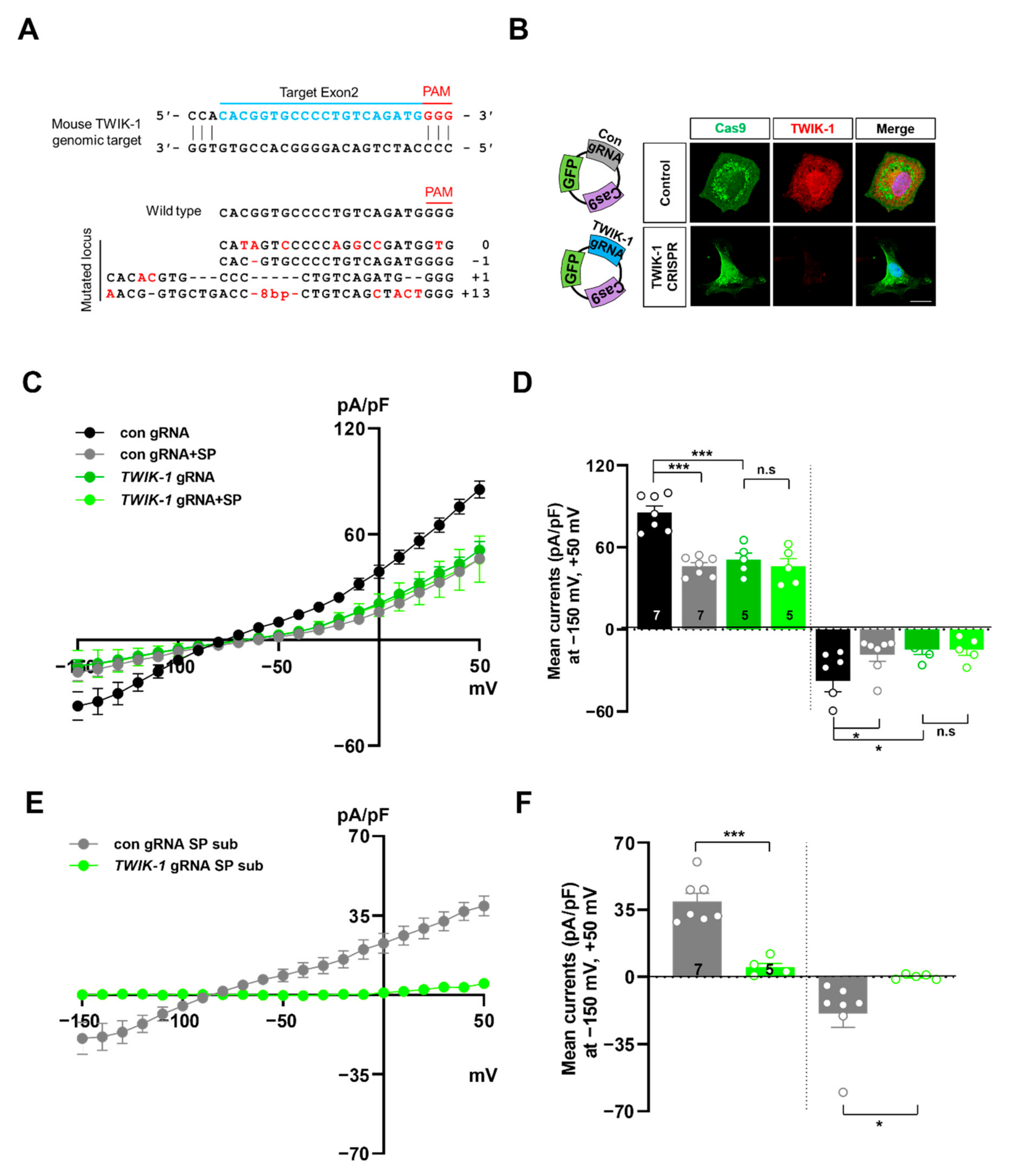
2.2. Spadin Inhibits TWIK-1 Mediated K+ Currents in Cultured Astrocytes
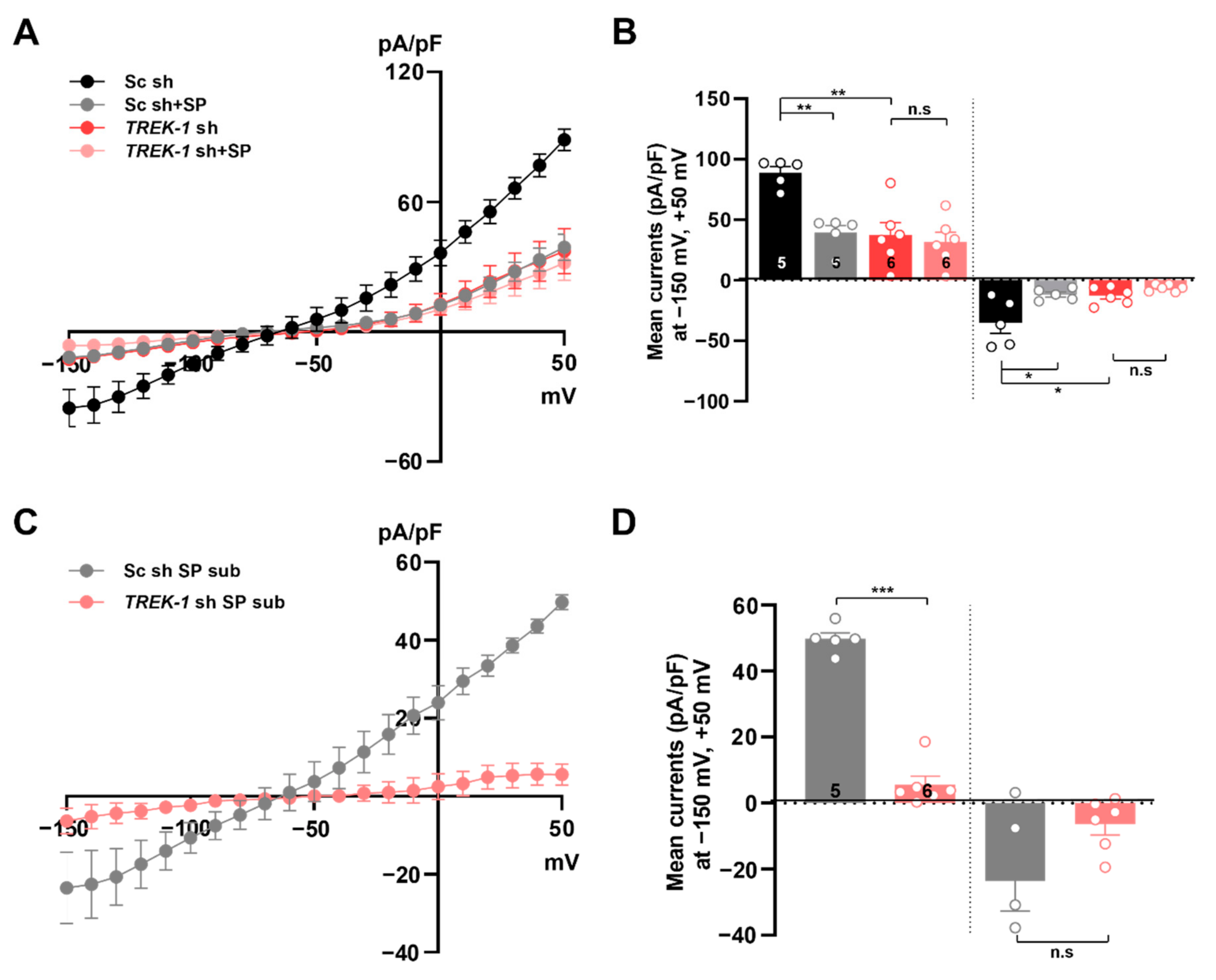
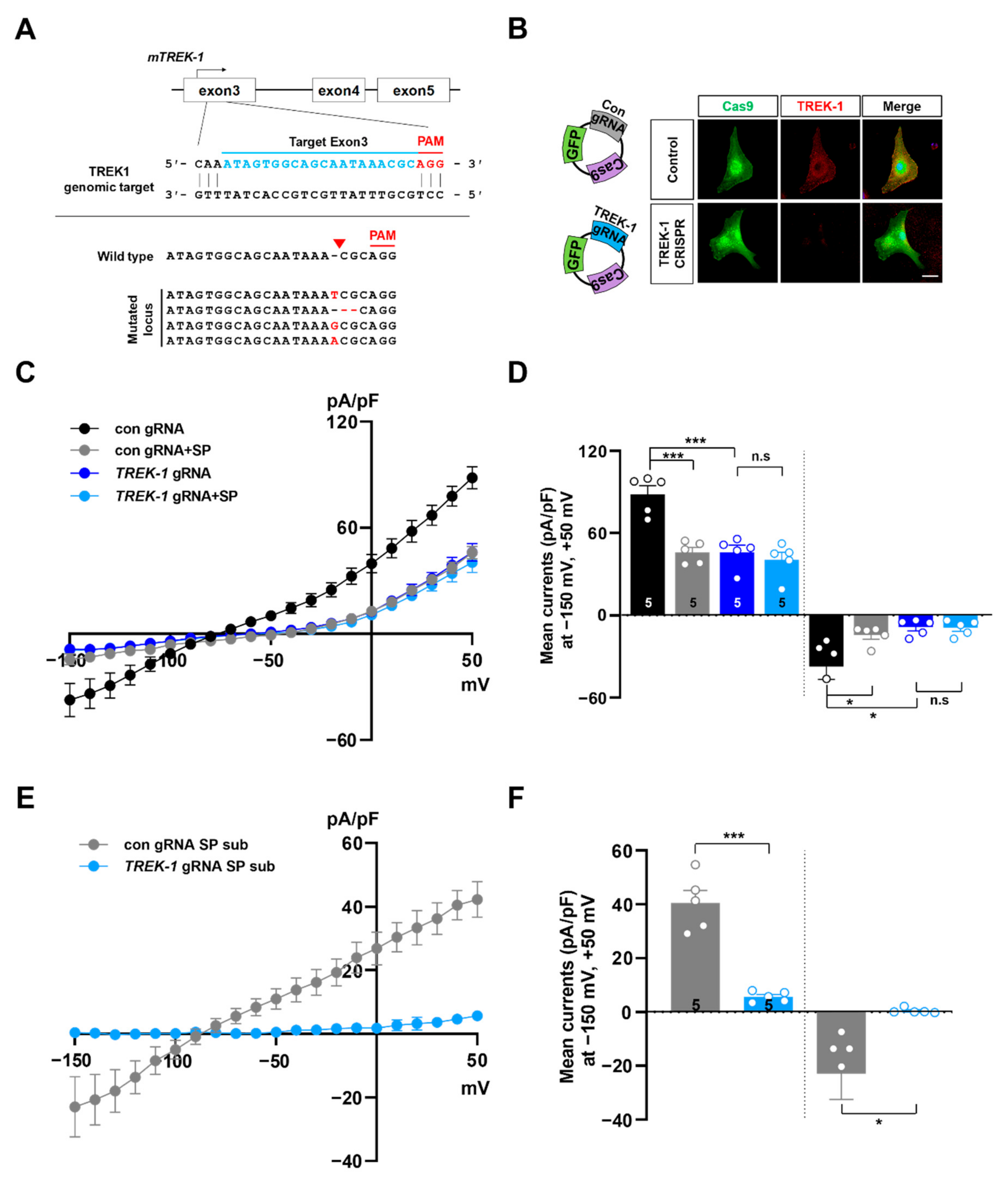
2.3. Knockout of TREK-1 or TWIK-1 with CRISPR/Cas9 System Abolishes Spadin-Sensitive Currents in Cultured Astrocytes
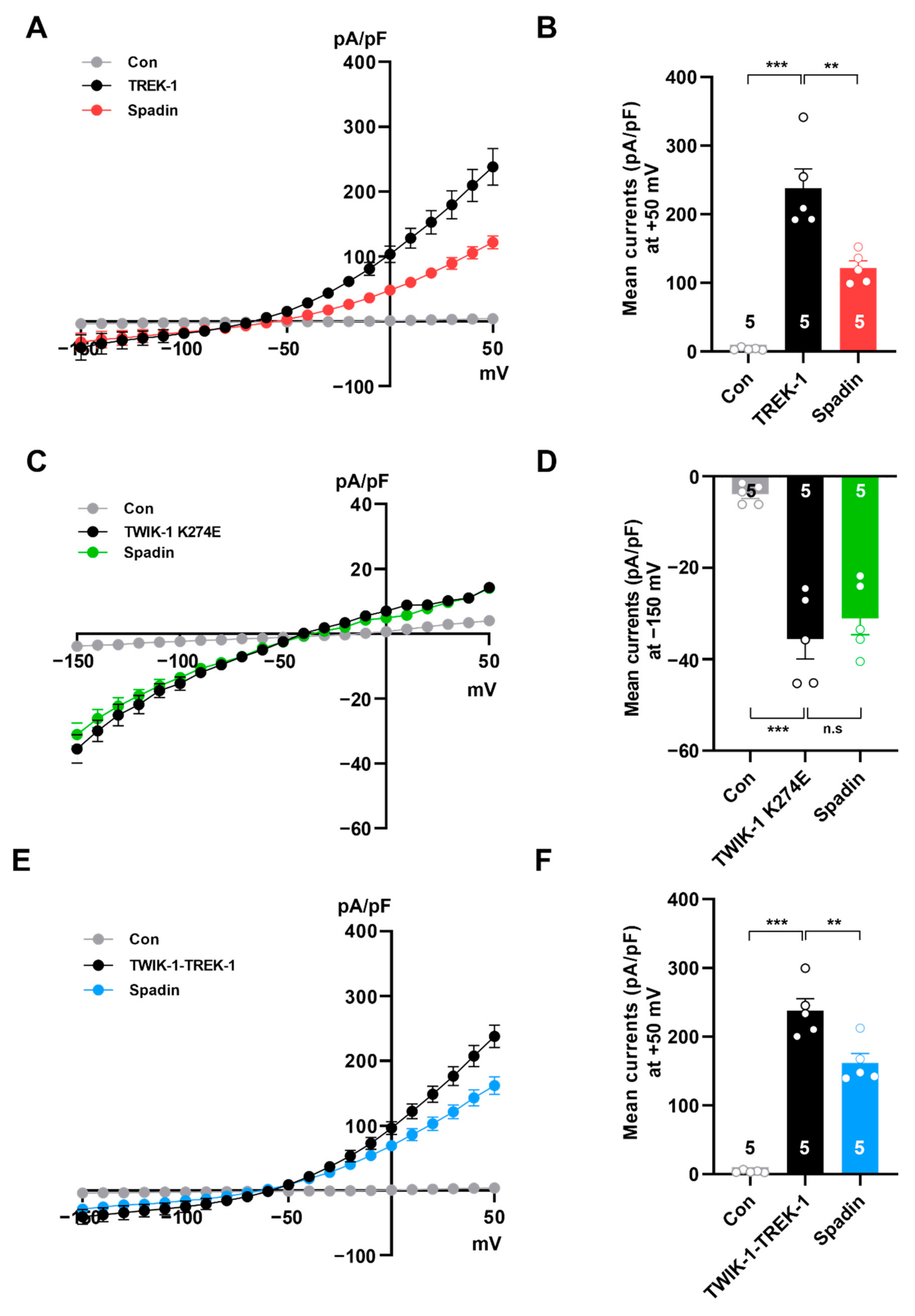
2.4. Spadin Inhibits Concatenated TWIK-1-TREK-1 Heterodimers in Heterologous Expression System
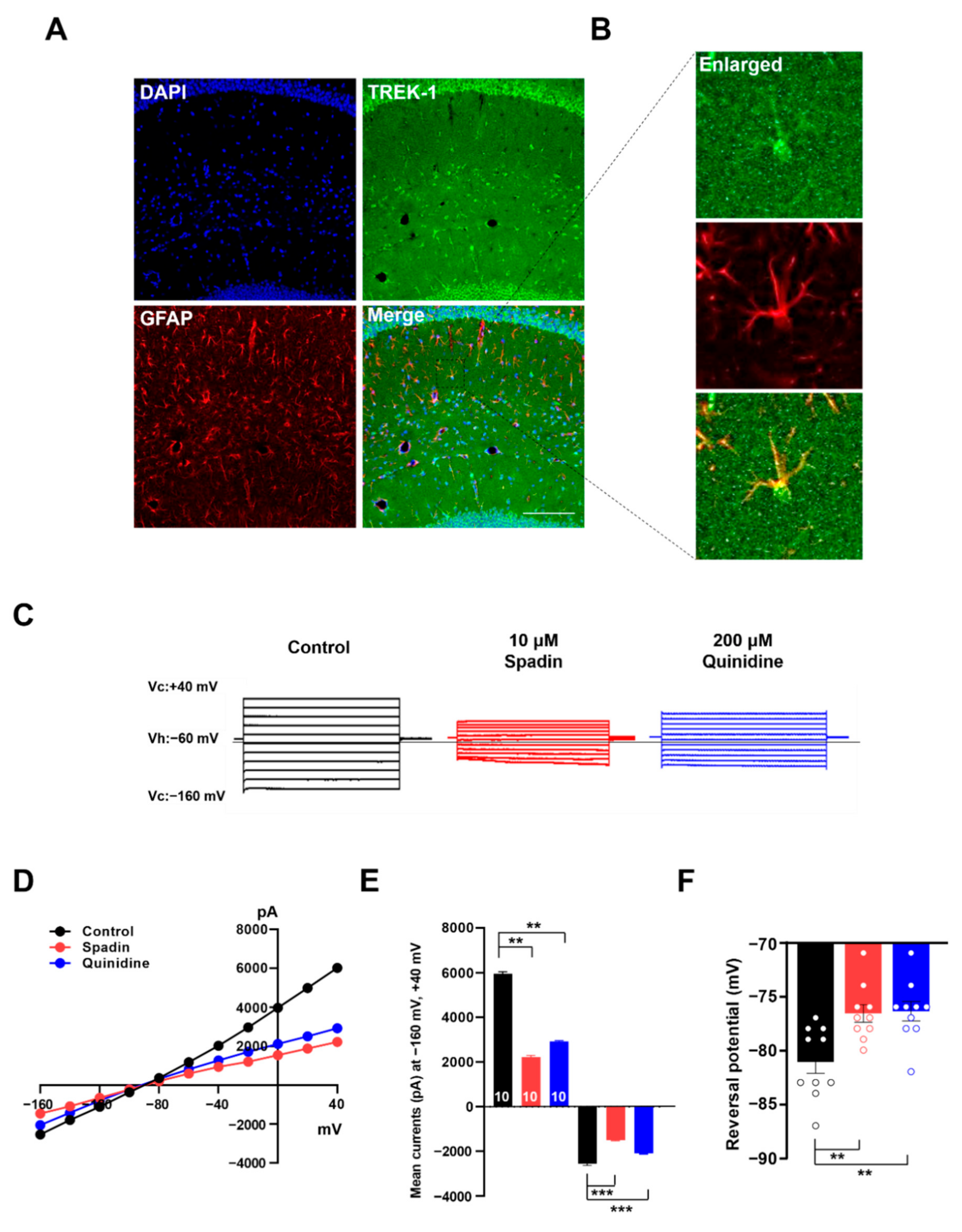
2.5. Spadin Inhibits Astrocytic Passive Conductance in Hippocampal Slices
2.6. Spadin Specifically Inhibits TWIK-1/TREK-1 Heterodimeric Channel-Mediated Astrocytic Passive Conductance
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Construction of Expression Vectors
4.3. Construction of shRNAs
4.4. Primary Cortical Astrocytes Culture and Transfection
4.5. COS-7 Cells Culture and Transfection
4.6. KO by CRISPR/Cas9 System
4.7. Immunocytochemistry
4.8. Electrophysiological Recording in Cultured Astrocytes
4.9. Electrophysiological Recording in COS-7 Cells
4.10. Immunohistochemistry
4.11. Lentivirus Construction and Electrophysiological Recording in Hippocampal Slices
4.12. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| RMP | Reversal membrane potential |
| 4-AP | 4-aminopyrimidine |
| TEA | Tetraethylammonium |
| shRNA | Short hairpin-forming interference RNA |
| CRISPR/Cas9 | Clustered regularly interspaced short palindromic repeats/CRISP-associated endonuclease 9 |
| sgRNA | Single guide RNA |
| AAV | Adeno-associated virus |
| AA | Arachidonic acid |
| K2P | Two-pore domain K+ channel |
| KO | Knockout |
| KD | Knockdown |
| TKDC | N-(4-cholorphenyl)-N-(2-(3,4-dihydrosioquinolin-2(1H)-yl)-2-oxoethyl)methanesulfonamide |
| ACSF | Artificial cerebrospinal fluid |
References
- Siracusa, R.; Fusco, R.; Cuzzocrea, S. Astrocytes: Role and functions in brain pathologies. Front. Pharmacol. 2019, 10, 1114. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.J. Astrocyte heterogeneity in the adult central nervous system. Front. Cell. Neurosci. 2018, 12, 401. [Google Scholar] [CrossRef] [Green Version]
- Vasile, F.; Dossi, E.; Rouach, N. Human astrocytes: Structure and functions in the healthy brain. Brain Struct. Funct. 2017, 222, 2017–2029. [Google Scholar] [CrossRef] [Green Version]
- Mederos, S.; González-Arias, C.; Perea, G. Astrocyte–neuron networks: A multilane highway of signaling for homeostatic brain function. Front. Synaptic Neurosci. 2018, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Du, Y.; Kiyoshi, C.M.; Ma, B.; Alford, C.C.; Wang, Q.; Yang, Y.; Liu, X.; Zhou, M. Electrophysiological behavior of neonatal astrocytes in hippocampal stratum radiatum. Mol. Brain 2016, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ryoo, K.; Park, J.-Y. Two-pore domain potassium channels in astrocytes. Exp. Neurobiol. 2016, 25, 222–232. [Google Scholar] [CrossRef]
- Steinhaüser, C.; Berger, T.; Frotscher, M.; Kettenmann, H. Heterogeneity in the membrane current pattern of identified glial cells in the hippocampal slice. Eur. J. Neurosci. 1992, 4, 472–484. [Google Scholar] [CrossRef]
- Kofuji, P.; Newman, E.A. Potassium buffering in the central nervous system. Neuroscience 2004, 129, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Bellot-Saez, A.; Kékesi, O.; Morley, J.W.; Buskila, Y. Astrocytic modulation of neuronal excitability through K+ spatial buffering. Neurosci. Biobehav. Rev. 2017, 77, 87–97. [Google Scholar] [CrossRef]
- Zhou, M.; Xu, G.; Xie, M.; Zhang, X.; Schools, G.P.; Ma, L.; Kimelberg, H.K.; Chen, H. TWIK-1 and TREK-1 are potassium channels contributing significantly to astrocyte passive conductance in rat hippocampal slices. J. Neurosci. 2009, 29, 8551–8564. [Google Scholar] [CrossRef]
- Hwang, E.M.; Kim, E.; Yarishkin, O.; Woo, D.H.; Han, K.-S.; Park, N.; Bae, Y.; Woo, J.; Kim, D.; Park, M. A disulphide-linked heterodimer of TWIK-1 and TREK-1 mediates passive conductance in astrocytes. Nat. Commun. 2014, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Kiyoshi, C.M.; Wang, Q.; Wang, W.; Ma, B.; Alford, C.C.; Zhong, S.; Wan, Q.; Chen, H.; Lloyd, E.E. Genetic deletion of TREK-1 or TWIK-1/TREK-1 potassium channels does not alter the basic electrophysiological properties of mature hippocampal astrocytes in situ. Front. Cell. Neurosci. 2016, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.J.; Honoré, E.; Maingret, F.; Lesage, F.; Fink, M.; Duprat, F.; Lazdunski, M. A mammalian two pore domain mechano-gated S-like K+ channel. EMBO J. 1998, 17, 4283–4290. [Google Scholar] [CrossRef]
- Maingret, F.; Patel, A.J.; Lesage, F.; Lazdunski, M.; Honoré, E. Mechano-or acid stimulation, two interactive modes of activation of the TREK-1 potassium channel. J. Biol. Chem. 1999, 274, 26691–26696. [Google Scholar] [CrossRef] [Green Version]
- Honoré, E.; Maingret, F.; Lazdunski, M.; Patel, A.J. An intracellular proton sensor commands lipid-and mechano-gating of the K+ channel TREK-1. EMBO J. 2002, 21, 2968–2976. [Google Scholar] [CrossRef] [Green Version]
- Chemin, J.; Patel, A.J.; Duprat, F.; Lauritzen, I.; Lazdunski, M.; Honoré, E. A phospholipid sensor controls mechanogating of the K+ channel TREK-1. EMBO J. 2005, 24, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Lopes, C.M.; Rohács, T.; Czirják, G.; Balla, T.; Enyedi, P.; Logothetis, D.E. PIP2 hydrolysis underlies agonist-induced inhibition and regulates voltage gating of two-pore domain K+ channels. J. Physiol. 2005, 564, 117–129. [Google Scholar] [CrossRef]
- Chemin, J.; Girard, C.; Duprat, F.; Lesage, F.; Romey, G.; Lazdunski, M. Mechanisms underlying excitatory effects of group I metabotropic glutamate receptors via inhibition of 2P domain K+ channels. EMBO J. 2003, 22, 5403–5411. [Google Scholar] [CrossRef] [Green Version]
- Murbartián, J.; Lei, Q.; Sando, J.J.; Bayliss, D.A. Sequential phosphorylation mediates receptor-and kinase-induced inhibition of TREK-1 background potassium channels. J. Biol. Chem. 2005, 280, 30175–30184. [Google Scholar] [CrossRef] [Green Version]
- Fink, M.; Duprat, F.; Lesage, F.; Reyes, R.; Romey, G.; Heurteaux, C.; Lazdunski, M. Cloning, functional expression and brain localization of a novel unconventional outward rectifier K+ channel. EMBO J. 1996, 15, 6854–6862. [Google Scholar] [CrossRef]
- Meadows, H.; Benham, C.; Cairns, W.; Gloger, I.; Jennings, C.; Medhurst, A.; Murdock, P.; Chapman, C. Cloning, localisation and functional expression of the human orthologue of the TREK-1 potassium channel. Pflügers Arch. 2000, 439, 714–722. [Google Scholar] [CrossRef]
- Mathie, A.; Veale, E.L. Therapeutic potential of neuronal two-pore domain potassium-channel modulators. Curr. Opin. Investig. Drugs 2007, 8, 555–562. [Google Scholar]
- Heurteaux, C.; Lucas, G.; Guy, N.; El Yacoubi, M.; Thümmler, S.; Peng, X.-D.; Noble, F.; Blondeau, N.; Widmann, C.; Borsotto, M. Deletion of the background potassium channel TREK-1 results in a depression-resistant phenotype. Nat. Neurosci. 2006, 9, 1134–1141. [Google Scholar] [CrossRef]
- Kennard, L.E.; Chumbley, J.R.; Ranatunga, K.M.; Armstrong, S.J.; Veale, E.L.; Mathie, A. Inhibition of the human two-pore domain potassium channel, TREK-1, by fluoxetine and its metabolite norfluoxetine. Br. J. Pharmacol. 2005, 144, 821. [Google Scholar] [CrossRef] [Green Version]
- Thümmler, S.; Duprat, F.; Lazdunski, M. Antipsychotics inhibit TREK but not TRAAK channels. Biochem. Biophys. Res. Commun. 2007, 354, 284–289. [Google Scholar] [CrossRef]
- Kim, E.-J.; Lee, D.K.; Hong, S.-G.; Han, J.; Kang, D. Activation of TREK-1, but Not TREK-2, channel by mood stabilizers. Int. J. Mol. Sci. 2017, 18, 2460. [Google Scholar]
- Mazella, J.; Pétrault, O.; Lucas, G.; Deval, E.; Béraud-Dufour, S.; Gandin, C.; El-Yacoubi, M.; Widmann, C.; Guyon, A.; Chevet, E. Spadin, a sortilin-derived peptide, targeting rodent TREK-1 channels: A new concept in the antidepressant drug design. PLoS Biol. 2010, 8, e1000355. [Google Scholar] [CrossRef] [Green Version]
- Ou Maati, H.M.; Veyssiere, J.; Labbal, F.; Coppola, T.; Gandin, C.; Widmann, C.; Mazella, J.; Heurteaux, C.; Borsotto, M. Spadin as a new antidepressant: Absence of TREK-1-related side effects. Neuropharmacology 2012, 62, 278–288. [Google Scholar] [CrossRef]
- Medhurst, A.D.; Rennie, G.; Chapman, C.G.; Meadows, H.; Duckworth, M.D.; Kelsell, R.E.; Gloger, I.I.; Pangalos, M.N. Distribution analysis of human two pore domain potassium channels in tissues of the central nervous system and periphery. Mol. Brain Res. 2001, 86, 101–114. [Google Scholar] [CrossRef]
- Talley, E.M.; Solórzano, G.; Lei, Q.; Kim, D.; Bayliss, D.A. CNS distribution of members of the two-pore-domain (KCNK) potassium channel family. J. Neurosci. 2001, 21, 7491–7505. [Google Scholar] [CrossRef]
- Woo, D.H.; Han, K.-S.; Shim, J.W.; Yoon, B.-E.; Kim, E.; Bae, J.Y.; Oh, S.-J.; Hwang, E.M.; Marmorstein, A.D.; Bae, Y.C. TREK-1 and Best1 channels mediate fast and slow glutamate release in astrocytes upon GPCR activation. Cell 2012, 151, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Echeverri, C.J.; Beachy, P.A.; Baum, B.; Boutros, M.; Buchholz, F.; Chanda, S.K.; Downward, J.; Ellenberg, J.; Fraser, A.G.; Hacohen, N. Minimizing the risk of reporting false positives in large-scale RNAi screens. Nat. Methods 2006, 3, 777–779. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Swiech, L.; Heidenreich, M.; Banerjee, A.; Habib, N.; Li, Y.; Trombetta, J.; Sur, M.; Zhang, F. In vivo interrogation of gene function in the mammalian brain using CRISPR-Cas9. Nat. Biotechnol. 2015, 33, 102–106. [Google Scholar] [CrossRef]
- Lengyel, M.; Czirják, G.; Enyedi, P. Formation of functional heterodimers by TREK-1 and TREK-2 two-pore domain potassium channel subunits. J. Biol. Chem. 2016, 291, 13649–13661. [Google Scholar] [CrossRef] [Green Version]
- Blin, S.; Soussia, I.B.; Kim, E.-J.; Brau, F.; Kang, D.; Lesage, F.; Bichet, D. Mixing and matching TREK/TRAAK subunits generate heterodimeric K2P channels with unique properties. Proc. Natl. Acad. Sci. USA 2016, 113, 4200–4205. [Google Scholar] [CrossRef] [Green Version]
- Rajan, S.; Plant, L.D.; Rabin, M.L.; Butler, M.H.; Goldstein, S.A. Sumoylation silences the plasma membrane leak K+ channel K2P1. Cell 2005, 121, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Kafitz, K.W.; Meier, S.D.; Stephan, J.; Rose, C.R. Developmental profile and properties of sulforhodamine 101—Labeled glial cells in acute brain slices of rat hippocampus. J. Neurosci. Methods 2008, 169, 84–92. [Google Scholar] [CrossRef]
- Wang, D.D.; Bordey, A. The astrocyte odyssey. Prog. Neurobiol. 2008, 86, 342–367. [Google Scholar] [CrossRef]
- Zhou, M.; Schools, G.P.; Kimelberg, H.K. Development of GLAST (+) astrocytes and NG2 (+) glia in rat hippocampus CA1: Mature astrocytes are electrophysiologically passive. J. Neurophysiol. 2006, 95, 134–143. [Google Scholar] [CrossRef] [Green Version]
- El-Brolosy, M.A.; Stainier, D.Y. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genet. 2017, 13, e1006780. [Google Scholar] [CrossRef] [Green Version]
- Ou Maati, H.M.; Peyronnet, R.; Devader, C.; Veyssiere, J.; Labbal, F.; Gandin, C.; Mazella, J.; Heurteaux, C.; Borsotto, M. A human TREK-1/HEK cell line: A highly efficient screening tool for drug development in neurological diseases. PLoS ONE 2011, 6, e25602. [Google Scholar] [CrossRef]
- Ma, R.; Lewis, A. Spadin selectively antagonizes arachidonic acid activation of TREK-1 channels. Front. Pharmacol. 2020, 11, 434. [Google Scholar] [CrossRef]
- Berg, A.P.; Talley, E.M.; Manger, J.P.; Bayliss, D.A. Motoneurons express heteromeric TWIK-related acid-sensitive K+ (TASK) channels containing TASK-1 (KCNK3) and TASK-3 (KCNK9) subunits. J. Neurosci. 2004, 24, 6693–6702. [Google Scholar] [CrossRef] [Green Version]
- Blin, S.; Chatelain, F.C.; Feliciangeli, S.; Kang, D.; Lesage, F.; Bichet, D. Tandem pore domain halothane-inhibited K+ channel subunits THIK1 and THIK2 assemble and form active channels. J. Biol. Chem. 2014, 289, 28202–28212. [Google Scholar] [CrossRef] [Green Version]
- Czirják, G.; Enyedi, P. Formation of functional heterodimers between the TASK-1 and TASK-3 two-pore domain potassium channel subunits. J. Biol. Chem. 2002, 277, 5426–5432. [Google Scholar] [CrossRef] [Green Version]
- Levitz, J.; Royal, P.; Comoglio, Y.; Wdziekonski, B.; Schaub, S.; Clemens, D.M.; Isacoff, E.Y.; Sandoz, G. Heterodimerization within the TREK channel subfamily produces a diverse family of highly regulated potassium channels. Proc. Natl. Acad. Sci. USA 2016, 113, 4194–4199. [Google Scholar] [CrossRef] [Green Version]
- Plant, L.D.; Zuniga, L.; Araki, D.; Marks, J.D.; Goldstein, S.A. SUMOylation silences heterodimeric TASK potassium channels containing K2P1 subunits in cerebellar granule neurons. Sci. Signal. 2012, 5, ra84. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.K.; Bhutani, M.K.; Bishnoi, M. Antidepressant activity of curcumin: Involvement of serotonin and dopamine system. Psychopharmacology 2008, 201, 435. [Google Scholar] [CrossRef]
- Ye, D.; Li, Y.; Zhang, X.; Guo, F.; Geng, L.; Zhang, Q.; Zhang, Z. TREK1 channel blockade induces an antidepressant-like response synergizing with 5-HT1A receptor signaling. Eur. Neuropsychopharmacol. 2015, 25, 2426–2436. [Google Scholar] [CrossRef]
- Luo, Q.; Chen, L.; Cheng, X.; Ma, Y.; Li, X.; Zhang, B.; Li, L.; Zhang, S.; Guo, F.; Li, Y. An allosteric ligand-binding site in the extracellular cap of K2P channels. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Moreno, S.; Devader, C.M.; Pietri, M.; Borsotto, M.; Heurteaux, C.; Mazella, J. Altered Trek-1 function in sortilin deficient mice results in decreased depressive-like behavior. Front. Pharmacol. 2018, 9, 863. [Google Scholar] [CrossRef]
- Kim, A.; Jung, H.-G.; Kim, Y.-E.; Kim, S.-C.; Park, J.-Y.; Lee, S.-G.; Hwang, E.M. The Knockdown of TREK-1 in Hippocampal Neurons Attenuate Lipopolysaccharide-Induced Depressive-Like Behavior in Mice. Int. J. Mol. Sci. 2019, 20, 5902. [Google Scholar] [CrossRef]
- Ventra, A.; Meissner, A.; Dillon, C.P.; McManus, M.; Sharp, P.A.; Parijs, L.V.; Jaenisch, R.; Jacks, T. Cre-lox-regulated conditional RNA interference from transgenes. Proc. Natl. Acad. Sci. USA 2004, 101, 10380. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, Y.; Choi, J.H.; Ryoo, K.; Kim, A.; Kwon, O.; Jung, H.-G.; Hwang, E.M.; Park, J.-Y. Spadin Modulates Astrocytic Passive Conductance via Inhibition of TWIK-1/TREK-1 Heterodimeric Channels. Int. J. Mol. Sci. 2020, 21, 9639. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249639
Bae Y, Choi JH, Ryoo K, Kim A, Kwon O, Jung H-G, Hwang EM, Park J-Y. Spadin Modulates Astrocytic Passive Conductance via Inhibition of TWIK-1/TREK-1 Heterodimeric Channels. International Journal of Molecular Sciences. 2020; 21(24):9639. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249639
Chicago/Turabian StyleBae, Yeonju, Jae Hyouk Choi, Kanghyun Ryoo, Ajung Kim, Osung Kwon, Hyun-Gug Jung, Eun Mi Hwang, and Jae-Yong Park. 2020. "Spadin Modulates Astrocytic Passive Conductance via Inhibition of TWIK-1/TREK-1 Heterodimeric Channels" International Journal of Molecular Sciences 21, no. 24: 9639. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249639