Methyl Jasmonate Induced Oxidative Stress and Accumulation of Secondary Metabolites in Plant Cell and Organ Cultures


Abstract
:1. Introduction
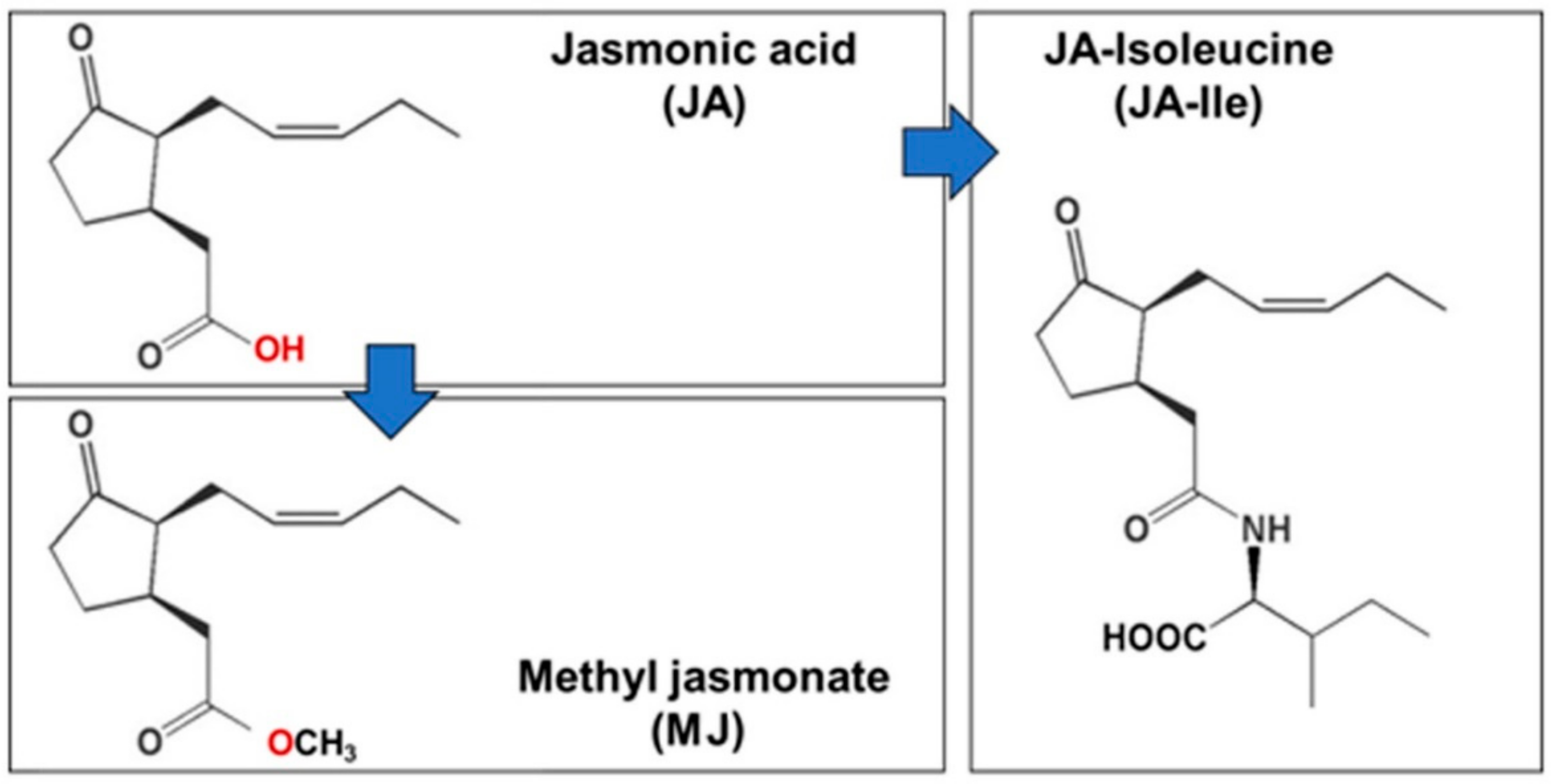
2. Jasmonic Acid (JA) and Methyl Jasmonate (MJ)
3. Usage of Methyl Jasmonate (MJ) on Oxidative Stress and Antioxidant Response
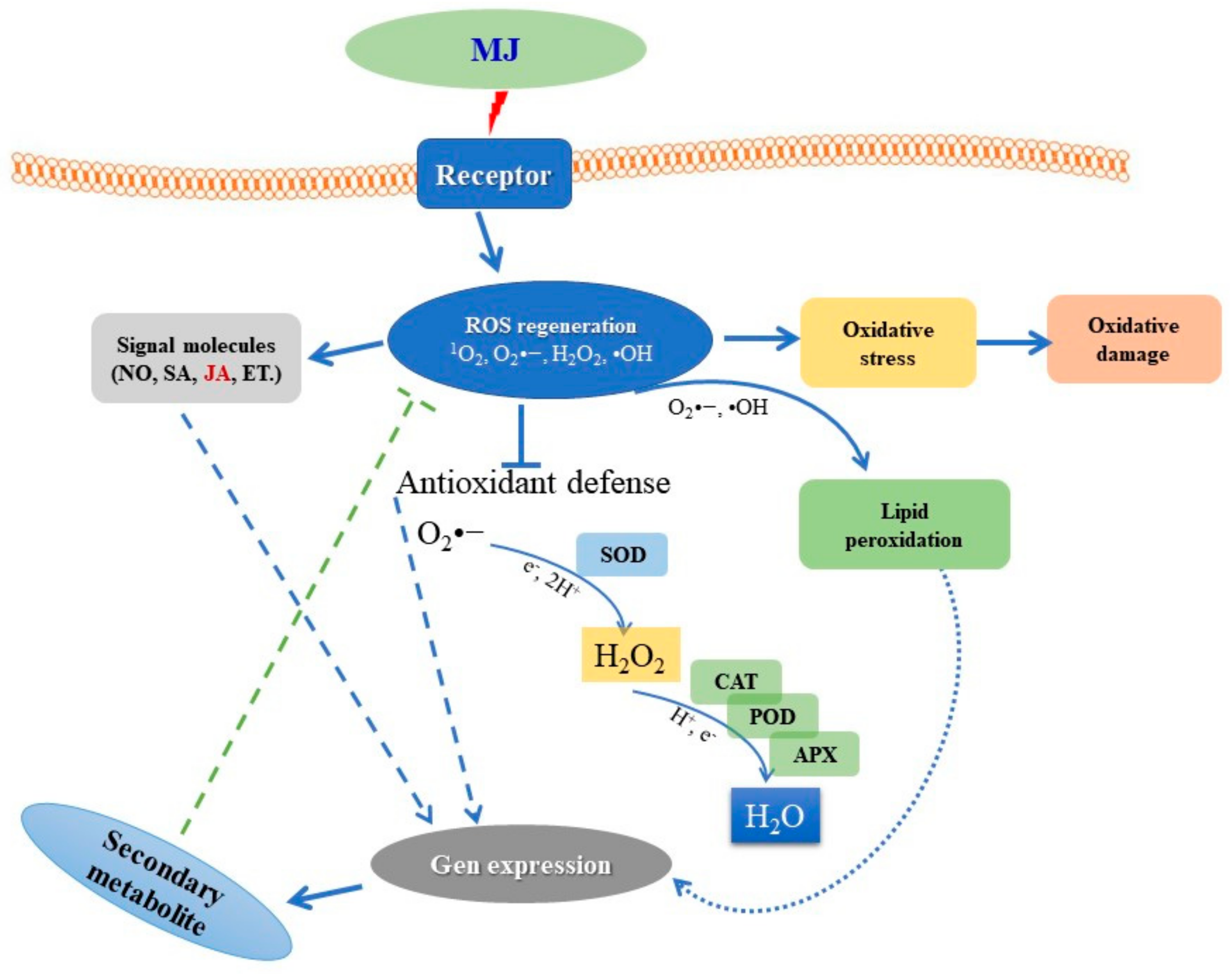
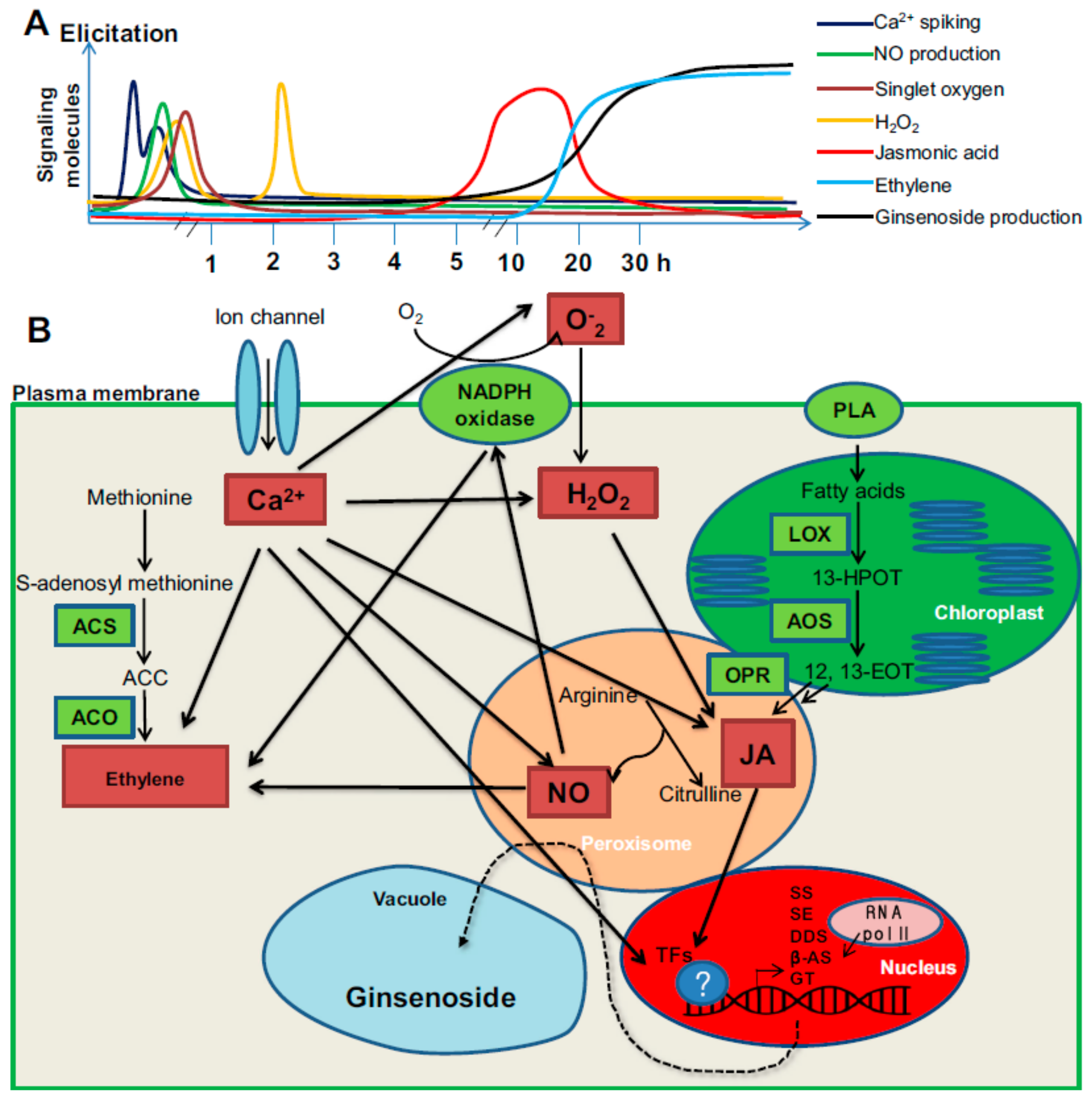
3.1. Summary of MJ Elicitation Mechanism
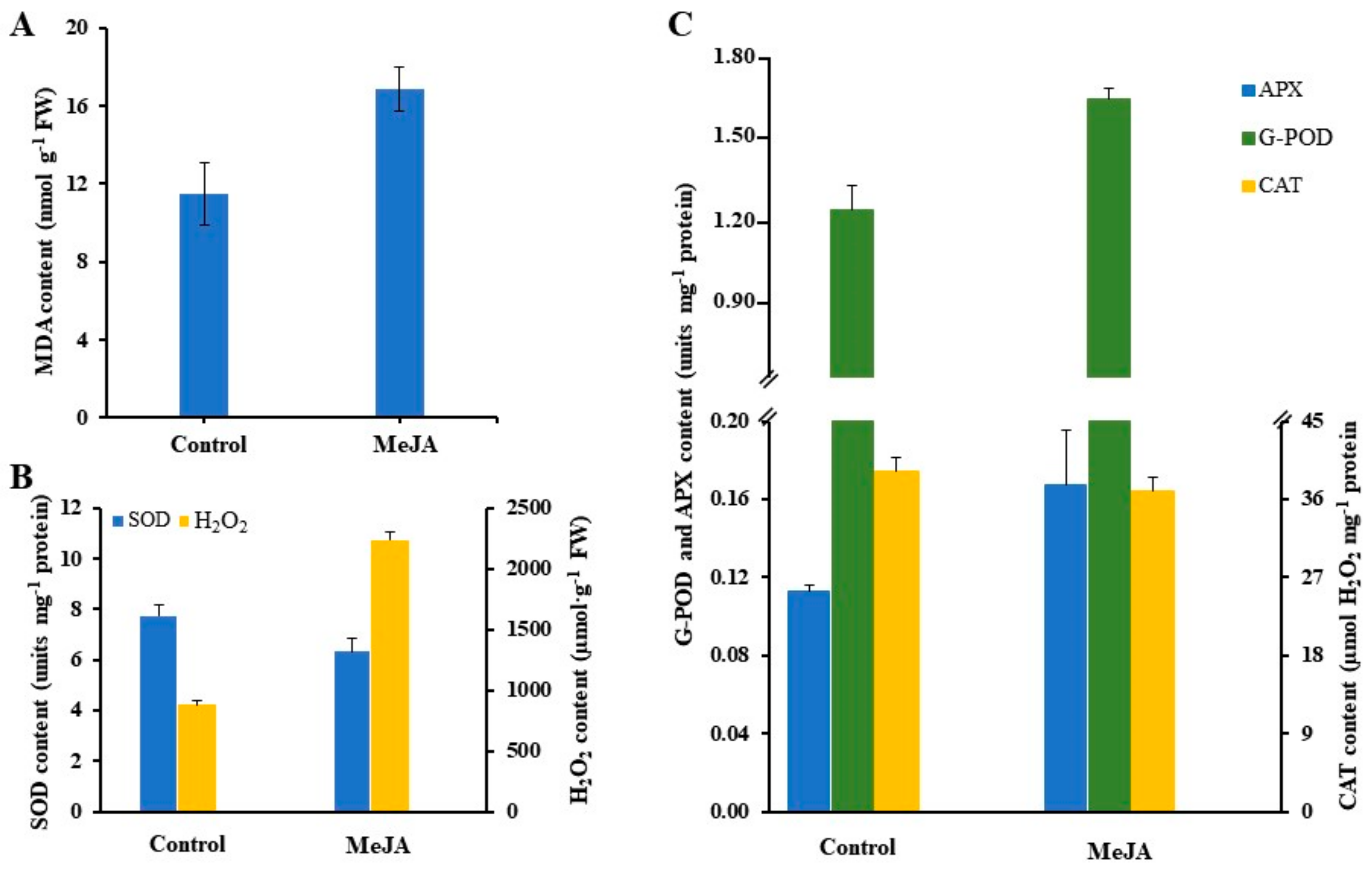
3.2. Oxidative Stress-Induced by Methyl Jasmonate
3.3. Antioxidant Response
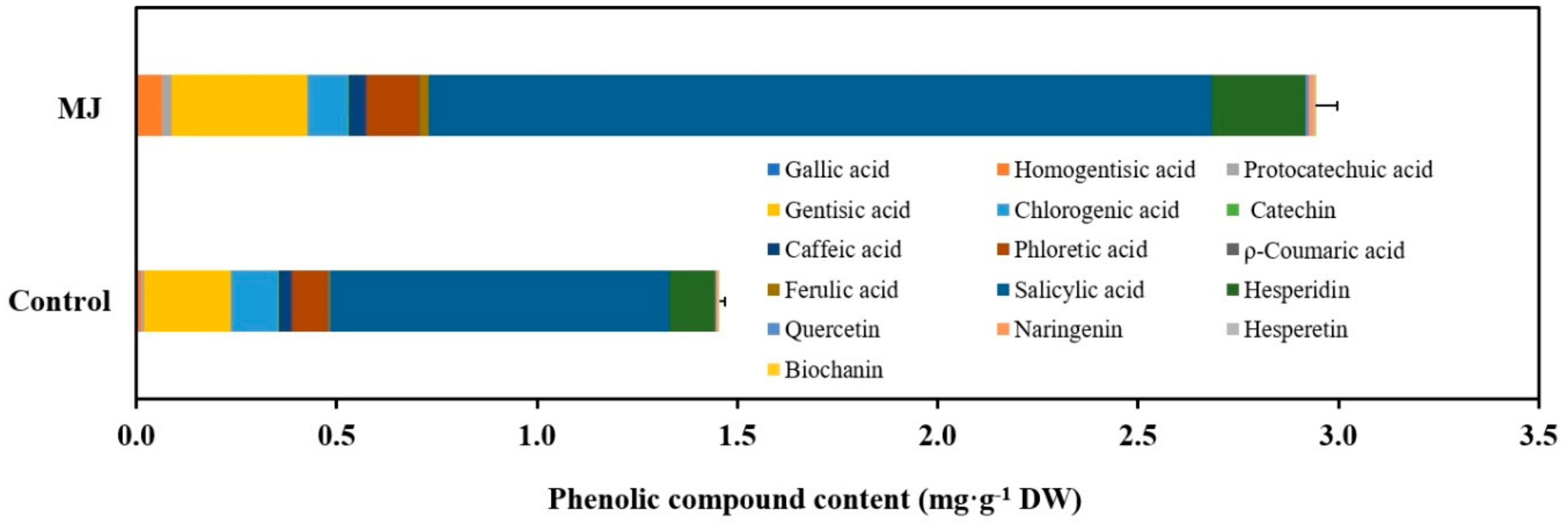
3.4. Signaling Pathway-Mediated Secondary Metabolite Accumulation in Plant Cell and Organ Cultures
4. Usage of Methyl Jasmonate (MJ) in Plant Cell and Organ Cultures
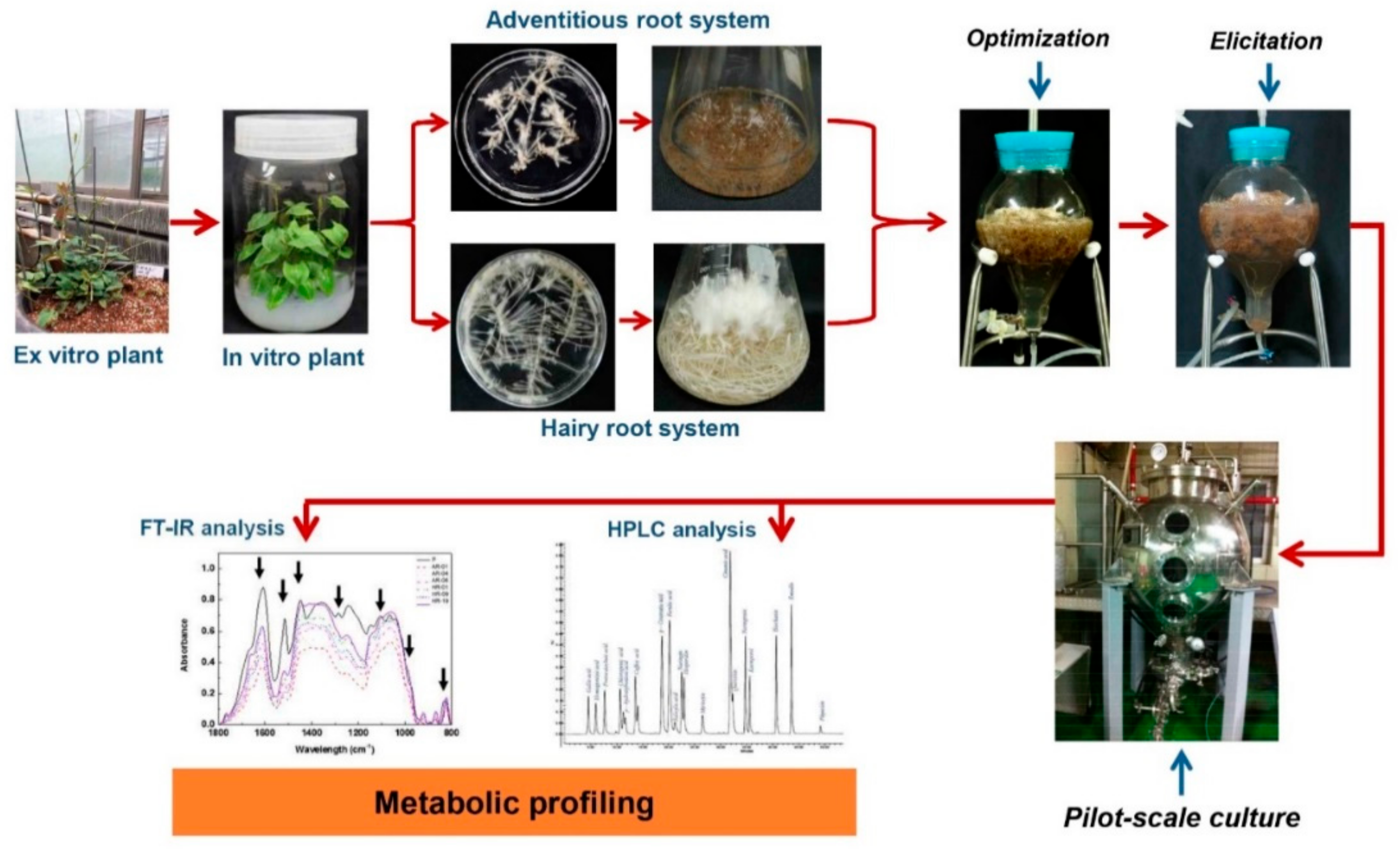
4.1. Application of MJ for Enhancing Secondary Metabolite in In Vitro Culture System
4.2. Application of MJ for Enhancement of Secondary Metabolites in Bioreactor Cultures
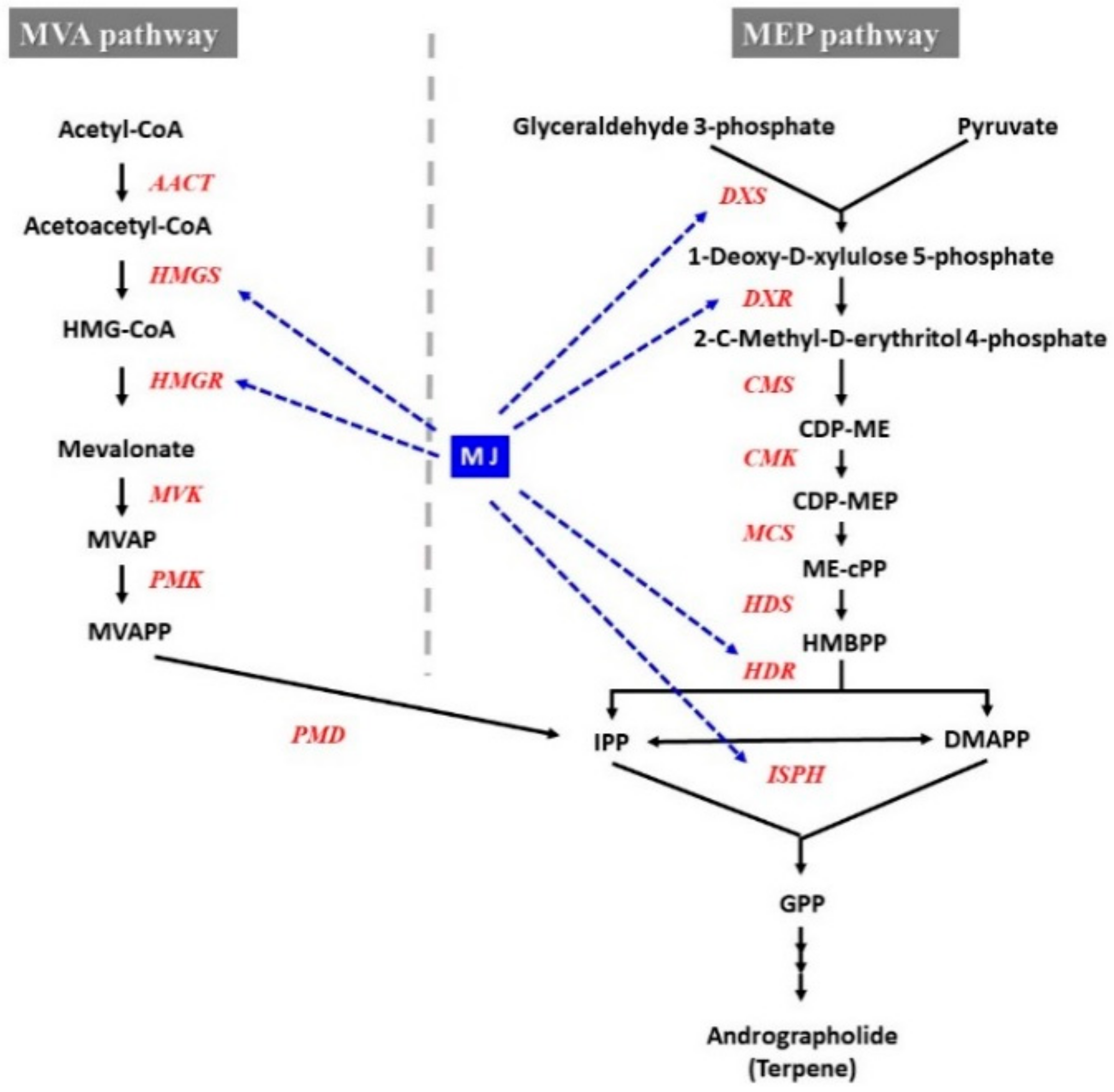
4.3. Effect of MJ on Gene Expression and Secondary Metabolite Accumulation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 4CL | Hydroxycinnamate coenzyme A ligase |
| AOC | Allene oxide cyclase |
| AOS | Allene oxide synthase |
| APX | Ascorbate peroxidase |
| ASA | Ascorbate |
| AsA-GSH | Ascorbateglutahione |
| C4H | Cinnamic acid 4-hydroxylase |
| CAT | Catalase |
| DHAR | Dehydroascorbate reductase |
| DXS | 1-Deoxy-d-xylulose 5-phosphate synthase |
| ET | Ethylene |
| FT-IR | Fourier-transform infrared spectroscopy |
| GGPPS | Geranylgeranyl diphosphate synthase |
| G-POD | Guaiacol peroxidase |
| GR | Glutathione reductase |
| GSH | Glutathione |
| H2O2 | Hydrogen peroxide |
| HDR | 1-Hydroxy-2-methyl-2-(E)-butenyl-4-diphosphate reductase |
| HPLC | High-performance liquid chromatography |
| HPPL | 4-Hydroxyphenylpyruvate reductase |
| ISPH | 1-Hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate reductase |
| JA | Jasmonic acid |
| JAs | Jasmonate |
| JA-Ile | (+)-7-iso-jasmonoyl-L-isoleucine |
| LOX | Lipoxygenase |
| MJ | Methyl jasmonate |
| MEP | Methylerythritol phosphate |
| MDHAR | Monodehydroascorbate reductase |
| MVA | Mevalonate |
| NO | Nitric oxide |
| PAL | Phenylalanine ammonia-lyase |
| RAS | Rosmarinic acid synthase |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| SA | Salicylic acid |
| SOD | Superoxide dismutase |
| TAT | Tyrosine aminotransferase |
References
- Ferrari, F.; Pasqua, G.; Monacelli, B. Xanthones from calli of Hypericum perforatum subsp. perforatum. Nat. Prod. Res. 2005, 19, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Brugger, G.A.; Lamotte, O.; Vandelle, E.; Bourque, S.; Lecourieux, D.; Poinssot, B.; Wendehenne, D.; Pugin, A. Early signalling events induced by elicitors of plant defences. Mol. Plant Microbe Interact. 2006, 19, 711–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paek, K.Y.; Murthy, H.N.; Hahn, E.J.; Zhong, J.J. Large scale culture of ginseng adventitious roots for production of ginsenosides. Adv. Biochem. Eng. Biotechnol. 2009, 113, 151–176. [Google Scholar]
- Murthy, H.N.; Lee, E.J.; Paek, K.Y. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tissue Organ Cult. 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Rahimi, S.; Kim, Y.J.; Yang, D.C. Production of ginseng saponins: Elicitation strategy and signal transductions. Appl. Microbiol. Biotechnol. 2015, 99, 6987–6996. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Harbinson, J. Oxygen metabolism and the regulation of photosynthetic electron transport. In Causes of Photooxidative Stresses and Amelioration of Defense Systems in Plants; Foyer, C.H., Mullineaux, P., Eds.; CRC Press: Boca Raton, FL, USA, 1994; pp. 1–42. [Google Scholar]
- Blokhina, O.; Fagerstedt, K.V. Reactive oxygen species and nitric oxide in plant mitochondria: Origin and redundant regulatory systems. Physiol. Plant. 2010, 138, 447–462. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Hu, W.H.; Song, X.S.; Shi, K.; Xia, X.J.; Zhou, Y.H.; Yu, J.Q. Changes in electron transport, superoxide dismutase and ascorbate peroxidase isoenzymes in chloroplasts and mitochondria of cucumber leaves as influenced by chilling. Photosynthetica 2008, 46, 581–588. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.; Mackerness, S.A.H.; Hancock, S.J.T.; Neill, S.J. Regulation of the Arabidopsis transcriptome by oxidative stress. Plant Physiol. 2001, 127, 9–172. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.B.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Differential responses of anti-oxidants enzymes, lipoxygenase activity, ascorbate content and the production of saponins in tissue cultured root of mountain Panax ginseng C.A. Mayer and Panax quinquefolium L. in bioreactor subjected to methyl jasmonate stress. Plant Sci. 2005, 169, 83–192. [Google Scholar]
- Ali, M.B.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate and salicylic acid elicitation induces ginsenosides accumulation, enzymatic and non-enzymatic antioxidant in suspension culture Panax ginseng roots in bioreactors. Plant Cell Rep. 2006, 25, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate and salicylic acid induced oxidative stress and accumulation of phenolics in Panax ginseng bioreactor root suspension culture. Molecules 2007, 12, 607–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, T.T. Strategies and Approaches for Improvement of Biomass and Bioactive Compounds in Adventitious Root and Hairy Root Cultures in Polygonum multiflorum. Ph.D. Thesis, Chungbuk National University, Cheongju, Korea, 2018. [Google Scholar]
- Ho, T.T.; Lee, K.J.; Lee, J.D.; Bhushan, S.; Paek, K.Y.; Park, S.Y. Adventitious root culture of Polygonum multiflorum for phenolic compounds and its pilot-scale production in 500 L-tank. Plant Cell Tissue Organ Cult. 2017, 130, 167–181. [Google Scholar] [CrossRef]
- Ho, T.T.; Murthy, H.N.; Dalawai, D.; Bhat, M.A.; Paek, K.Y.; Park, S.Y. Attributes of Polygonum multiflorum to transfigure red biotechnology. App. Microbiol. Biotechnol. 2019, 103, 3317–3326. [Google Scholar] [CrossRef] [PubMed]
- Demole, E.; Lederer, E.; Mercier, D. Isolement et détermination de la structure du jasmonate de méthyle, constituant odorant caractéristique de lessence de jasmin. Helv. Chim. Acta 1962, 45, 675–685. [Google Scholar] [CrossRef]
- Aldridge, D.C.; Galt, S.; Giles, D.; Turner, W.B. Metabolites of Lasiodiplodia theobromae. J. Chem. Soc. C 1971, 1623–1627. [Google Scholar] [CrossRef]
- Hyun, Y.; Choi, S.; Hwang, H.J.; Yu, J.; Nam, S.J.; Ko, J.; Park, J.Y.; Seo, Y.S.; Kim, E.Y.; Ryu, S.B.; et al. Cooperation and functional diversification of two closely related galactolipase genes for jasmonate biosynthesis. Dev. Cell 2008, 14, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Santino, A.; Taurino, M.; De Domenico, S.; Bonsegna, S.; Pastor, V.; Flors, V. Jasmonate signalling in plant development and defence response to multiple abiotic stresses. Plant Cell Tissue Organ Cult. 2013, 32, 1085–1098. [Google Scholar]
- Ziegler, J.; Stenzel, I.; Hause, B.; Mauche, H.; Hamberg, M.; Grimm, R.; Ganal, M.; Wasternack, C. Molecular cloning of allene oxide cyclase. The enzyme establishing the stereochemistry of octadecanoids and jasmonates. J. Biol. Chem. 2000, 275, 19132–19138. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Hause, B. Jasmonates and octadecanoids: Signal in plant stress responses and development. Prog. Nucl. Res. Mol. Biol. 2002, 72, 165–221. [Google Scholar]
- Yan, J.; Zhang, C.; Gu, M.; Bai, Z.; Zhang, W.; Qi, T.; Cheng, Z.; Peng, W.; Luo, H.; Nan, F.; et al. The Arabidopsis CORONATINE INSENSITIVE1 Protein is a jasmonate receptor. Plant Cell 2009, 21, 2220–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic acid signalling pathways in plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creelman, R.A.; Mullet, J.E. Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress. Proc. Natl. Acad. Sci. USA 1995, 92, 4114–4119. [Google Scholar] [CrossRef] [Green Version]
- Bertini, L.; Palazzi, L.; Proietti, S.; Pollastri, S.; Arrigoni, G.; de Laureto, P.; Carusco, C. Proteomic analysis of MeJa-induced defense responses in rice against wounding. Int. J. Mol. Sci. 2019, 2020, 2525. [Google Scholar] [CrossRef] [Green Version]
- Bertini, L.; Proietti, S.; Focaracci, F.; Sabatin, B.; Carusco, C. Epigenetic control of dense genes following MeJA-induced priming in rice (O. sativa). J. Plant Physiol. 2018, 228, 166–177. [Google Scholar]
- Giri, C.C.; Zaheer, M. Chemical elicitors versus secondary metabolite production in vitro using plant cell, tissue and organ cultures: Recent trends and a sky eye view appraisal. Plant Cell Tissue Organ Cult. 2016, 126, 1–18. [Google Scholar] [CrossRef]
- Baenas, N.; Garcia-Viguera, C.; Moreno, D.A. Elicitation: A tool for enriching the bioactive composition of foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Shohael, A.M.; Murthy, H.N.; Lee, H.L.; Hahn, E.J.; Paek, K.Y. Increased eleutheroside production in Eleutherococcus sessiliflorus embryogenic suspension cultures with methyl jasmonate treatment. Biochem. Eng. J. 2008, 38, 270–273. [Google Scholar] [CrossRef]
- Durango, D.; Pulgarin, N.; Echeverri, F.; Escobar, G.; Quiñones, W. Effect of salicylic acid and structurally related compounds in the accumulation of phytoalexins in cotyledons of common bean (Phaseolus vulgaris L.) cultivars. Molecules 2013, 18, 10609–10628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, T.T.; Lee, J.D.; Ahn, M.S.; Kim, S.W.; Park, S.Y. Enhanced production of phenolic compounds in hairy root cultures of Polygonum multiflorum and its metabolite discrimination using HPLC and FT-IR methods. Appl. Microbiol. Biotechnol. 2018, 102, 9563–9575. [Google Scholar] [CrossRef] [PubMed]
- Vasconsuelo, A.; Boland, R. Molecular aspects of the early stages of elicitation of secondary metabolites in plants. Plant Sci. 2007, 172, 861–875. [Google Scholar] [CrossRef]
- Mir, M.Y.; Hamid, S.; Kamili, A.N.; Hassan, Q.P. Sneak peek of Hypericum perforatum L.: Phytochemistry, phytochemical efficacy and biotechnological interventions. J. Plant Biochem. Biotechnol. 2019, 28, 357–375. [Google Scholar] [CrossRef]
- Scandalios, L.G. Oxygen stress and superoxide dismutase. Plant Physiol. 1993, 101, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.Q.; Cen, C.; Yang, M.L. Studies on the salicylic acid induced lipid peroxidation and defence gene expression in tabacco cell culture. Acta Bot. Sin. 1999, 41, 977–982. [Google Scholar]
- Chong, T.M.; Abdullah, M.A.; Fadzillah, N.M.; Lai, O.M.; Lajis, N.H. Jasmonic acid elicitation of anthraquinones with some associated. Enzyme Microb. Technol. 2005, 36, 469–477. [Google Scholar] [CrossRef]
- Creelman, R.A.; Mullet, J.E. Biosynthesis and action of jasmonates in plants. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 1997, 48, 355–381. [Google Scholar] [CrossRef] [Green Version]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Seo, Y. Antioxidant activity of the chemical constituents from the flower buds of Magnolia denudata. Biotechnol. Bioprocess Eng. 2010, 15, 400–406. [Google Scholar] [CrossRef]
- Parra-Lobato, M.G.; Fernandez-Garcia, N.; Olmos, E.; Alvarez-Tinaut, M.G.; Gomez-Jiménez, M.C. Methyl jasmonate-induced antioxidant defence in root apoplast from sunflower seedlings. Environ. Exp. Bot. 2009, 66, 9–17. [Google Scholar] [CrossRef]
- Wasternack, C.; Parthier, B. Jasmonate signaled plant gene expression. Trends Plant Sci. 1997, 2, 302–307. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Furutera, A.; Seki, K.; Toyoda, Y.; Tanaka, K.; Sugimoto, Y. Malondialdehyde generated from peroxidized linolenic acid causes protein modification in heat-stressed plants. Plant Physiol. Biochem. 2008, 8–9, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Welinder, K.G. Superfamily of plant, fungal and bacterial peroxidases. Curr. Opin. Struct. Biol. 1992, 2, 388–393. [Google Scholar] [CrossRef]
- Farooq, M.A.; Gill, R.A.; Islam, F.; Ali, B.; Liu, H.; Xu, J.; He, S.; Zhou, W. Methyl jasmonate regulates antioxidant defence and suppresses arsenic uptake in Brassica napus L. Front. Plant Sci. 2016, 7, 468. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Rahimi, S.; Devi, B.S.R.; Khorolragcha, A.; Kim, Y.J.; Kim, J.H.; Jung, S.K.; Yang, D.C. Effect of salicylic acid and yeast extract on the accumulation of jasmonic acid and sesquiterpenoids in Panax ginseng adventitious roots. Russ. J. Plant Physiol. 2014, 61, 811–817. [Google Scholar] [CrossRef]
- Kim, O.T.; Bang, K.H.; Kim, Y.C.; Hyun, D.Y.; Kim, M.Y.; Cha, S.W. Upregulation of ginsenoside and gene expression related to triterpene biosynthesis in ginseng hairy root cultures elicited by methyl jasmonate. Plant Cell Tissue Organ Cult. 2009, 98, 25–33. [Google Scholar] [CrossRef]
- Xie, D.X.; Feys, B.F.; James, S.; Nieto-Rostro, M.; Turner, J.G. COL1, an Arabidopsis gene required for jasmonae-regulated defense and fertility. Science 1998, 280, 1091–1094. [Google Scholar] [CrossRef]
- Staswick, P.E.; Tiryaki, I. The oxylipin signal jasmonic acid is activated by an enzyme that conjugates it it to isoleucine in Arabidopsis. Plan Cell 2004, 16, 2117–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, O.; Chico, J.M.; Sanchez-Serrano, J.J.; Solano, R. JASMONATE INSENSTIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The crosstalk between jasmonic acid and other plant hormone sginalling highlight the involvement of jasmonic acid as a core component in plant reponses to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, F.X.; Zhong, J.J. Jasmonic acid mediates gene transcription of ginsenoside biosynthesis in cell cultures of Panax notoginseng treated with chemically synthesized 2-hydroxyethyl jasmonate. Process Biochem. 2008, 43, 113–118. [Google Scholar] [CrossRef]
- Xu, X.; Hu, X.; Neill, S.J.; Fang, J.; Cai, W. Fungal elicitor induces singlet oxygen generation, ethylene release and saponin synthesis in cultured cells of Panax ginseng CA Meyer. Plant Cell Physiol. 2005, 46, 947–954. [Google Scholar] [CrossRef]
- Tewari, R.K.; Paek, K.Y. Salicylic acid-induced nitric oxide and ROS generation stimulate ginsenoside accumulation in Panax ginseng roots. J. Plant Growth Regul. 2011, 30, 396–404. [Google Scholar] [CrossRef]
- Jeong, G.T.; Park, D.H. Enhancement of growth and secondary metabolite biosynthesis: Effect of elicitor derived from plant and insects. Biotechnol. Bioprocess Eng. 2005, 10, 73–77. [Google Scholar] [CrossRef]
- Yin, Z.; Shangguan, X.; Chen, J.; Zhao, Q.; Li, D. Growth and triterpenic acid accumulation of Cyclocarya paliurus cell suspension cultures. Biotechnol. Bioprocess Eng. 2013, 18, 606–614. [Google Scholar] [CrossRef]
- Gorelick, J.; Bernstein, N. Elicitation: An underutilized tool in the development of medicinal plants as a source of therapeutic secondary metabolites. Adv. Agron. 2014, 124, 201–230. [Google Scholar]
- Ho, T.T.; Lee, J.D.; Jeong, C.S.; Paek, K.Y.; Park, S.Y. Improvement of biosynthesis and accumulation of bioactive compounds by elicitation in adventitious root cultures of Polygonum multiflorum. Appl. Microbiol. Biotechnol. 2018, 102, 199–209. [Google Scholar] [CrossRef]
- Belhadj, A.; Teleg, N.; Saigne, C.; Cluzet, S.; Barrieu, F.; Hamdi, S.; Merillon, J.M. Effect of methyl jasmonate in combination with carbohydrates on gene expression of PR proteins, stilbene and anthocyanin accumulation in grapevine cell cultures. Plant Physiol. Biochem. 2008, 46, 493–499. [Google Scholar] [CrossRef]
- Zhou, M.L.; Zhu, X.M.; Shao, J.R.; Tang, J.X.; Wu, Y.M. Production and metabolic engineering of bioactive substances in plant hairy root culture. Appl. Microbiol. Biotechnol. 2011, 90, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Han, R.B.; Yuan, Y.J. Oxidative burst in suspension culture of Taxus cuspidata induced by a laminar shear stress in short-term. Biotechnol. Prog. 2004, 20, 507–513. [Google Scholar] [CrossRef]
- Kim, Y.S.; Hahn, E.J.; Murthy, H.N.; Paek, K.Y. Adventitious root growth and ginsenoside accumulation in Panax ginseng cultures as affected by methyl jasmonate. Biotechnol. Lett. 2004, 26, 1619–1622. [Google Scholar] [CrossRef]
- Cai, J.; Ma, Y.; Hu, P.; Zhang, Y.; Chen, J.; Li, X. Elicitation of furanocoumarins in Changium smyrnioides suspension cells. Plant Cell Tissue Organ Cult. 2017, 130, 1–12. [Google Scholar] [CrossRef]
- Sampedro, M.A.S.; Tárrago, J.F.; Corchete, P. Yeast extract and methyl jasmonate-induced silymarin production in cell cultures of Silybum marianum (L.) Gaertn. J. Biotechnol. 2005, 119, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.E.; Ho, T.T.; Lee, H.S.; Jung, G.Y.; Kim, Y.E.; Jeong, C.S.; Park, S.Y. Enhanced biosynthesis of triterpenoids in Centella asiatica hairy root culture by precursor feeding and elicitation. Plant Biotechnol. Rep. 2019. [Google Scholar] [CrossRef]
- Shilpha, J.; Satish, L.; Kavikkuil, M.; Largia, M.J.V.; Ramesh, M. Methyl jasmonate elicits the solasodine production and antioxidant activity in hairy root cultures of Solanum trilobatum L. Ind. Crop. Prod. 2015, 71, 54–64. [Google Scholar] [CrossRef]
- Sivanandhan, G.; Dev, K.G.; Jeyaraj, M.; Rajesh, M.; Arjunan, A.; Muthuselvam, M.; Manickavasagam, M.; Ganapathi, A. Increased production of withanolide A, withanone and withaferin A in hairy root cultures of Withania somnifera (L.) Dunal elicited with methyl jasmonate and salicylic acid. Plant Cell Tissue Organ Cult. 2013, 114, 121–129. [Google Scholar] [CrossRef]
- Xing, B.; Yang, D.; Liu, L.; Han, R.; Sun, Y.; Liang, Z. Phenolic acid production is more effectively enhanced than tanshinone production by methyl jasmonate in Salvia miltiorrhiza hairy roots. Plant Cell Tissue Organ Cult. 2018, 134, 119–129. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, Z.J.; Xu, Y.; Qian, X.; Zhong, J.J. Efficient induction of ginsenoside biosynthesis and alteration of ginsenoside heterogeneity in cell cultures of Panax notoginseng by using chemically synthesized 2-hydroxyethyl jasmonate. Appl. Microbiol. Biotechnol. 2006, 70, 298–307. [Google Scholar] [CrossRef]
- Komaraiah, P.; Kavi-Kishor, P.B.; Carlsson, M.; Magnusson, K.E.; Mandenius, C.F. Enhancement of anthraquinone accumulation in Morinda citrifolia suspension cultures. Plant Sci. 2005, 168, 1337–1344. [Google Scholar] [CrossRef]
- Krzyzanowska, J.; Czubacka, A.; Pecio, L.; Przybys, M.; Doroszewska, T.; Stochmal, A.; Oleszek, W. The effects of jasmonic acid and methyl jasmonate on rosmarinic acid production in Mentha x piperita cell suspension cultures. Plant Cell Tissue Organ Cult. 2012, 108, 73–81. [Google Scholar] [CrossRef]
- Kang, S.M.; Jung, H.Y.; Kang, Y.M.; Yun, D.J.; Bahk, J.D.; Yang, J.K. Effects of methyl jasmonate and salicylic acid on the production of tropane alkaloids and the expression of PMT and H6H in adventitious root cultures of Scopolia praviflora. Plant Sci. 2004, 166, 745–751. [Google Scholar] [CrossRef]
- Zaker, A.; Sykora, C.; Gössnitzer, F.; Abrishamchi, P.; Asili, J.; Mousavi, S.H.; Wawrosch, C. Effects of some elicitors on tanshinone production in adventitious root cultures of Perovskia abrotanoides Karel. Ind. Crop. Prod. 2015, 67, 97–102. [Google Scholar] [CrossRef]
- Lee, E.J.; Park, S.Y.; Paek, K.Y. Enhancement strategies of bioactive compound production in adventitious root cultures of Eleutherococcus koreanum Nakai subjected to methyl jasmonate and salicylic acid elicitation through airlift bioreactors. Plant Cell Tissue Organ Cult. 2015, 120, 1–10. [Google Scholar] [CrossRef]
- Jiang, X.L.; Piao, X.C.; Gao, R.; Jin, M.Y.; Jiang, J.; Jin, X.H.; Lian, M.L. Improvement of bioactive compound accumulation in adventitious root cultures of an endangered plant species, Oplopanax elatus. Acta Physiol. Plant. 2017, 39, 226. [Google Scholar] [CrossRef]
- Thanh, N.T.; Murthy, H.N.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate elicitation enhanced synthesis of ginsenoside by cell suspension cultures of Panax ginseng in 5-l balloon type bubble bioreactor. Appl. Micorbiol. Biotechnol. 2005, 67, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shao, C.H.; Park, S.Y.; Piao, X.C.; Lian, M.L. Production of salidroside and polysaccharides in Rhodiola sachalinensis using airlift bioreactor systems. Acta Physiol. Plant. 2014, 36, 2975–2983. [Google Scholar] [CrossRef]
- Shohael, A.M.; Murthy, H.N.; Lee, H.L.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate induced overproduction of eleutherosides in somatic embryos of Eleutherococcus senticosus cultures in bioreactors. Electron. J. Biotechnol. 2007, 10, 633–637. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.A.; Wu, C.H.; Murthy, H.N.; Hahn, E.J.; Paek, K.Y. Application of an airlift bioreactor system for the production of adventitious root biomass and caffeic acid derivatives of Echinacea purpurea. Biotechnol. Bioprocess Eng. 2009, 14, 91–98. [Google Scholar] [CrossRef]
- Baque, M.A.; Moh, S.H.; Lee, E.J.; Zhong, J.J.; Paek, K.Y. Production of biomass and useful compounds from adventitious roots of high-value added medicinal plants using bioreactor. Biotechnol. Adv. 2012, 30, 1255–1267. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Dandin, V.S.; Paek, K.Y. Tools for biotechnological production of useful phytochemicals from adventitious root cultures. Phytochem. Rev. 2016, 15, 129–145. [Google Scholar] [CrossRef]
- Wang, G.R.; Qi, N.M. Influence of mist intervals and aeration rate on growth and second metabolite production of Pseudostellaria heterophylla adventitious roots in a siphon-mist bioreactor. Bioprocess Eng. 2010, 15, 1059–1098. [Google Scholar] [CrossRef]
- Cui, H.Y.; Baque, M.A.; Lee, E.J.; Paek, K.Y. Scale-up of adventitious root cultures of Echinacea angustifolia in a pilot-scale bioreactor for the production of biomass and caffeic acid derivatives. Plant. Biotechnol. Rep. 2013, 7, 297–308. [Google Scholar] [CrossRef]
- Baque, M.A.; Murthy, H.N.; Paek, K.Y. Adventitious root culture of Morinda citrifolia in bioreactors for production of bioactive compounds. In Production of Biomass and Bioactive Compounds Using Bioreactor Technology, 1st ed.; Paek, K.Y., Murthy, H.N., Zhong, J.J., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 185–222. [Google Scholar]
- Yu, H.; Liu, X.; Gao, S.; Han, X.; Cheng, A.; Lou, H. Molecular cloning and functional characterization of a phenylalanine ammonialyase from liverwort Plagiochasma appendiculatum. Plant Cell Tissue Organ Cult. 2014, 117, 265–277. [Google Scholar] [CrossRef]
- Mueller, M.J.; Brodschelm, W.; Spannagl, E.; Zenk, M.H. Signaling in the elicitation process is mediated through the octadecanoid pathway leading to jasmonic acid. Proc. Natl. Acad. Sci. USA 1993, 90, 7490–7494. [Google Scholar] [CrossRef] [Green Version]
- Sinha, R.K.; Sharma, S.N.; Verma, S.S.; Zha, J. Effects of lovastin, fosmidomycin and methyl jasmonate on andrographolide biosynthesis in the Andrographis paniculata. Acta Physiol. Plant. 2008, 40, 165. [Google Scholar] [CrossRef]
- Yan, Y.; Stolz, S.; Chetelat, A.; Reymond, P.; Pagni, M.; Dubugnon, L.; Farmer, E.E. A downstream mediator in the growth repression limb of the jasmonate pathway. Plant Cell 2007, 19, 2470–2483. [Google Scholar] [CrossRef] [Green Version]
- Murthy, H.N.; Georgiev, M.I.; Kim, Y.S.; Jeong, C.S.; Kim, S.J.; Park, S.Y.; Paek, K.Y. Ginsenosides: Prospective for sustainable biotechnological production. Appl. Microbiol. Biotechnol. 2014, 98, 6243–6254. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Material z | Culture System | MJ | Elicitor Duration | Biomass | Target Compound | Fold Increase Compound | Reference |
|---|---|---|---|---|---|---|---|---|
| Panax notoginseng | CS | Flask | 200 µM | 7 day | Not significant | Rb, Rg ginsenosides | 3.38 | [73] |
| Silybum marimum | CS | Flask | 100 µM | 3 day | Decreased | Silymarin | 4 | [68] |
| Morinda citrifolia | CS | Flask | 150 µM | 2 day | Slightly decreased | Anthraquinone | 4 | [74] |
| Mentha x piperita | CS | Flask | 100 µM | 2 day | Decreased | Rosmarinic acid | 1.8 | [75] |
| Changium smyrnioides | CS | Flask | 100 µM | 5 day | Decreased | Xanthotoxin, bergapten | 1.1 and 39.6 | [67] |
| Scopolia parviflora | AR | Flask | 1 mM | 1 day | Slightly decreased | Scopolamine and hyoscyamine | 2–3 | [76] |
| Perovskia abrotanoides | AR | Flask | 10 µM | 7 day | Not significant | Tanshinone | 2.3 | [77] |
| Polygonum multiflorum | AR | Flask | 50 µM | 7 day | Decreased | Phenolic | 2 | [62] |
| Withania somnifera | HR | Flask | 15 µM | 4 h | Slightly decreased | Withanolide derivatives | 14 | [71] |
| Solanum trilobatum | HR | Flask | 4 µM | 14 day | Slightly decreased | Solasodine | 6.5 | [70] |
| Salvia miltiorrhiza | HR | Flask | 100 µM | 6 day | Decreased | Phenolic and tanshinones | 3.3 and 2.5 | [71] |
| Polygonum multiflorum | HR | Flask | 50 µM | 5 day | Slightly decreased | Phenolic | 3.2 | [34] |
| Echinacea purpurea | HR | Flask/Bioreactor | 400 µM | 7 day | Not significant | Triterpenoid | 4.5 | [78] |
| Polygonum multiflorum | AR | Bioreactor | 50 µM | 7 day | Slightly decreased | Phenolic | 2.1 | [62] |
| Panax ginseng | AR | Bioreactor | 100 µM | 10 day | Decreased | Ginsenoside | 11-fold in Rb group | [66] |
| Panax ginseng and Panax quinquefolium | AR | Bioreactor | 200 µM | 9 day | Not significant | Saponin | 4 | [12,13] |
| Eleutherococcus koreanum | AR | Bioreactor | 50 µM | 7 day | Not significant | Eleutherosides B and E, and chlorogenic acid | 1.1, 1.4 and 1.2 | [79] |
| Olopanax elatus | AR | Bioreactor | 200 µM | 8 day | Decreased | Polysaccharide, phenolic | 1.83 and 1.49 | [80] |
| Panax ginseng | CS | Bioreactor | 200 µM | 8 day | Decreased | Ginsenoside | 1.6–3.7 | [81] |
| Rhodiola sachalinensis | CS | Bioreactor | 125 µM | 7 day | Decreased | Salidroside | 5 | [82] |
| Eleutherococcus senticosus | SE | Bioreactor | 200 µM | 7 day | Decreased | Eleutheroside | 7.3 | [83] |
| Systems | FW (g·L−1) | DW (g·L−1) | Dry Matter (%) | Growth Ratio z | Total Bioactive Compounds (mg·g−1 DW) y | Total Productivity (mg·L−1) x |
|---|---|---|---|---|---|---|
| Polygonum multiflorumw | ||||||
| Control | 93.95 ± 3.23 | 10.61 ± 0.32 | 11.30 ± 0.11 | 20.23 ± 0.64 | 11.20 ± 0.17 | 118.82 ± 1.78 |
| MJ50 | 90.09 ± 1.51 | 9.99 ± 0.23 | 11.10 ± 0.44 | 18.98 ± 0.46 | 22.83 ± 0.30 | 228.08 ± 3.00 |
| 5-year-old root | 24.78 ± 0.82 | |||||
| Echinacea purpureav | ||||||
| Control | 83.81 ± 2.12 | 8.10 ± 0.72 | 9.63 ± 0.64 | 15.19 ± 1.45 | 25.22 ± 1.27 | 204.16 ± 10.24 |
| MJ200 | 67.26 ± 1.72 | 6.24 ± 0.40 | 9.27 ± 0.53 | 11.48 ± 0.80 | 38.32 ± 0.34 | 239.20 ± 4.44 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, T.-T.; Murthy, H.N.; Park, S.-Y. Methyl Jasmonate Induced Oxidative Stress and Accumulation of Secondary Metabolites in Plant Cell and Organ Cultures. Int. J. Mol. Sci. 2020, 21, 716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030716
Ho T-T, Murthy HN, Park S-Y. Methyl Jasmonate Induced Oxidative Stress and Accumulation of Secondary Metabolites in Plant Cell and Organ Cultures. International Journal of Molecular Sciences. 2020; 21(3):716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030716
Chicago/Turabian StyleHo, Thanh-Tam, Hosakatte Niranjana Murthy, and So-Young Park. 2020. "Methyl Jasmonate Induced Oxidative Stress and Accumulation of Secondary Metabolites in Plant Cell and Organ Cultures" International Journal of Molecular Sciences 21, no. 3: 716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030716