Regulation of Small GTPase Rab20 by Ikaros in B-Cell Acute Lymphoblastic Leukemia

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
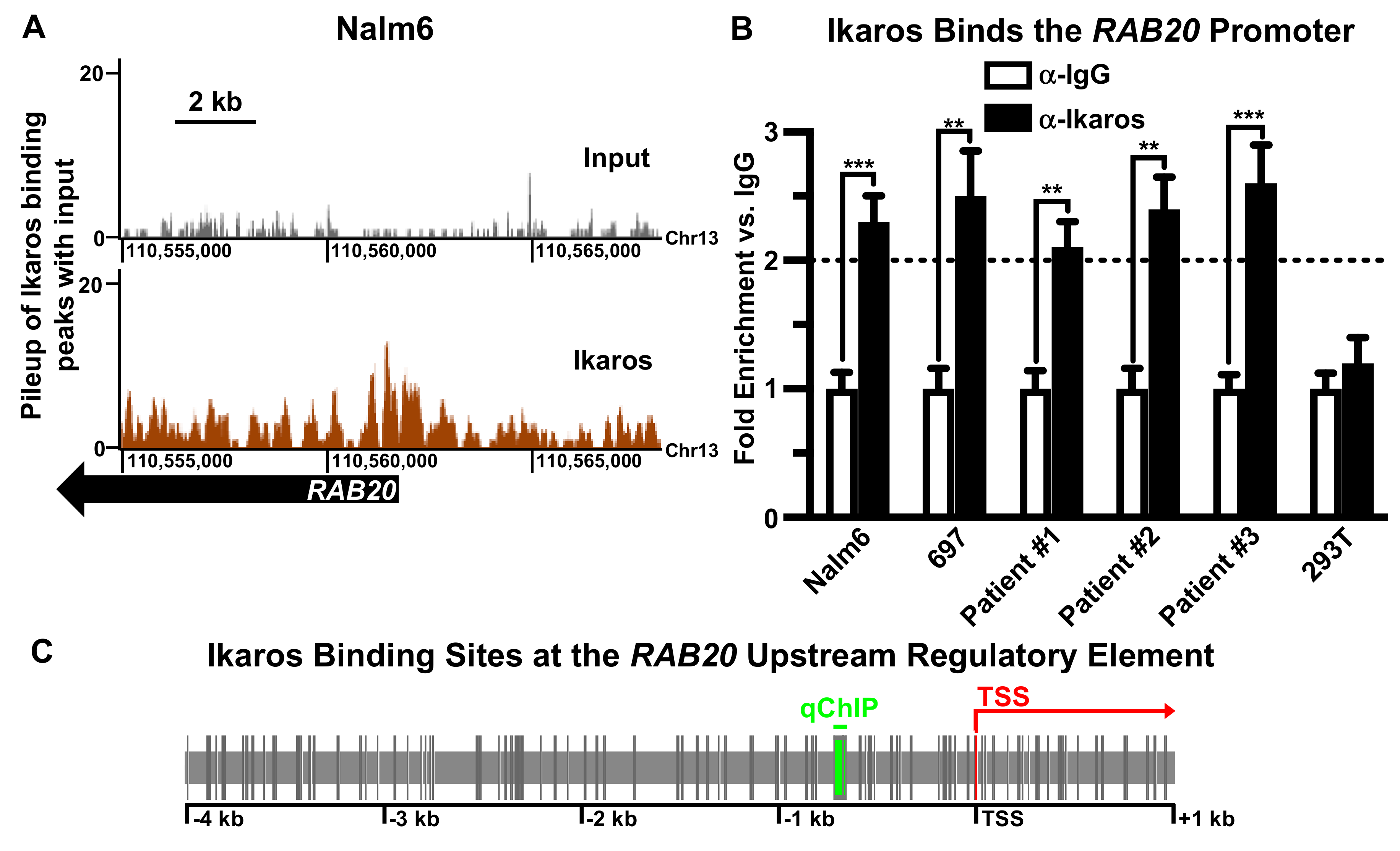
2.1. Ikaros Binds to the RAB20 Promoter in B-ALL Cells
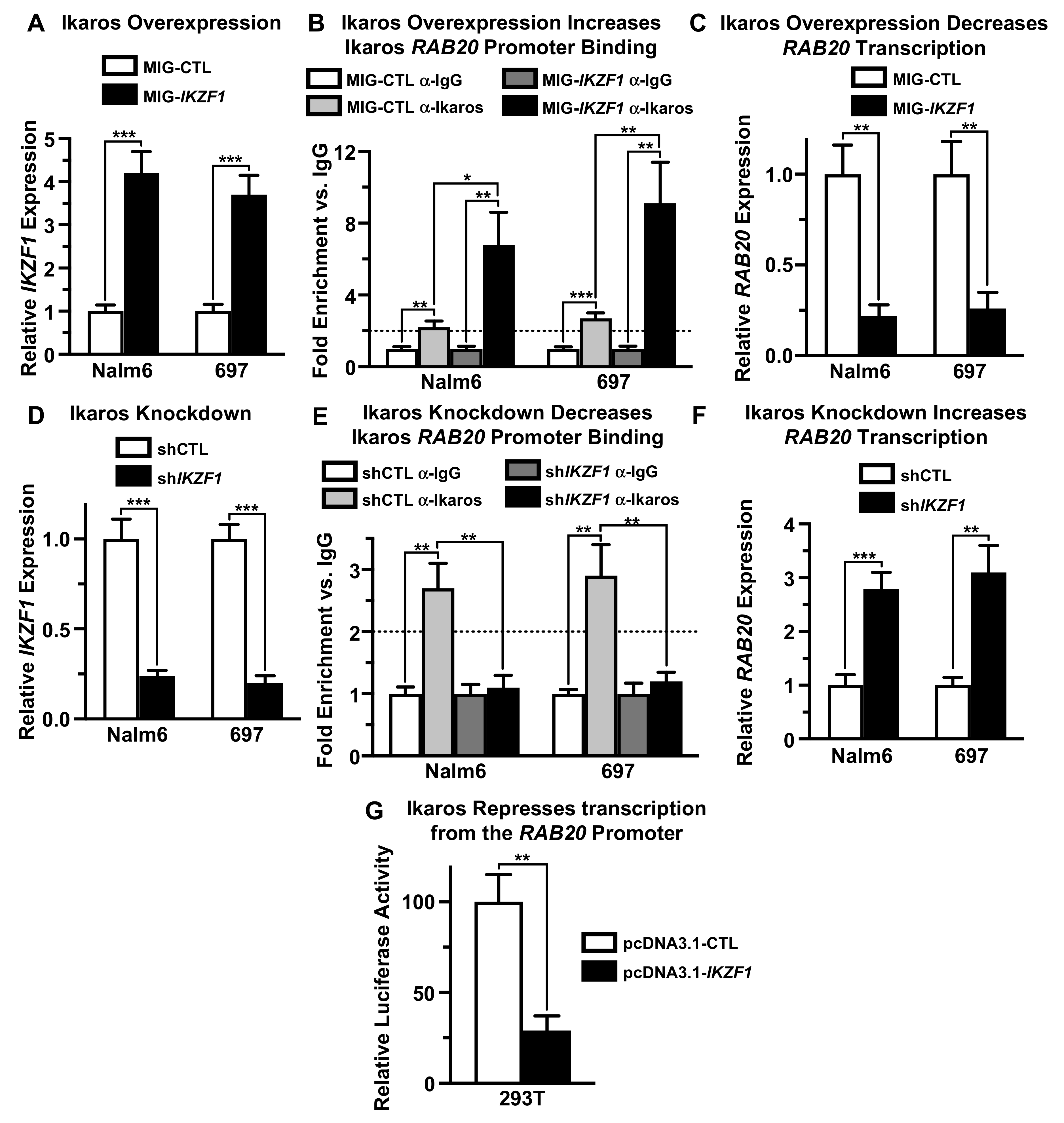
2.2. Ikaros Represses Transcription of the RAB20 Gene
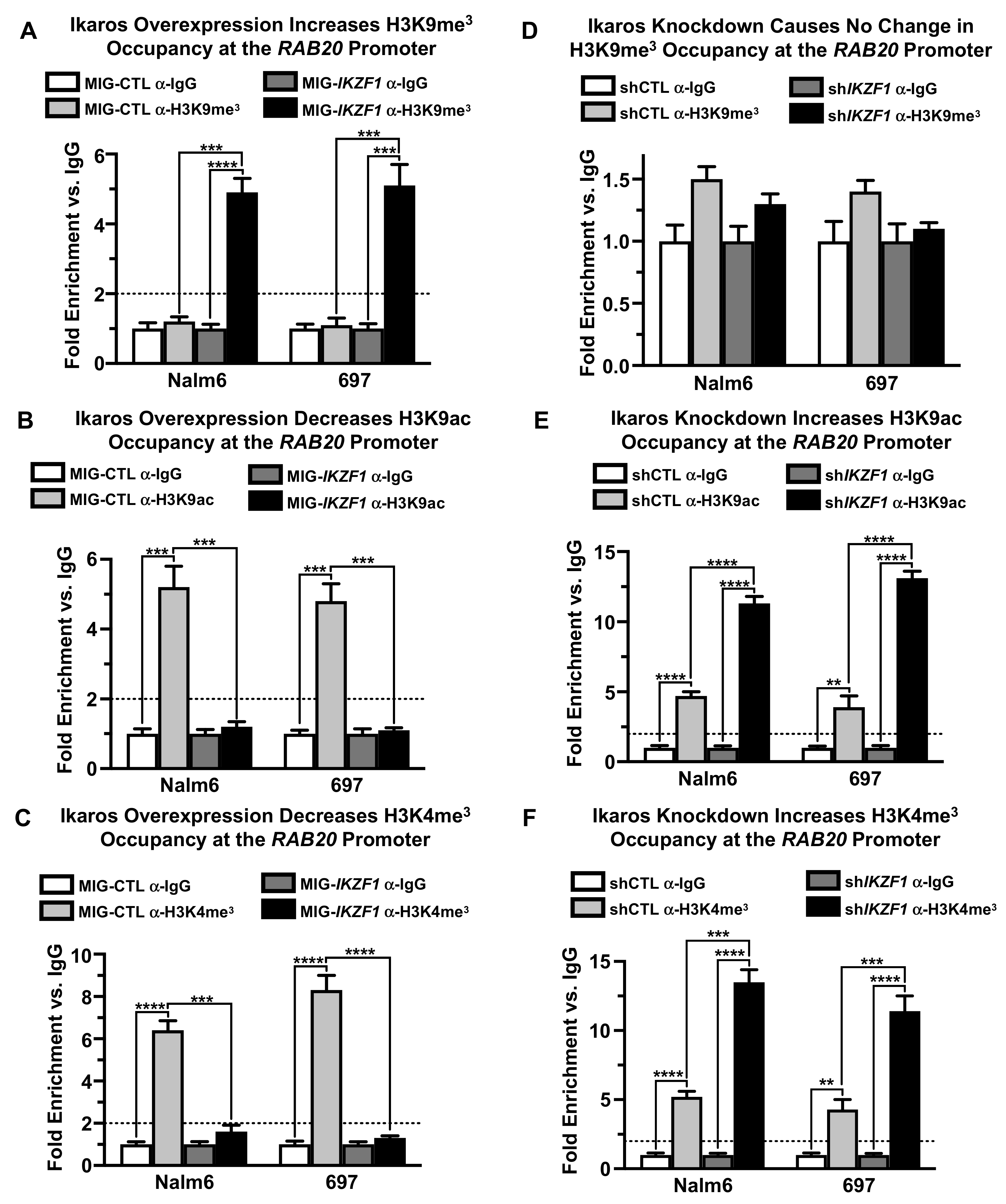
2.3. Ikaros Represses RAB20 Transcription via Chromatin Remodeling
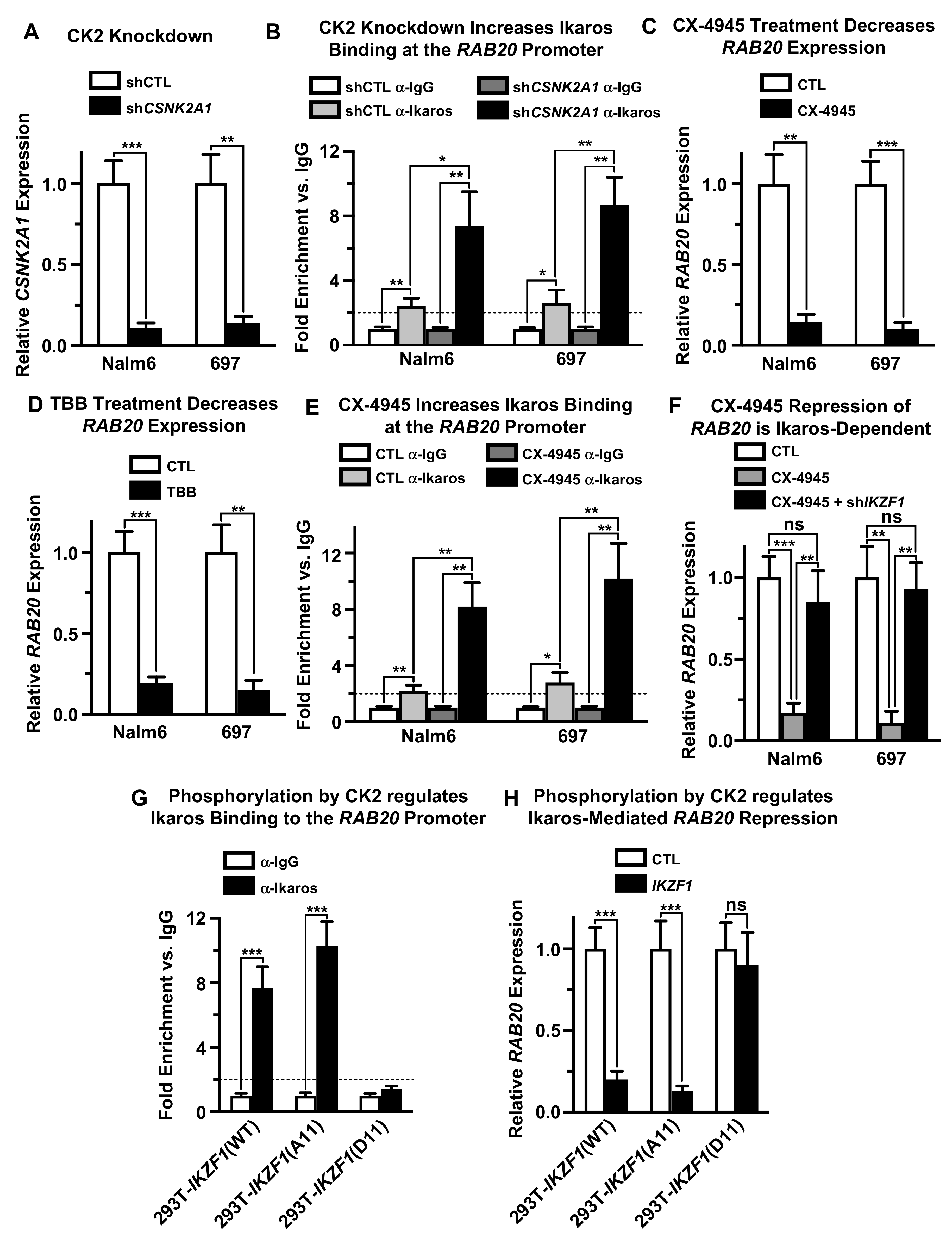
2.4. Casein Kinase II (CK2) Regulates Ikaros’ Ability to Repress RAB20 Transcription
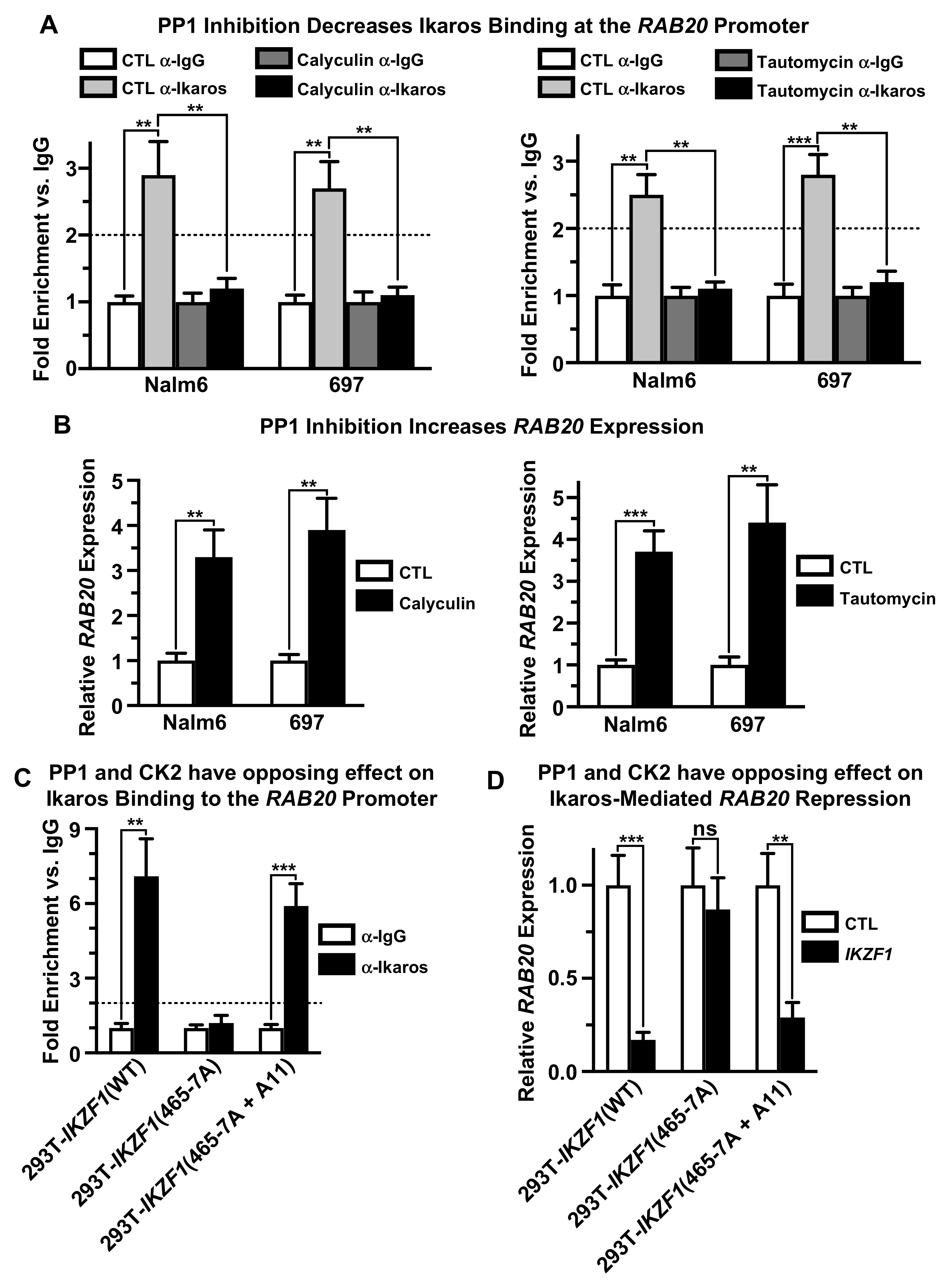
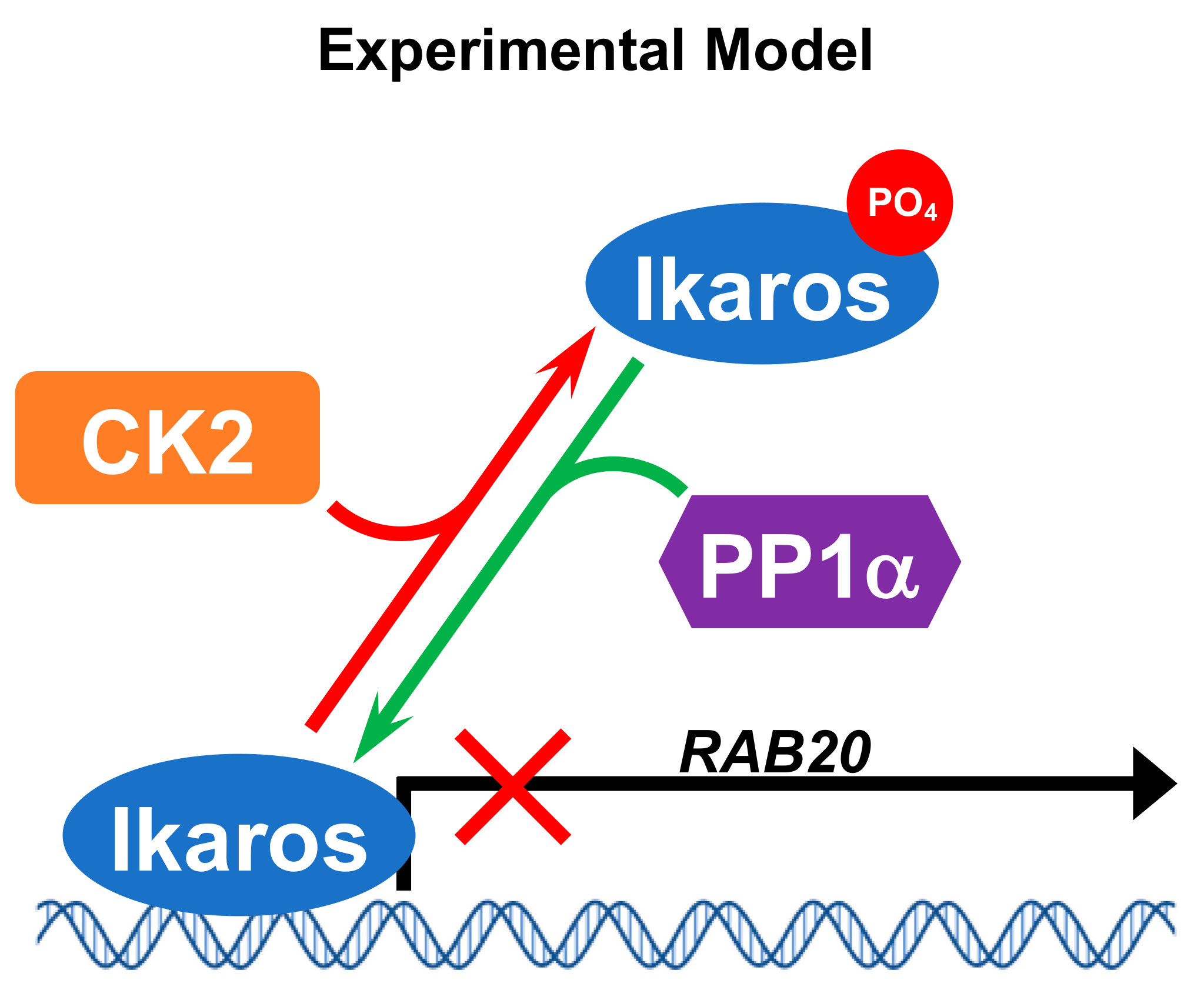
2.5. Protein Phosphatase 1 (PP1) Regulates Ikaros’ Function as Repressor of RAB20 Transcription
3. Discussion
4. Materials and Methods
4.1. Cells and Cell Culture
4.2. Metanalyses
4.3. Reagents
4.4. CaPO4 Transfection and 293T Gene Expression
4.5. Expression Plasmids
4.6. Luciferase Reporter Assay
4.7. Retroviral Transduction
4.8. Ikaros and CK2 shRNA Knockdown
4.9. Quantitative RT-PCR (qRT-PCR)
4.10. qChIP
4.11. ChIP-Seq Experiments
4.12. Antibodies
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| B-ALL | B-Cell Acute Lymphoblastic Leukemia |
| ChIP-seq | Chromatin Immunoprecipitation and Sequencing |
| GEO | Gene Expression Omnibus |
| PDX | Patient-Derived Xenograft |
| qChIP | Quantitative Chromatin Immunoprecipitation |
| qRT-PCR | Quantitative Reverse Transcription Polymerase Chain Reaction |
| TSS | Transcription Start Site |
References
- Georgopoulos, K.; Moore, D.D.; Derfler, B. Ikaros, an early lymphoid-specific transcription factor and a putative mediator for T cell commitment. Science 1992, 258, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.; Landau, N.R.; Smale, S.T. LyF-1, a transcriptional regulator that interactswith a novel class of promoters for lymphocyte-specific genes. Mollecular Cellular Biology 1991, 11, 5229–5243. [Google Scholar]
- Molnár, A.; Georgopoulos, K. The Ikaros gene encodes a family of functionally diverse zinc finger DNA-binding proteins. Mol. Cell Biol 1994, 14, 8292–8303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahm, K.; Ernst, P.; Lo, K.; Kim, G.S.; Turck, C.; Smale, S.T. The lymphoid transcription factor LyF-1 is encoded by specific, alternatively spliced mRNAs derived from the Ikaros gene. Mol. Cell Biol 1994, 14, 7111–7123. [Google Scholar] [CrossRef] [Green Version]
- Georgopoulos, K.; Bigby, M.; Wang, J.H.; Molnar, A.; Wu, P.; Winandy, S.; Sharpe, A. The Ikaros gene is required for the development of all lymphoid lineages. Cell 1994, 79, 143–156. [Google Scholar] [CrossRef]
- Wang, J.H.; Nichogiannopoulou, A.; Wu, L.; Sun, L.; Sharpe, A.H.; Bigby, M.; Georgopoulos, K. Selective defects in the development of the fetal and adult lymphoid system in mice with an Ikaros null mutation. Immunity 1996, 5, 537–549. [Google Scholar] [CrossRef] [Green Version]
- Sigvardsson, M. Molecular Regulation of Differentiation in Early B-Lymphocyte Development. Int J. Mol. Sci 2018, 19, 1928. [Google Scholar] [CrossRef] [Green Version]
- Winandy, S.; Wu, P.; Georgopoulos, K. A dominant mutation in the Ikaros gene leads to rapid development of leukemia and lymphoma. Cell 1995, 83, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Mullighan, C.G.; Goorha, S.; Radtke, I.; Miller, C.B.; Coustan-Smith, E.; Dalton, J.D.; Girtman, K.; Mathew, S.; Ma, J.; Pounds, S.B.; et al. Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia. Nature 2007, 446, 758–764. [Google Scholar] [CrossRef]
- Mullighan, C.; Downing, J. Ikaros and acute leukemia. Leuk Lymphoma 2008, 49, 847–849. [Google Scholar] [CrossRef]
- Mullighan, C.G.; Miller, C.B.; Radtke, I.; Phillips, L.A.; Dalton, J.; Ma, J.; White, D.; Hughes, T.P.; Le Beau, M.M.; Pui, C.H.; et al. BCR-ABL1 lymphoblastic leukaemia is characterized by the deletion of Ikaros. Nature 2008, 453, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, R.P.; Schoenmakers, E.F.; van Reijmersdal, S.V.; Hehir-Kwa, J.Y.; van Kessel, A.G.; van Leeuwen, F.N.; Hoogerbrugge, P.M. High-resolution genomic profiling of childhood ALL reveals novel recurrent genetic lesions affecting pathways involved in lymphocyte differentiation and cell cycle progression. Leukemia 2007, 21, 1258–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacobucci, I.; Lonetti, A.; Cilloni, D.; Messa, F.; Ferrari, A.; Zuntini, R.; Ferrari, S.; Ottaviani, E.; Arruga, F.; Paolini, S.; et al. Identification of different Ikaros cDNA transcripts in Philadelphia-positive adult acute lymphoblastic leukemia by a high-throughput capillary electrophoresis sizing method. Haematologica 2008, 93, 1814–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacobucci, I.; Storlazzi, C.T.; Cilloni, D.; Lonetti, A.; Ottaviani, E.; Soverini, S.; Astolfi, A.; Chiaretti, S.; Vitale, A.; Messa, F.; et al. Identification and molecular characterization of recurrent genomic deletions on 7p12 in the IKZF1 gene in a large cohort of BCR-ABL1-positive acute lymphoblastic leukemia patients: on behalf of Gruppo Italiano Malattie Ematologiche dell’Adulto Acute Leukemia Working Party (GIMEMA AL WP). Blood 2009, 114, 2159–2167. [Google Scholar]
- Coccaro, N.; Anelli, L.; Zagaria, A.; Specchia, G.; Albano, F. Next-Generation Sequencing in Acute Lymphoblastic Leukemia. Int J. Mol. Sci 2019, 20, 2929. [Google Scholar] [CrossRef] [Green Version]
- Mata-Rocha, M.; Rangel-Lopez, A.; Jimenez-Hernandez, E.; Morales-Castillo, B.A.; Gonzalez-Torres, C.; Gaytan-Cervantes, J.; Alvarez-Olmos, E.; Nunez-Enriquez, J.C.; Fajardo-Gutierrez, A.; Martin-Trejo, J.A.; et al. Identification and Characterization of Novel Fusion Genes with Potential Clinical Applications in Mexican Children with Acute Lymphoblastic Leukemia. Int J. Mol. Sci 2019, 20, 2394. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ding, L.; Holmfeldt, L.; Wu, G.; Heatley, S.L.; Payne-Turner, D.; Easton, J.; Chen, X.; Wang, J.; Rusch, M.; et al. The genetic basis of early T-cell precursor acute lymphoblastic leukaemia. Nature 2012, 481, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Goldman, F.D.; Gurel, Z.; Al-Zubeidi, D.; Fried, A.J.; Icardi, M.; Song, C.; Dovat, S. Congenital pancytopenia and absence of B lymphocytes in a neonate with a mutation in the Ikaros gene. Pediatr Blood Cancer 2012, 58, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Sakaguchi, H.; Muramatsu, H.; Okuno, Y.; Song, C.; Dovat, S.; Shimada, A.; Ozeki, M.; Ohnishi, H.; Teramoto, T.; et al. Germline IKAROS mutation associated with primary immunodeficiency that progressed to T-cell acute lymphoblastic leukemia. Leukemia 2017, 31, 1221–1223. [Google Scholar] [CrossRef]
- De Rooij, J.D.; Beuling, E.; van den Heuvel-Eibrink, M.M.; Obulkasim, A.; Baruchel, A.; Trka, J.; Reinhardt, D.; Sonneveld, E.; Gibson, B.E.; Pieters, R.; et al. Recurrent deletions of IKZF1 in pediatric acute myeloid leukemia. Haematologica 2015, 100, 1151–1159. [Google Scholar] [CrossRef] [Green Version]
- Theocharides, A.P.; Dobson, S.M.; Laurenti, E.; Notta, F.; Voisin, V.; Cheng, P.Y.; Yuan, J.S.; Guidos, C.J.; Minden, M.D.; Mullighan, C.G.; et al. Dominant-negative Ikaros cooperates with BCR-ABL1 to induce human acute myeloid leukemia in xenografts. Leukemia 2015, 29, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Unnikrishnan, A.; Guan, Y.F.; Huang, Y.; Beck, D.; Thoms, J.A.; Peirs, S.; Knezevic, K.; Ma, S.; de Walle, I.V.; de Jong, I.; et al. A quantitative proteomics approach identifies ETV6 and IKZF1 as new regulators of an ERG-driven transcriptional network. Nucleic Acids Res. 2016, 44, 10644–10661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Sligte, N.E.; Krumbholz, M.; Pastorczak, A.; Scheijen, B.; Tauer, J.T.; Nowasz, C.; Sonneveld, E.; de Bock, G.H.; Meeuwsen-de Boer, T.G.; van Reijmersdal, S.; et al. DNA copy number alterations mark disease progression in paediatric chronic myeloid leukaemia. Br. J. Haematol 2014, 166, 250–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosseler, M.; Marani, V.; Broukou, A.; Lequeux, A.; Kaoma, T.; Schlesser, V.; Francois, J.H.; Palissot, V.; Berchem, G.J.; Aouali, N.; et al. Inhibition of HIF1alpha-Dependent Upregulation of Phospho-l-Plastin Resensitizes Multiple Myeloma Cells to Frontline Therapy. Int J. Mol. Sci 2018, 19. [Google Scholar]
- Wang, L.; Howarth, A.; Clark, R.E. Ikaros transcripts Ik6/10 and levels of full-length transcript are critical for chronic myeloid leukaemia blast crisis transformation. Leukemia 2014, 28, 1745–1747. [Google Scholar] [CrossRef] [PubMed]
- Yagi, T.; Hibi, S.; Takanashi, M.; Kano, G.; Tabata, Y.; Imamura, T.; Inaba, T.; Morimoto, A.; Todo, S.; Imashuku, S. High frequency of Ikaros isoform 6 expression in acute myelomonocytic and monocytic leukemias: implications for up-regulation of the antiapoptotic protein Bcl-XL in leukemogenesis. Blood 2002, 99, 1350–1355. [Google Scholar] [CrossRef] [Green Version]
- Tefferi, A. Novel mutations and their functional and clinical relevance in myeloproliferative neoplasms: JAK2, MPL, TET2, ASXL1, CBL, IDH and IKZF1. Leukemia 2010, 24, 1128–1138. [Google Scholar] [CrossRef]
- Bavaro, L.; Martelli, M.; Cavo, M.; Soverini, S. Mechanisms of Disease Progression and Resistance to Tyrosine Kinase Inhibitor Therapy in Chronic Myeloid Leukemia: An Update. Int J. Mol. Sci 2019, 20, 6141. [Google Scholar] [CrossRef] [Green Version]
- Mullighan, C.G.; Su, X.; Zhang, J.; Radtke, I.; Phillips, L.A.; Miller, C.B.; Ma, J.; Liu, W.; Cheng, C.; Schulman, B.A.; et al. Deletion of IKZF1 and prognosis in acute lymphoblastic leukemia. N Engl J. Med. 2009, 360, 470–480. [Google Scholar] [CrossRef]
- Harvey, R.C.; Mullighan, C.G.; Chen, I.M.; Wharton, W.; Mikhail, F.M.; Carroll, A.J.; Kang, H.; Liu, W.; Dobbin, K.K.; Smith, M.A.; et al. Rearrangement of CRLF2 is associated with mutation of JAK kinases, alteration of IKZF1, Hispanic/Latino ethnicity, and a poor outcome in pediatric B-progenitor acute lymphoblastic leukemia. Blood 2010, 115, 5312–5321. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.C.; Mullighan, C.G.; Wang, X.; Dobbin, K.K.; Davidson, G.S.; Bedrick, E.J.; Chen, I.M.; Atlas, S.R.; Kang, H.; Ar, K.; et al. Identification of novel cluster groups in pediatric high-risk B-precursor acute lymphoblastic leukemia with gene expression profiling: correlation with genome-wide DNA copy number alterations, clinical characteristics, and outcome. Blood 2010, 116, 4874–4884. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, R.P.; Waanders, E.; van der Velden, V.H.; van Reijmersdal, S.V.; Venkatachalam, R.; Scheijen, B.; Sonneveld, E.; van Dongen, J.J.; Veerman, A.J.; van Leeuwen, F.N.; et al. IKZF1 deletions predict relapse in uniformly treated pediatric precursor B-ALL. Leukemia 2010, 24, 1258–1264. [Google Scholar] [CrossRef] [PubMed]
- Van der Veer, A.; Waanders, E.; Pieters, R.; Willemse, M.E.; Van Reijmersdal, S.V.; Russell, L.J.; Harrison, C.J.; Evans, W.E.; van der Velden, V.H.; Hoogerbrugge, P.M.; et al. Independent prognostic value of BCR-ABL1-like signature and IKZF1 deletion, but not high CRLF2 expression, in children with B-cell precursor ALL. Blood 2013, 122, 2622–2629. [Google Scholar] [CrossRef] [PubMed]
- Marke, R.; Havinga, J.; Cloos, J.; Demkes, M.; Poelmans, G.; Yuniati, L.; van Ingen Schenau, D.; Sonneveld, E.; Waanders, E.; Pieters, R.; et al. Tumor suppressor IKZF1 mediates glucocorticoid resistance in B-cell precursor acute lymphoblastic leukemia. Leukemia 2016, 30, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, G.; Iacobucci, I.; Soverini, S.; Piccaluga, P.P.; Cilloni, D.; Pane, F. New mechanisms of resistance in Philadelphia chromosome acute lymphoblastic leukemia. Expert Rev. Hematol 2009, 2, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, G.; Iacobucci, I.; Storlazzi, C.T.; Vignetti, M.; Paoloni, F.; Cilloni, D.; Soverini, S.; Vitale, A.; Chiaretti, S.; Cimino, G.; et al. IKZF1 (Ikaros) deletions in BCR-ABL1-positive acute lymphoblastic leukemia are associated with short disease-free survival and high rate of cumulative incidence of relapse: a GIMEMA AL WP report. J. Clin. Oncol 2009, 27, 5202–5207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Sif, S.; Jones, B.; Jackson, A.; Koipally, J.; Heller, E.; Winandy, S.; Viel, A.; Sawyer, A.; Ikeda, T.; et al. Ikaros DNA-binding proteins direct formation of chromatin remodeling complexes in lymphocytes. Immunity 1999, 10, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Koipally, J.; Kim, J.; Jones, B.; Jackson, A.; Avitahl, N.; Winandy, S.; Trevisan, M.; Nichogiannopoulou, A.; Kelley, C.; Georgopoulos, K. Ikaros chromatin remodeling complexes in the control of differentiation of the hemo-lymphoid system. Cold Spring Harb Symp Quant. Biol 1999, 64, 79–86. [Google Scholar] [CrossRef]
- Su, R.C.; Brown, K.E.; Saaber, S.; Fisher, A.G.; Merkenschlager, M.; Smale, S.T. Dynamic assembly of silent chromatin during thymocyte maturation. Nat. Genet. 2004, 36, 502–506. [Google Scholar] [CrossRef] [Green Version]
- Sridharan, R.; Smale, S.T. Predominant interaction of both Ikaros and Helios with the NuRD complex in immature thymocytes. J. Biol Chem 2007, 282, 30227–30238. [Google Scholar] [CrossRef] [Green Version]
- Koipally, J.; Renold, A.; Kim, J.; Georgopoulos, K. Repression by Ikaros and Aiolos is mediated through histone deacetylase complexes. Embo J. 1999, 18, 3090–3100. [Google Scholar] [CrossRef] [Green Version]
- Georgopoulos, K. Haematopoietic cell-fate decisions, chromatin regulation and ikaros. Nature Rev. Immunol 2002, 2, 162–174. [Google Scholar] [CrossRef]
- Brown, K.E.; Guest, S.S.; Smale, S.T.; Hahm, K.; Merkenschlager, M.; Fisher, A.G. Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin. Cell 1997, 91, 845–854. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Ebersole, T.; Kouprina, N.; Noskov, V.N.; Ohzeki, J.; Masumoto, H.; Mravinac, B.; Sullivan, B.A.; Pavlicek, A.; Dovat, S.; et al. Human gamma-satellite DNA maintains open chromatin structure and protects a transgene from epigenetic silencing. Genome Res. 2009, 19, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-del Arco, P.; Maki, K.; Georgopoulos, K. Phosphorylation controls Ikaros’s ability to negatively regulate the G(1)-S transition. Mol. Cell Biol 2004, 24, 2797–2807. [Google Scholar] [CrossRef] [Green Version]
- Gomez-del Arco, P.; Koipally, J.; Georgopoulos, K. Ikaros SUMOylation: switching out of repression. Mol. Cell Biol 2005, 25, 2688–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.M.; Soares, M.V.; Ribeiro, P.; Caldas, J.; Povoa, V.; Martins, L.R.; Melao, A.; Serra-Caetano, A.; de Sousa, A.B.; Lacerda, J.F.; et al. Adult B-cell acute lymphoblastic leukemia cells display decreased PTEN activity and constitutive hyperactivation of PI3K/Akt pathway despite high PTEN protein levels. Haematologica 2014, 99, 1062–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evangelisti, C.; Chiarini, F.; McCubrey, J.A.; Martelli, A.M. Therapeutic Targeting of mTOR in T-Cell Acute Lymphoblastic Leukemia: An Update. Int J. Mol. Sci 2018, 19, 1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurel, Z.; Ronni, T.; Ho, S.; Kuchar, J.; Payne, K.J.; Turk, C.W.; Dovat, S. Recruitment of ikaros to pericentromeric heterochromatin is regulated by phosphorylation. J. Biol Chem 2008, 283, 8291–8300. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Gowda, C.; Pan, X.; Ding, Y.; Tong, Y.; Tan, B.H.; Wang, H.; Muthusami, S.; Ge, Z.; Sachdev, M.; et al. Targeting casein kinase II restores Ikaros tumor suppressor activity and demonstrates therapeutic efficacy in high-risk leukemia. Blood 2015, 126, 1813–1822. [Google Scholar] [CrossRef] [Green Version]
- Gowda, C.; Sachdev, M.; Muthusami, S.; Kapadia, M.; Petrovic-Dovat, L.; Hartman, M.; Ding, Y.; Song, C.; Payne, J.L.; Tan, B.H.; et al. Casein Kinase II (CK2) as a Therapeutic Target for Hematological Malignancies. Curr Pharm Des. 2017, 23, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Gowda, C.; Soliman, M.; Kapadia, M.; Ding, Y.; Payne, K.; Dovat, S. Casein Kinase II (CK2), Glycogen Synthase Kinase-3 (GSK-3) and Ikaros mediated regulation of leukemia. Adv. Biol Regul 2017, 65, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Gowda, C.; Song, C.; Kapadia, M.; Payne, J.L.; Hu, T.; Ding, Y.; Dovat, S. Regulation of cellular proliferation in acute lymphoblastic leukemia by Casein Kinase II (CK2) and Ikaros. Adv. Biol Regul 2017, 63, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Popescu, M.; Gurel, Z.; Ronni, T.; Song, C.; Hung, K.Y.; Payne, K.J.; Dovat, S. Ikaros stability and pericentromeric localization are regulated by protein phosphatase 1. J. Biol Chem 2009, 284, 13869–13880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumby, M.C.; Walter, G. Protein serine/threonine phosphatases: structure, regulation, and functions in cell growth. Physiol Rev. 1993, 73, 673–699. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Lee, E.Y. Targeting of the catalytic subunit of protein phosphatase-1 to the glycolytic enzyme phosphofructokinase. Biochemistry 1997, 36, 8318–8324. [Google Scholar] [CrossRef]
- Armstrong, C.G.; Doherty, M.J.; Cohen, P.T. Identification of the separate domains in the hepatic glycogen-targeting subunit of protein phosphatase 1 that interact with phosphorylase a, glycogen and protein phosphatase 1. Biochem J. 1998, 336 (Pt 3), 699–704. [Google Scholar] [CrossRef] [Green Version]
- Cohen, P.T. Protein phosphatase 1--targeted in many directions. J. Cell Sci 2002, 115 (Pt 2), 241–256. [Google Scholar]
- Ceulemans, H.; Bollen, M. Functional diversity of protein phosphatase-1, a cellular economizer and reset button. Physiol Rev. 2004, 84, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, B.; Li, N.; Li, H.; Qiu, J.; Zhang, Y.; Cao, X. IPP5, a novel protein inhibitor of protein phosphatase 1, promotes G1/S progression in a Thr-40-dependent manner. J. Biol Chem 2008, 283, 12076–12084. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.; Beullens, M.; Bollen, M.; Van Eynde, A. Functions and therapeutic potential of protein phosphatase 1: Insights from mouse genetics. Biochim Biophys Acta Mol. Cell Res. 2019, 1866, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Gil, R.S.; Vagnarelli, P. Protein phosphatases in chromatin structure and function. Biochim Biophys Acta Mol. Cell Res. 2019, 1866, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Song, C.; Gurel, Z.; Song, N.; Ma, J.; Ouyang, H.; Lai, L.; Payne, K.J.; Dovat, S. Protein Phosphatase 1 (PP1) and Casein Kinase II (CK2) regulate Ikaros-mediated repression of TdT in thymocytes and T-cell leukemia. Pediatr Blood Cancer 2014, 61, 2230–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Li, Z.; Erbe, A.K.; Savic, A.; Dovat, S. Regulation of Ikaros function by casein kinase 2 and protein phosphatase 1. World J. Biol Chem 2011, 2, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Dovat, S.; Song, C.; Payne, K.J.; Li, Z. Ikaros, CK2 kinase, and the road to leukemia. Mol. Cell Biochem 2011, 356, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Z.; Gu, Y.; Xiao, L.; Han, Q.; Li, J.; Chen, B.; Yu, J.; Kawasawa, Y.I.; Payne, K.J.; Dovat, S.; et al. Co-existence of IL7R high and SH2B3 low expression distinguishes a novel high-risk acute lymphoblastic leukemia with Ikaros dysfunction. Oncotarget 2016, 7, 46014–46027. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Gu, Y.; Zhao, G.; Li, J.; Chen, B.; Han, Q.; Guo, X.; Liu, J.; Li, H.; Yu, M.D.; et al. High CRLF2 expression associates with IKZF1 dysfunction in adult acute lymphoblastic leukemia without CRLF2 rearrangement. Oncotarget 2016, 7, 49722–49732. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Guo, X.; Li, J.; Hartman, M.; Kawasawa, Y.I.; Dovat, S.; Song, C. Clinical significance of high c-MYC and low MYCBP2 expression and their association with Ikaros dysfunction in adult acute lymphoblastic leukemia. Oncotarget 2015, 6, 42300–42311. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Han, Q.; Gu, Y.; Ge, Q.; Ma, J.; Sloane, J.; Gao, G.; Payne, K.J.; Szekely, L.; Song, C.; et al. Aberrant ARID5B expression and its association with Ikaros dysfunction in acute lymphoblastic leukemia. Oncogenesis 2018, 7, 84. [Google Scholar] [CrossRef]
- Ge, Z.; Li, M.; Zhao, G.; Xiao, L.; Gu, Y.; Zhou, X.; Yu, M.D.; Li, J.; Dovat, S.; Song, C. Novel dynamin 2 mutations in adult T-cell acute lymphoblastic leukemia. Oncol Lett 2016, 12, 2746–2751. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Song, E.J.; Kawasawa, Y.I.; Li, J.; Dovat, S.; Song, C. WDR5 high expression and its effect on tumorigenesis in leukemia. Oncotarget 2016, 7, 37740–37754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Z.; Zhou, X.; Gu, Y.; Han, Q.; Li, J.; Chen, B.; Ge, Q.; Dovat, E.; Payne, J.L.; Sun, T.; et al. Ikaros regulation of the BCL6/BACH2 axis and its clinical relevance in acute lymphoblastic leukemia. Oncotarget 2017, 8, 8022–8034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dovat, S. Ikaros in hematopoiesis and leukemia. World J. Biol Chem 2011, 2, 105–107. [Google Scholar] [CrossRef]
- Wang, H.; Ouyang, H.; Lai, L.; Petrovic-Dovat, L.; Stankov, K.; Bogdanovic, G.; Dovat, S. Pathogenesis and regulation of cellular proliferation in acute lymphoblastic leukemia—the role of Ikaros. J. BUON 2014, 19, 22–28. [Google Scholar] [PubMed]
- Ronni, T.; Payne, K.J.; Ho, S.; Bradley, M.N.; Dorsam, G.; Dovat, S. Human Ikaros function in activated T cells is regulated by coordinated expression of its largest isoforms. J. Biol Chem 2007, 282, 2538–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, K.J.; Dovat, S. Ikaros and tumor suppression in acute lymphoblastic leukemia. Crit Rev. Oncog 2011, 16, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Zhang, B.; Payne, J.L.; Song, C.; Ge, Z.; Gowda, C.; Iyer, S.; Dhanyamraju, P.K.; Dorsam, G.; Reeves, M.E.; et al. Ikaros tumor suppressor function includes induction of active enhancers and super-enhancers along with pioneering activity. Leukemia 2019, 33, 2720–2731. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Ma, J.; Gu, Y.; Song, H.; Kapadia, M.; Kawasawa, Y.I.; Dovat, S.; Song, C.; Ge, Z. RAG1 high expression associated with IKZF1 dysfunction in adult B-cell acute lymphoblastic leukemia. J. Cancer 2019, 10, 3842–3850. [Google Scholar] [CrossRef] [Green Version]
- Habermann, J.K.; Brucker, C.A.; Freitag-Wolf, S.; Heselmeyer-Haddad, K.; Kruger, S.; Barenboim, L.; Downing, T.; Bruch, H.P.; Auer, G.; Roblick, U.J.; et al. Genomic instability and oncogene amplifications in colorectal adenomas predict recurrence and synchronous carcinoma. Mod. Pathol 2011, 24, 542–555. [Google Scholar] [CrossRef] [Green Version]
- Simoneau, E.; Chicoine, J.; Negi, S.; Salman, A.; Lazaris, A.; Hassanain, M.; Beauchemin, N.; Petrillo, S.; Valenti, D.; Amre, R.; et al. Next generation sequencing of progressive colorectal liver metastases after portal vein embolization. Clin. Exp. Metastasis 2017, 34, 351–361. [Google Scholar] [CrossRef]
- Javierre, B.M.; Rodriguez-Ubreva, J.; Al-Shahrour, F.; Corominas, M.; Grana, O.; Ciudad, L.; Agirre, X.; Pisano, D.G.; Valencia, A.; Roman-Gomez, J.; et al. Long-range epigenetic silencing associates with deregulation of Ikaros targets in colorectal cancer cells. Mol. Cancer Res. 2011, 9, 1139–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellagatti, A.; Esoof, N.; Watkins, F.; Langford, C.F.; Vetrie, D.; Campbell, L.J.; Fidler, C.; Cavenagh, J.D.; Eagleton, H.; Gordon, P.; et al. Gene expression profiling in the myelodysplastic syndromes using cDNA microarray technology. Br. J. Haematol 2004, 125, 576–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crescenzi, B.; La Starza, R.; Romoli, S.; Beacci, D.; Matteucci, C.; Barba, G.; Aventin, A.; Marynen, P.; Ciolli, S.; Nozzoli, C.; et al. Submicroscopic deletions in 5q- associated malignancies. Haematologica 2004, 89, 281–285. [Google Scholar] [PubMed]
- Kakinuma, S.; Kodama, Y.; Amasaki, Y.; Yi, S.; Tokairin, Y.; Arai, M.; Nishimura, M.; Monobe, M.; Kojima, S.; Shimada, Y. Ikaros is a mutational target for lymphomagenesis in Mlh1-deficient mice. Oncogene 2007, 26, 2945–2949. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Lin, S.; Zou, L.; Zhou, H.; Zhang, J.; Su, B.; Wan, Y. Expression profiling of Rab GTPases reveals the involvement of Rab20 and Rab32 in acute brain inflammation in mice. Neurosci Lett 2012, 527, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Pei, G.; Bronietzki, M.; Gutierrez, M.G. Immune regulation of Rab proteins expression and intracellular transport. J. Leukoc Biol 2012, 92, 41–50. [Google Scholar] [CrossRef]
- Torri, A.; Beretta, O.; Ranghetti, A.; Granucci, F.; Ricciardi-Castagnoli, P.; Foti, M. Gene expression profiles identify inflammatory signatures in dendritic cells. PLoS One 2010, 5, e9404. [Google Scholar] [CrossRef]
- Pei, G.; Schnettger, L.; Bronietzki, M.; Repnik, U.; Griffiths, G.; Gutierrez, M.G. Interferon-gamma-inducible Rab20 regulates endosomal morphology and EGFR degradation in macrophages. Mol. Biol Cell 2015, 26, 3061–3070. [Google Scholar] [CrossRef]
- Gutierrez, M.G.; Mishra, B.B.; Jordao, L.; Elliott, E.; Anes, E.; Griffiths, G. NF-kappa B activation controls phagolysosome fusion-mediated killing of mycobacteria by macrophages. J. Immunol 2008, 181, 2651–2663. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.J.; Huh, J.E.; Song, J.; Rhee, D.K.; Pyo, S. Ikaros negatively regulates inducible nitric oxide synthase expression in macrophages: involvement of Ikaros phosphorylation by casein kinase 2. Cell Mol. Life Sci 2008, 65, 3290–3303. [Google Scholar] [CrossRef]
- Dumortier, A.; Kirstetter, P.; Kastner, P.; Chan, S. Ikaros regulates neutrophil differentiation. Blood 2003, 101, 2219–2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, H.; Ishimaru, F.; Katayama, Y.; Nakase, K.; Sezaki, N.; Takenaka, K.; Shinagawa, K.; Ikeda, K.; Niiya, K.; Harada, M. Ikaros expression in human hematopoietic lineages. Exp. Hematol 2000, 28, 1232–1238. [Google Scholar] [CrossRef]
- Winandy, S.; Wu, L.; Wang, J.H.; Georgopoulos, K. Pre-T cell receptor (TCR) and TCR-controlled checkpoints in T cell differentiation are set by Ikaros. J. Exp. Med. 1999, 190, 1039–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amillet, J.M.; Ferbus, D.; Real, F.X.; Antony, C.; Muleris, M.; Gress, T.M.; Goubin, G. Characterization of human Rab20 overexpressed in exocrine pancreatic carcinoma. Hum. Pathol 2006, 37, 256–263. [Google Scholar] [CrossRef]
- Turner, N.; Lambros, M.B.; Horlings, H.M.; Pearson, A.; Sharpe, R.; Natrajan, R.; Geyer, F.C.; van Kouwenhove, M.; Kreike, B.; Mackay, A.; et al. Integrative molecular profiling of triple negative breast cancers identifies amplicon drivers and potential therapeutic targets. Oncogene 2010, 29, 2013–2023. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.R.; Chapeaublanc, E.; Kirkwood, L.; Nicolle, R.; Benhamou, S.; Lebret, T.; Allory, Y.; Southgate, J.; Radvanyi, F.; Goud, B. Deregulation of Rab and Rab effector genes in bladder cancer. PLoS One 2012, 7, e39469. [Google Scholar] [CrossRef]
- Song, C.; Pan, X.; Ge, Z.; Gowda, C.; Ding, Y.; Li, H.; Li, Z.; Yochum, G.; Muschen, M.; Li, Q.; et al. Epigenetic regulation of gene expression by Ikaros, HDAC1 and Casein Kinase II in leukemia. Leukemia 2016, 30, 1436–1440. [Google Scholar] [CrossRef] [Green Version]
- Stenmark, H.; Olkkonen, V.M. The Rab GTPase family. Genome Biol 2001, 2. [Google Scholar]
- Fukuda, M. Regulation of secretory vesicle traffic by Rab small GTPases. Cell Mol. Life Sci 2008, 65, 2801–2813. [Google Scholar] [CrossRef]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol 2009, 10, 513–525. [Google Scholar] [CrossRef]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol 2001, 2, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.G. Functional role(s) of phagosomal Rab GTPases. Small GTPases 2013, 4, 148–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, G.; Repnik, U.; Griffiths, G.; Gutierrez, M.G. Identification of an immune-regulated phagosomal Rab cascade in macrophages. J. Cell Sci 2014, 127, 2071–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, M.H.; Li, N.; Lao, Q.; Gottschalk, R.A.; Hager, G.L.; Fraser, I.D. Switching of the relative dominance between feedback mechanisms in lipopolysaccharide-induced NF-kappaB signaling. Sci Signal. 2014, 7. [Google Scholar]
- Oh, K.S.; Gottschalk, R.A.; Lounsbury, N.W.; Sun, J.; Dorrington, M.G.; Baek, S.; Sun, G.; Wang, Z.; Krauss, K.S.; Milner, J.D.; et al. Dual Roles for Ikaros in Regulation of Macrophage Chromatin State and Inflammatory Gene Expression. J. Immunol 2018, 201, 757–771. [Google Scholar] [CrossRef]
- Navasa, N.; Martin-Ruiz, I.; Atondo, E.; Sutherland, J.D.; Angel Pascual-Itoiz, M.; Carreras-Gonzalez, A.; Izadi, H.; Tomas-Cortazar, J.; Ayaz, F.; Martin-Martin, N.; et al. Ikaros mediates the DNA methylation-independent silencing of MCJ/DNAJC15 gene expression in macrophages. Sci Rep. 2015, 5, 14692. [Google Scholar] [CrossRef]
- Dovat, S.; Ronni, T.; Russell, D.; Ferrini, R.; Cobb, B.S.; Smale, S.T. A common mechanism for mitotic inactivation of C2H2 zinc finger DNA-binding domains. Genes Dev. 2002, 16, 2985–2990. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Song, C.; Ouyang, H.; Lai, L.; Payne, K.J.; Dovat, S. Cell cycle-specific function of Ikaros in human leukemia. Pediatr Blood Cancer 2012, 59, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Gu, Y.; Han, Q.; Sloane, J.; Ge, Q.; Gao, G.; Ma, J.; Song, H.; Hu, J.; Chen, B.; et al. Plant homeodomain finger protein 2 as a novel IKAROS target in acute lymphoblastic leukemia. Epigenomics 2018, 10, 59–69. [Google Scholar] [CrossRef]
- Ge, Z.; Gu, Y.; Han, Q.; Zhao, G.; Li, M.; Li, J.; Chen, B.; Sun, T.; Dovat, S.; Gale, R.P.; et al. Targeting High Dynamin-2 (DNM2) Expression by Restoring Ikaros Function in Acute Lymphoblastic Leukemia. Sci Rep. 2016, 6, 38004. [Google Scholar] [CrossRef] [Green Version]
- Gowda, C.S.; Song, C.; Ding, Y.; Kapadia, M.; Dovat, S. Protein signaling and regulation of gene transcription in leukemia: role of the Casein Kinase II-Ikaros axis. J. Investig Med. 2016, 64, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Schjerven, H.; Ayongaba, E.F.; Aghajanirefah, A.; McLaughlin, J.; Cheng, D.; Geng, H.; Boyd, J.R.; Eggesbo, L.M.; Lindeman, I.; Heath, J.L.; et al. Genetic analysis of Ikaros target genes and tumor suppressor function in BCR-ABL1(+) pre-B ALL. J. Exp. Med. 2017, 214, 793–814. [Google Scholar] [CrossRef] [PubMed]
- Polak, R.; Bierings, M.B.; van der Leije, C.S.; Sanders, M.A.; Roovers, O.; Marchante, J.R.M.; Boer, J.M.; Cornelissen, J.J.; Pieters, R.; den Boer, M.L.; et al. Autophagy inhibition as a potential future targeted therapy for ETV6-RUNX1-driven B-cell precursor acute lymphoblastic leukemia. Haematologica 2019, 104, 738–748. [Google Scholar] [CrossRef]
- Cobb, B.S.; Morales-Alcelay, S.; Kleiger, G.; Brown, K.E.; Fisher, A.G.; Smale, S.T. Targeting of Ikaros to pericentromeric heterochromatin by direct DNA binding. Genes Dev. 2000, 14, 2146–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Song, C.; Ding, Y.; Pan, X.; Ge, Z.; Tan, B.H.; Gowda, C.; Sachdev, M.; Muthusami, S.; Ouyang, H.; et al. Transcriptional Regulation of JARID1B/KDM5B Histone Demethylase by Ikaros, Histone Deacetylase 1 (HDAC1), and Casein Kinase 2 (CK2) in B-cell Acute Lymphoblastic Leukemia. J. Biol Chem 2016, 291, 4004–4018. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, T.; O’Geen, H.; Keles, S.; Blahnik, K.; Linnemann, A.K.; Kang, Y.A.; Choi, K.; Farnham, P.J.; Bresnick, E.H. Discovering hematopoietic mechanisms through genome-wide analysis of GATA factor chromatin occupancy. Mol. Cell 2009, 36, 667–681. [Google Scholar] [CrossRef] [Green Version]
- Hahm, K.; Cobb, B.S.; McCarty, A.S.; Brown, K.E.; Klug, C.A.; Lee, R.; Akashi, K.; Weissman, I.L.; Fisher, A.G.; Smale, S.T. Helios, a T cell-restricted Ikaros family member that quantitatively associates with Ikaros at centromeric heterochromatin. Genes Dev. 1998, 12, 782–796. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Payne, J.L.; Song, C.; Ding, Y.; Dhanyamraju, P.K.; Bamme, Y.; Schramm, J.W.; Desai, D.; Sharma, A.; Gowda, C.; Dovat, S. Regulation of Small GTPase Rab20 by Ikaros in B-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2020, 21, 1718. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051718
Payne JL, Song C, Ding Y, Dhanyamraju PK, Bamme Y, Schramm JW, Desai D, Sharma A, Gowda C, Dovat S. Regulation of Small GTPase Rab20 by Ikaros in B-Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences. 2020; 21(5):1718. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051718
Chicago/Turabian StylePayne, Jonathon L, Chunhua Song, Yali Ding, Pavan Kumar Dhanyamraju, Yevgeniya Bamme, Joseph W Schramm, Dhimant Desai, Arati Sharma, Chandrika Gowda, and Sinisa Dovat. 2020. "Regulation of Small GTPase Rab20 by Ikaros in B-Cell Acute Lymphoblastic Leukemia" International Journal of Molecular Sciences 21, no. 5: 1718. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051718