Male sterile 305 Mutation Leads the Misregulation of Anther Cuticle Formation by Disrupting Lipid Metabolism in Maize

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
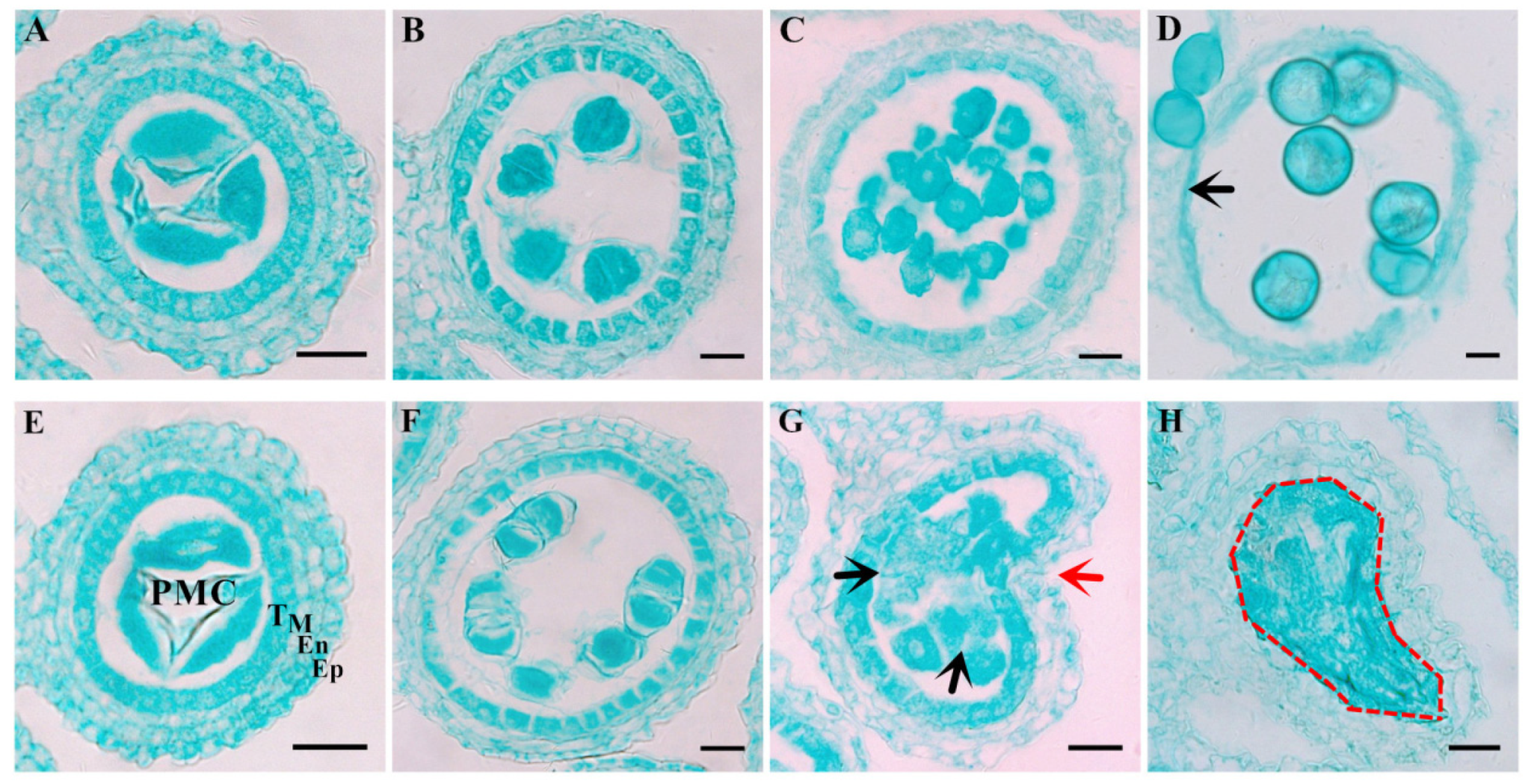
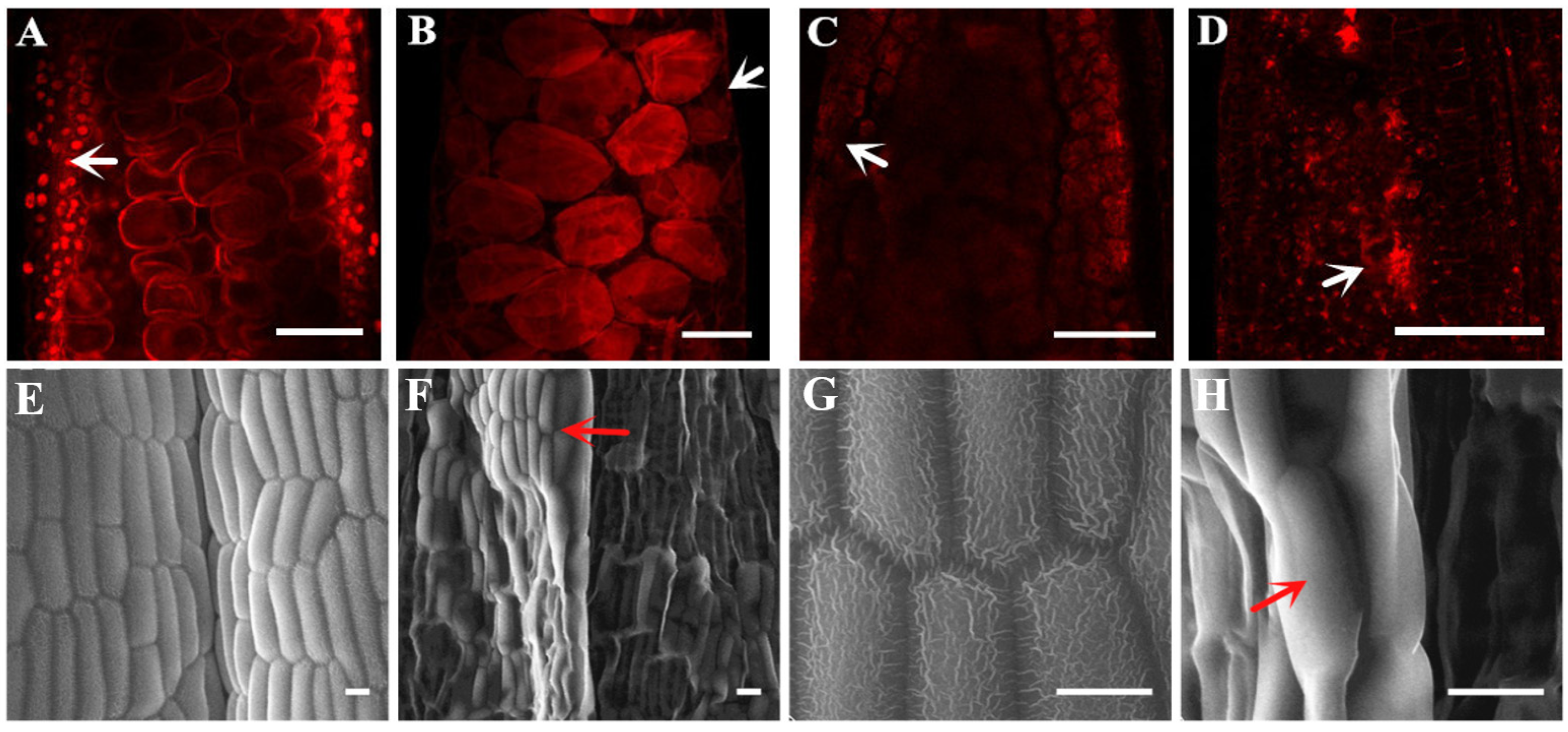
2.1. Characterization of the Male Sterile Mutant ms305 Anthers
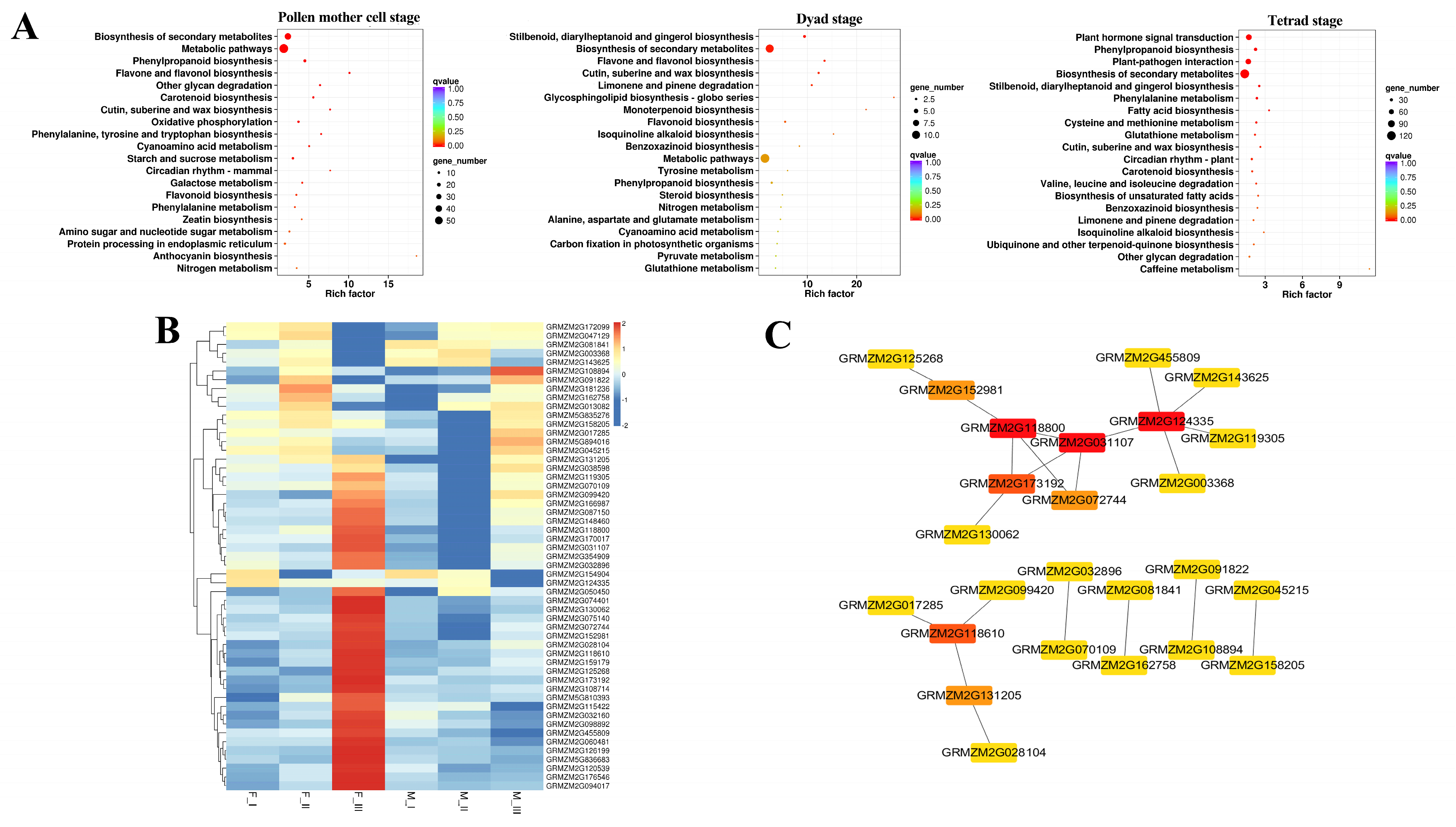
2.2. Transcriptional Differential Expression Analysis of ms305 and its Fertile Sibling Anther
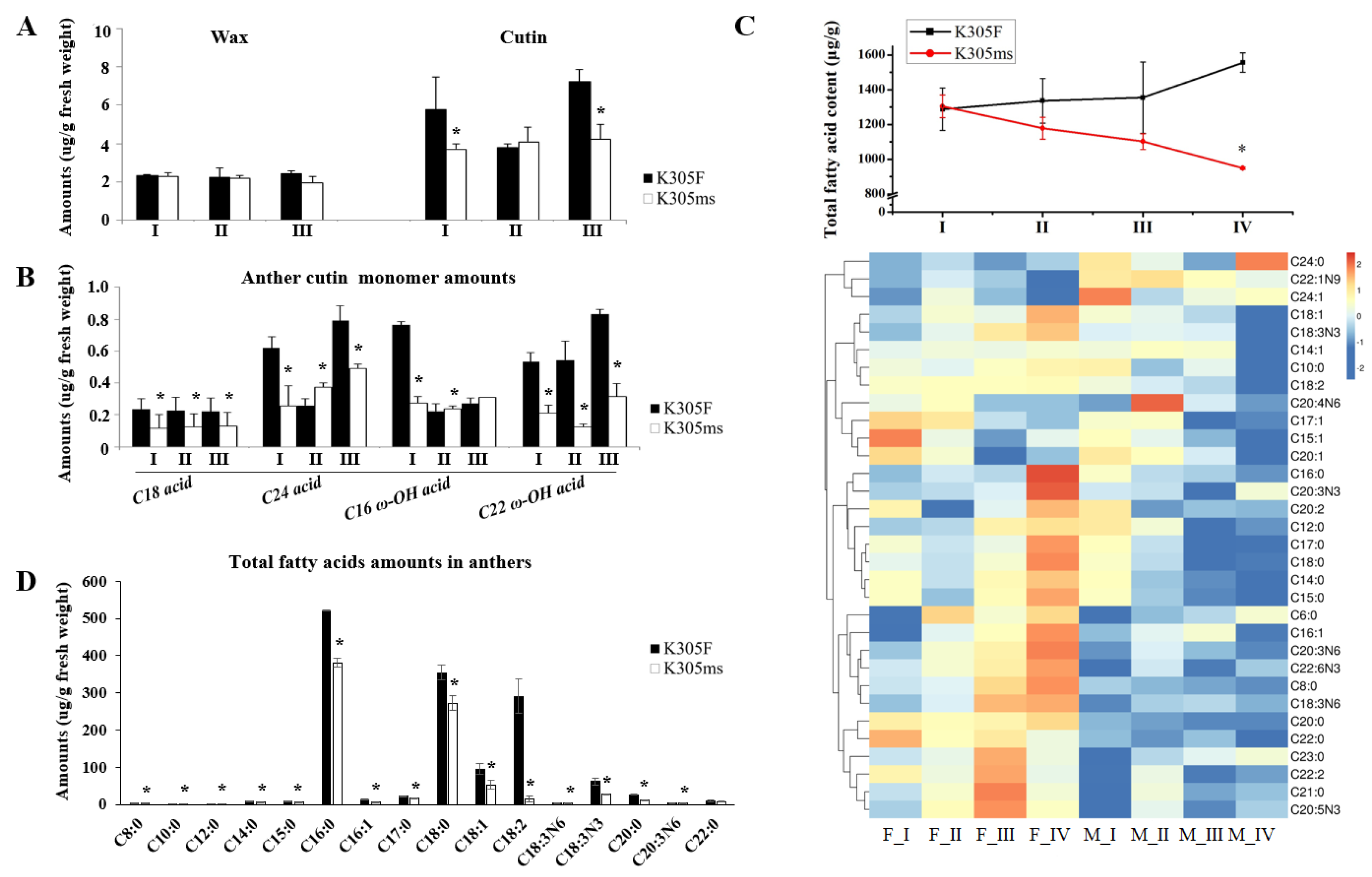
2.3. Aliphatic Alteration of ms305 Anther
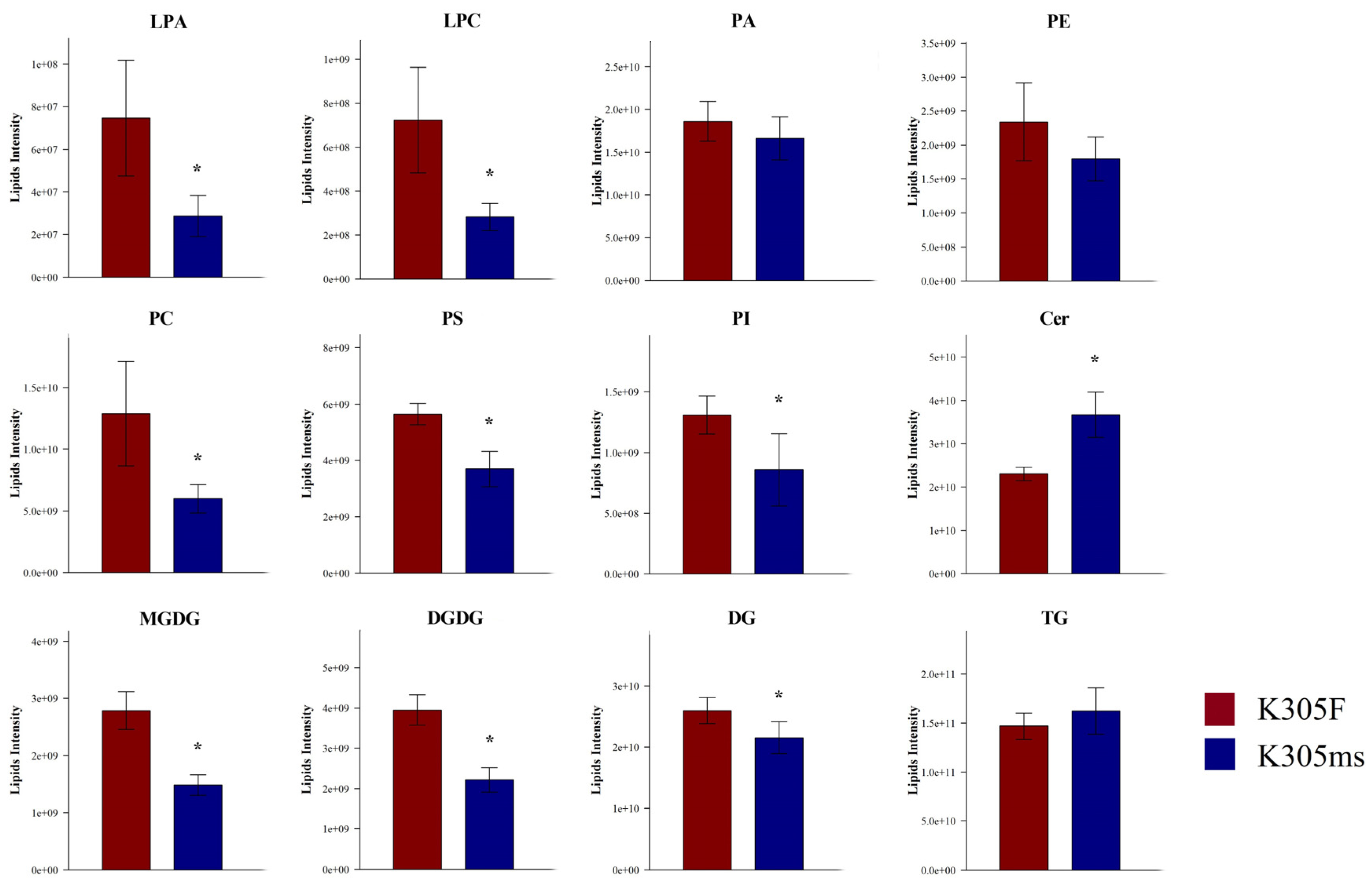
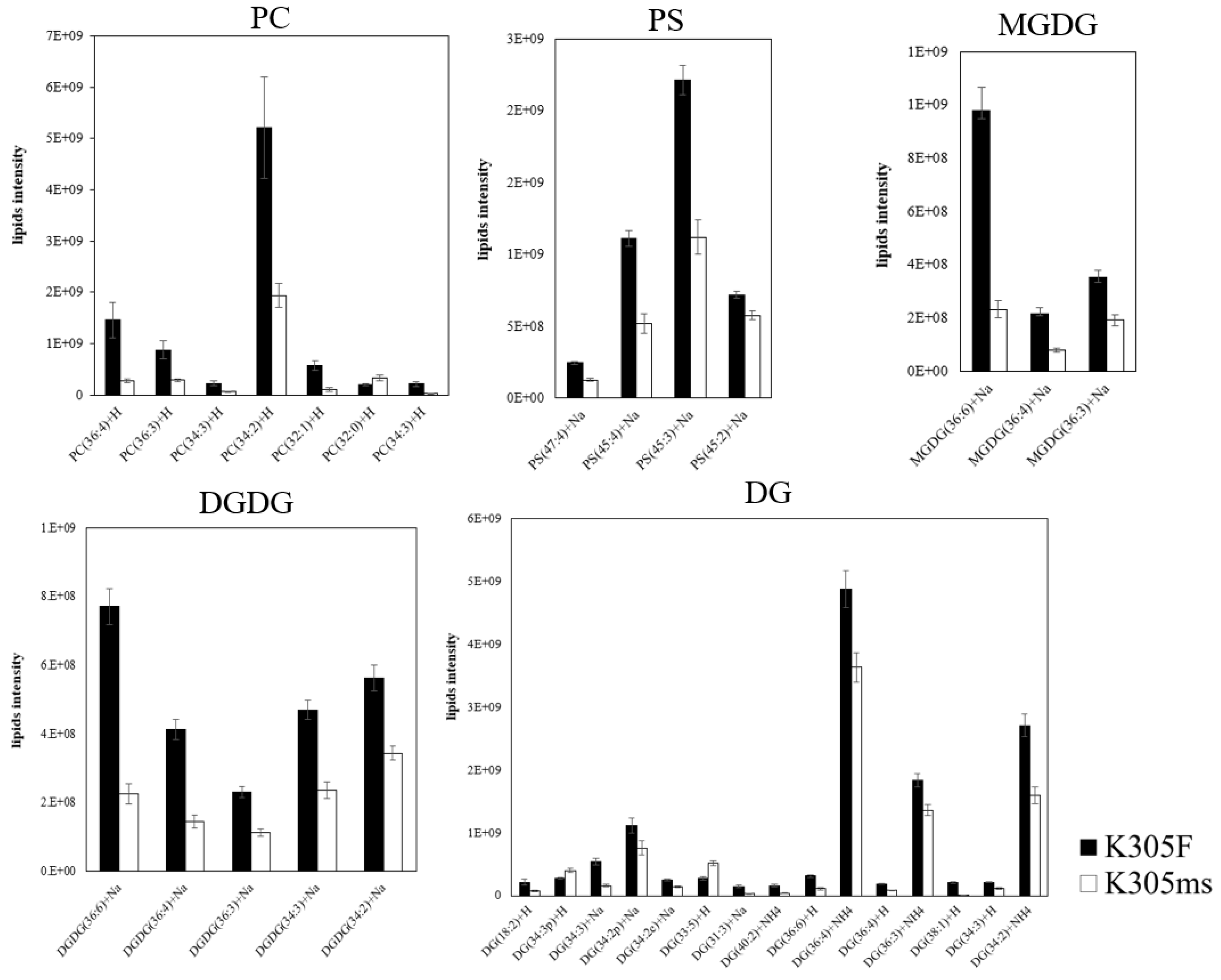
2.4. Lipids Alteration of ms305 Anther
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Phenotypic Analysis of ms305
4.3. Aliphatic Components Analysis
4.4. RNA Extraction and qRT-PCR
4.5. Transcriptome Analysis
4.6. Lipidome Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Miller, F.P. Detasseling; Alphascript Publishing: Saarbrucken, Germany, 2010; ISBN 978-613-1-79269-4. [Google Scholar]
- Cantrell, G.R.; Geadelmann, J.L. Contribution of Husk Leaves to Maize Grain Yield. Cropence 1981, 21, 544–546. [Google Scholar] [CrossRef]
- Wilhelm, W.W.; Johnson, B.E.; Schepers, J.S. Yield, Quality, and Nitrogen Use of Inbred Corn with Varying Numbers of Leaves during Detasseling. Crop Sci. 1995, 35, 209–212. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.-G. Male Sterility and Fertility Restoration in Crops. Ann. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Kempe, K.; Gils, M. Pollination control technologies for hybrid breeding. Mol. Breed. 2011, 27, 417–437. [Google Scholar] [CrossRef]
- Jung, K.H. Rice Undeveloped Tapetum1 is a major regulator of early tapetum development. Plant Cell 2005, 17, 2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J. Male reproductive development: Gene expression profiling of maize anther and pollen ontogeny. Genome Biol. 2008, 9, R181. [Google Scholar] [CrossRef] [Green Version]
- Skibbe, D.S. Mutator transposon activity reprograms the transcriptomes and proteomes of developing maize anthers. Plant J. 2009, 59, 622–633. [Google Scholar] [CrossRef]
- Zhang, D. Construction of a multi-control sterility system for a maize male-sterile line and hybrid seed production based on the ZmMs7 gene encoding a PHD-finger transcription factor. Plant Biotechnol. J. 2017, 16, 459. [Google Scholar] [CrossRef] [Green Version]
- Wang, D. A low molecular weight proteome comparison of fertile and male sterile 8 anthers of Zea mays. Plant Biotechnol. J. 2012, 10, 925–935. [Google Scholar] [CrossRef] [Green Version]
- Albertsen, M.C. Cloning and Use of the MS9 Gene from Maize. WIPO Patent Application WO/2014/152447, 13 September 2018. [Google Scholar]
- Albertsen, M.C. MSCA1 Nucleotide Sequences Impacting Plant Male Fertility and Method of Using Same. U.S. Patent 7,915,478, 29 March 2011. [Google Scholar]
- Nan, G.-L. MS23, a master basic helix-loop-helix factor, regulates the specification and development of the tapetum in maize. Development 2017, 144, 163. [Google Scholar] [CrossRef] [Green Version]
- Djukanovic, V. Male-sterile maize plants produced by targeted mutagenesis of the cytochrome P450-like gene (MS26) using a re-designed I-CreI homing endonuclease. Plant J. Cell Mol. Biol. 2013, 76, 888–899. [Google Scholar] [CrossRef] [PubMed]
- An, X. ZmMs30 Encoding a Novel GDSL Lipase Is Essential for Male Fertility and Valuable for Hybrid Breeding in Maize. Mol. Plant 2019, 12, 343–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J. Regulation of cell divisions and differentiation by MALE STERILITY32 is required for anther development in maize. Plant J. 2013, 76, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Xie, K. Map-based cloning and characterization of Zea mays male sterility33 (ZmMs33) gene, encoding a glycerol-3-phosphate acyltransferase. Theor. Appl. Genet. 2018, 131, 1363–1378. [Google Scholar] [CrossRef] [Green Version]
- Cigan, A.M. Phenotypic complementation of ms45 maize requires tapetal expression of MS45. Sex. Plant Reprod. 2001, 14, 135–142. [Google Scholar] [CrossRef]
- Somaratne, Y. ABNORMAL POLLEN VACUOLATION1 (APV1) is required for male fertility by contributing to anther cuticle and pollen exine formation in maize. Plant J. 2017, 90, 96–110. [Google Scholar] [CrossRef]
- Chen, X. Irregular Pollen Exine1 is a Novel Factor in Anther Cuticle and Pollen Exine Formation. Plant Physiol. 2017, 173, 307. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.J. Maize multiple archesporial cells 1 (mac1), an ortholog of rice TDL1A, modulates cell proliferation and identity in early anther development. Development 2012, 139, 2594–2603. [Google Scholar] [CrossRef] [Green Version]
- Vernoud, V. The HD-ZIP IV transcription factor OCL4 is necessary for trichome patterning and anther development in maize. Plant J. 2009, 59, 883–894. [Google Scholar] [CrossRef]
- Gómez, J.F.; Talle, B.; Wilson, Z.A. Anther and pollen development: A conserved developmental pathway. J. Integr. Plant Biol. 2015, 57, 876–891. [Google Scholar] [CrossRef] [Green Version]
- Wilson, Z.A.; Zhang, D.B. From Arabidopsis to rice: Pathways in pollen development. J. Exp. Bot. 2009, 60, 1479–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, E. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2009, 50, S9–S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Shi, J.; Yang, X. Role of Lipid Metabolism in Plant Pollen Exine Development. Subcell. Biochem. 2016, 86, 315–337. [Google Scholar] [PubMed]
- Narayanan, S.; Prasad, P.V.V.; Welti, R. Alterations in wheat pollen lipidome during high day and night temperature stress: Heat induced alterations in wheat pollen lipidome. Plant Cell Environ. 2018, 41, 1749–1761. [Google Scholar] [CrossRef]
- de Azevedo Souza, C. A novel fatty Acyl-CoA Synthetase is required for pollen development and sporopollenin biosynthesis in Arabidopsis. Plant Cell 2009, 21, 507–525. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y. MALE STERILE6021 (MS6021) is required for the development of anther cuticle and pollen exine in maize. Sci. Rep. 2017, 7, 16736. [Google Scholar] [CrossRef]
- Mondol, P.C. Defective Pollen Wall 3 (DPW3), a novel alpha integrin-like protein, is required for pollen wall formation in rice. New Phytol. 2020, 225, 807–822. [Google Scholar] [CrossRef]
- Zienkiewicz, A. New insights into the early steps of oil body mobilization during pollen germination. J. Exp. Bot. 2013, 64, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.R. ABNORMAL POLLEN TUBE GUIDANCE1, an Endoplasmic Reticulum-Localized Mannosyltransferase Homolog of Glycosylphosphatidylinositol10 in Yeast and Phosphatidylinositol glycan anchor biosynthesis B in Human, is Required for Arabidopsis Pollen Tube Micropylar Gui. Plant Physiol. 2014, 165, 1544–1556. [Google Scholar] [CrossRef]
- Li, X.C. Glycerol-3-Phosphate Acyltransferase 6 (GPAT6) is Important for Tapetum Development in Arabidopsis and Plays Multiple Roles in Plant Fertility. Mol. Plant 2012, 5, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Shi, H. Combining ability and yield heterosis of maize nuclear sterility mutant K305ms. J. Nucl. Agric. Sci. 2016, 30, 621–628. [Google Scholar]
- Wang, Y. Characterization and genetic mapping of a novel recessive genic male sterile gene ms305 in maize (Zea mays L.). Isr. J. Plant Sci. 2015, 62, 208–214. [Google Scholar] [CrossRef]
- Huawei, C. Genetic analysis of male sterility mutants induced by (60)Co-γ ray in maize. J. Nucl. Agric. Sci. 2016, 30, 829–834. [Google Scholar]
- Song, S. OsFTIP7 determines auxin-mediated anther dehiscence in rice. Nat. Plants 2018, 4, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Wan, L. A rice β-1,3-glucanase geneOsg1is required for callose degradation in pollen development. Planta 2011, 233, 309–323. [Google Scholar] [CrossRef]
- Zhang, L. Maize male sterile 33 encodes a putative glycerol-3-phosphate acyltransferase that mediates anther cuticle formation and microspore development. BMC Plant Biol. 2018, 18, 318. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y. Metacaspase gene family in Rosaceae genomes: Comparative genomic analysis and their expression during pear pollen tube and fruit development. PLoS ONE 2019, 14, e0211635. [Google Scholar] [CrossRef] [Green Version]
- Singh, R. Reactive Oxygen Species (ROS): Beneficial Companions of Plants’ Developmental Processes. Front. Plant Sci. 2016, 7, 1299. [Google Scholar] [CrossRef] [Green Version]
- Xu, D. Defective Pollen Wall 2 (DPW2) Encodes an Acyl Transferase Required for Rice Pollen Development. Plant Physiol. 2017, 173, 240–255. [Google Scholar] [CrossRef] [Green Version]
- Cigan, A.M. Targeted Mutagenesis of a Conserved Anther-Expressed P450 Gene Confers Male Sterility in Monocots. Plant Biotechnol. J. 2016, 15, 379–389. [Google Scholar] [CrossRef]
- Kirch, H.H. The ALDH gene superfamily of Arabidopsis. Trends Plant Sci. 2004, 9, 371–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F. Mitochondrial aldehyde dehydrogenase activity is required for male fertility in maize. Plant Cell 2001, 13, 1063–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, G.L.; Wang, X.M. Role of Aminoalcoholphosphotransferases 1 and 2 in Phospholipid Homeostasis in Arabidopsis. Plant Cell 2015, 27, 1512–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vance, J.E. Thematic Review Series: Glycerolipids. Phosphatidylserine and Phosphatidylethanolamine in Mammalian Cells: Two Metabolically Related Aminophospholipids. J. Lipid Res. 2008, 49, 1377–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaoka, Y. Phosphatidylserine synthase1 is required for microspore development in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2011, 67, 648–661. [Google Scholar] [CrossRef]
- Ischebeck, T. Lipids in pollen—They are different. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1315–1328. [Google Scholar] [CrossRef]
- Botté, C.Y. Chemical inhibitors of monogalactosyldiacylglycerol synthases in Arabidopsis thaliana. Nat. Chem. Biol. 2011, 7, 834–842. [Google Scholar] [CrossRef]
- Shi, J. Defective pollen wall is required for anther and microspore development in rice and encodes a fatty acyl carrier protein reductase. Plant Cell 2011, 23, 2225–2246. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, H.; Yu, Y.; Gu, R.; Feng, C.; Fu, Y.; Yu, X.; Yuan, J.; Sun, Q.; Ke, Y. Male sterile 305 Mutation Leads the Misregulation of Anther Cuticle Formation by Disrupting Lipid Metabolism in Maize. Int. J. Mol. Sci. 2020, 21, 2500. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072500
Shi H, Yu Y, Gu R, Feng C, Fu Y, Yu X, Yuan J, Sun Q, Ke Y. Male sterile 305 Mutation Leads the Misregulation of Anther Cuticle Formation by Disrupting Lipid Metabolism in Maize. International Journal of Molecular Sciences. 2020; 21(7):2500. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072500
Chicago/Turabian StyleShi, Haichun, Yang Yu, Ronghuan Gu, Chenxi Feng, Yu Fu, Xuejie Yu, Jichao Yuan, Qun Sun, and Yongpei Ke. 2020. "Male sterile 305 Mutation Leads the Misregulation of Anther Cuticle Formation by Disrupting Lipid Metabolism in Maize" International Journal of Molecular Sciences 21, no. 7: 2500. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072500