LPMO AfAA9_B and Cellobiohydrolase AfCel6A from A. fumigatus Boost Enzymatic Saccharification Activity of Cellulase Cocktail

 , , , , ,
, , , , ,
Abstract
:1. Introduction
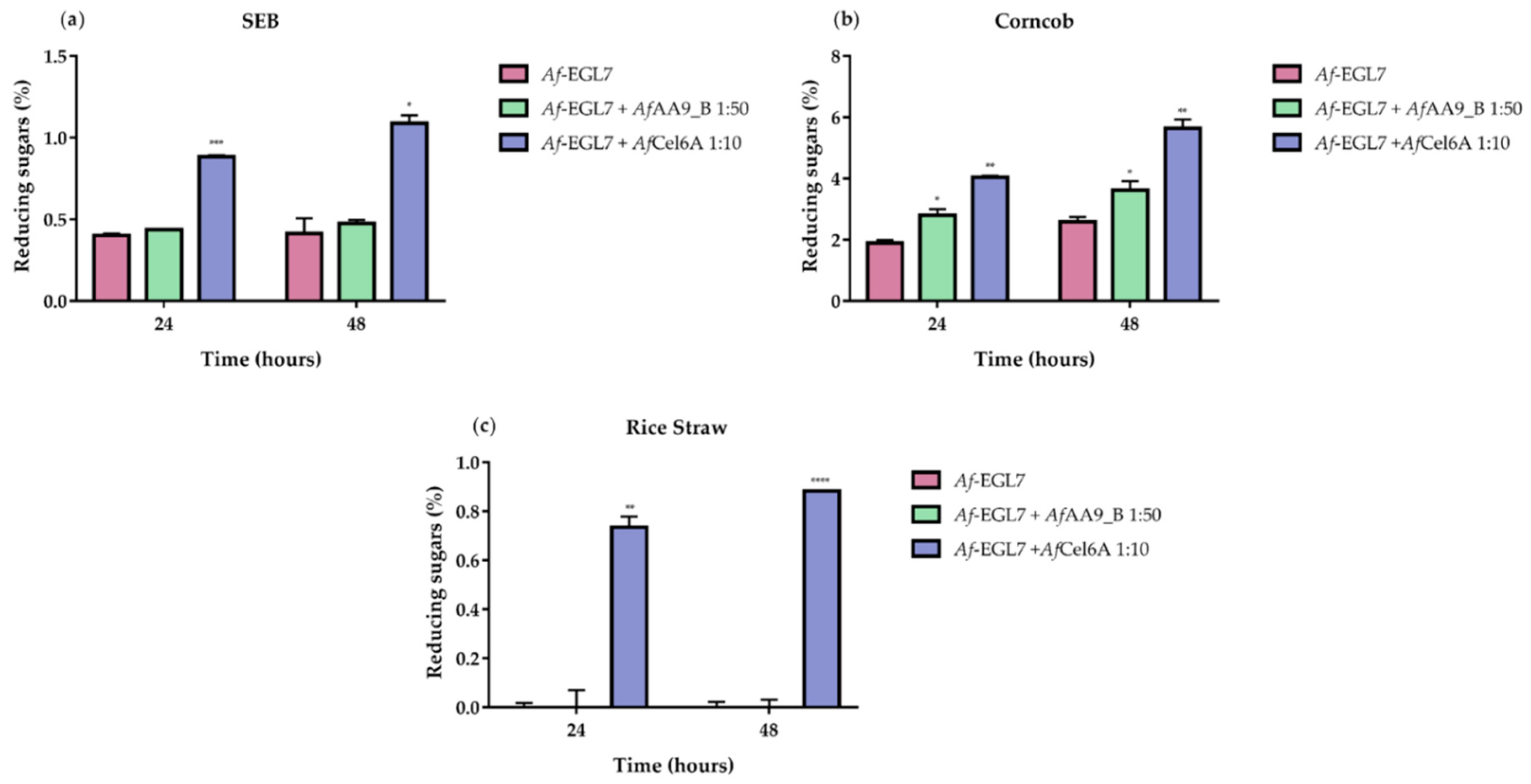
2. Results and Discussion
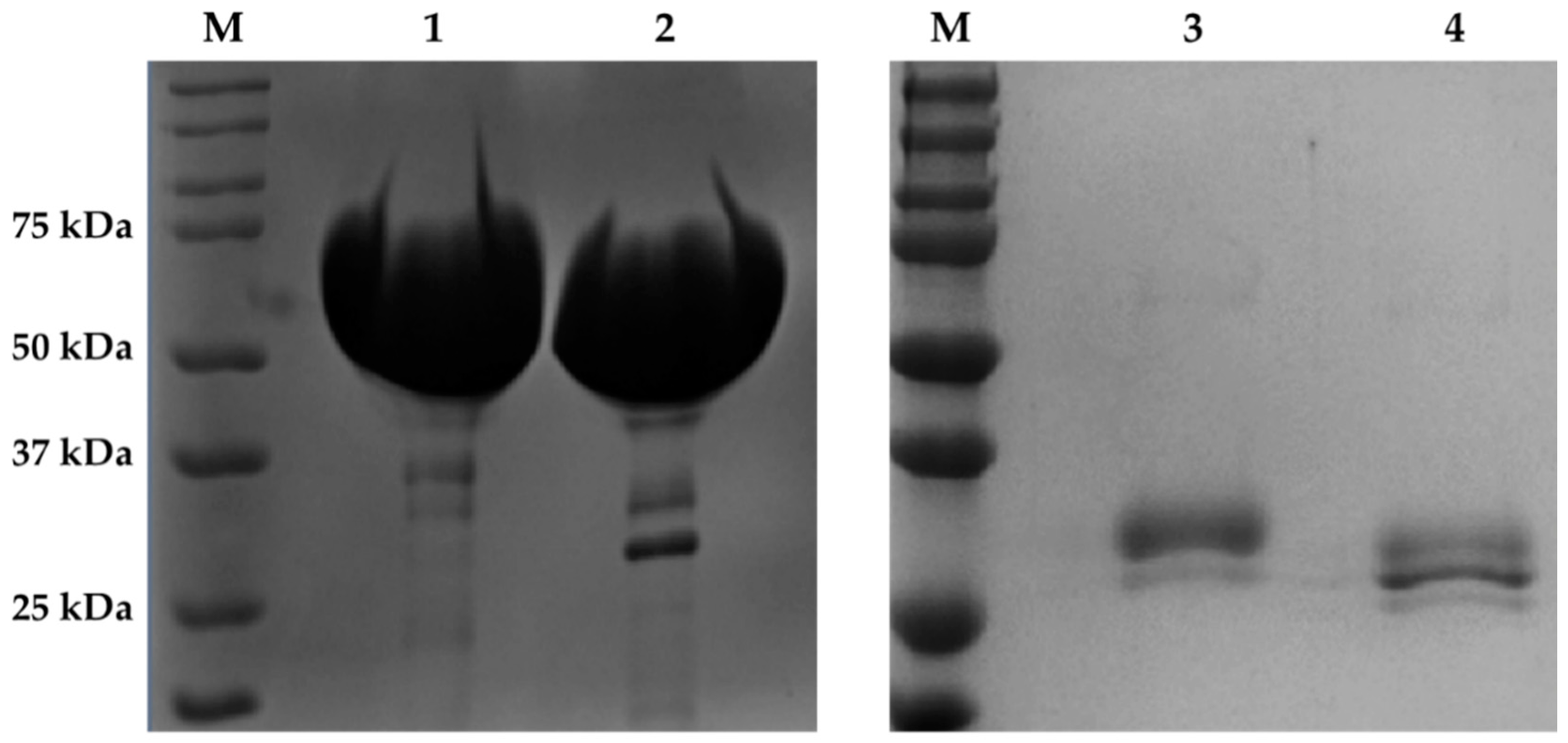
2.1. Expression and Purification of Recombinant AfCel6A and AfAA9_B
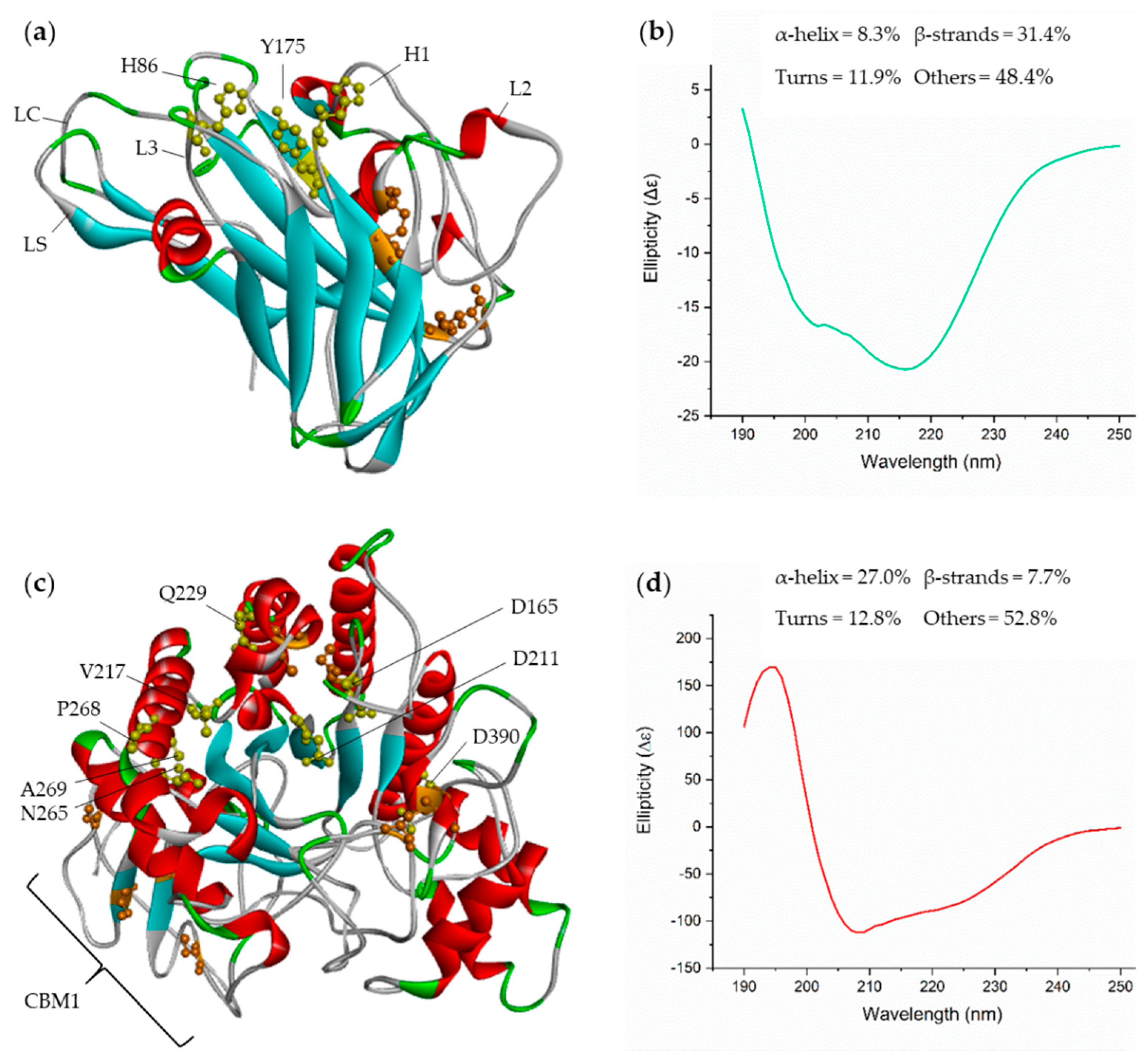
2.2. Structural Analysis and Predictions by Circular Dichroism (CD)
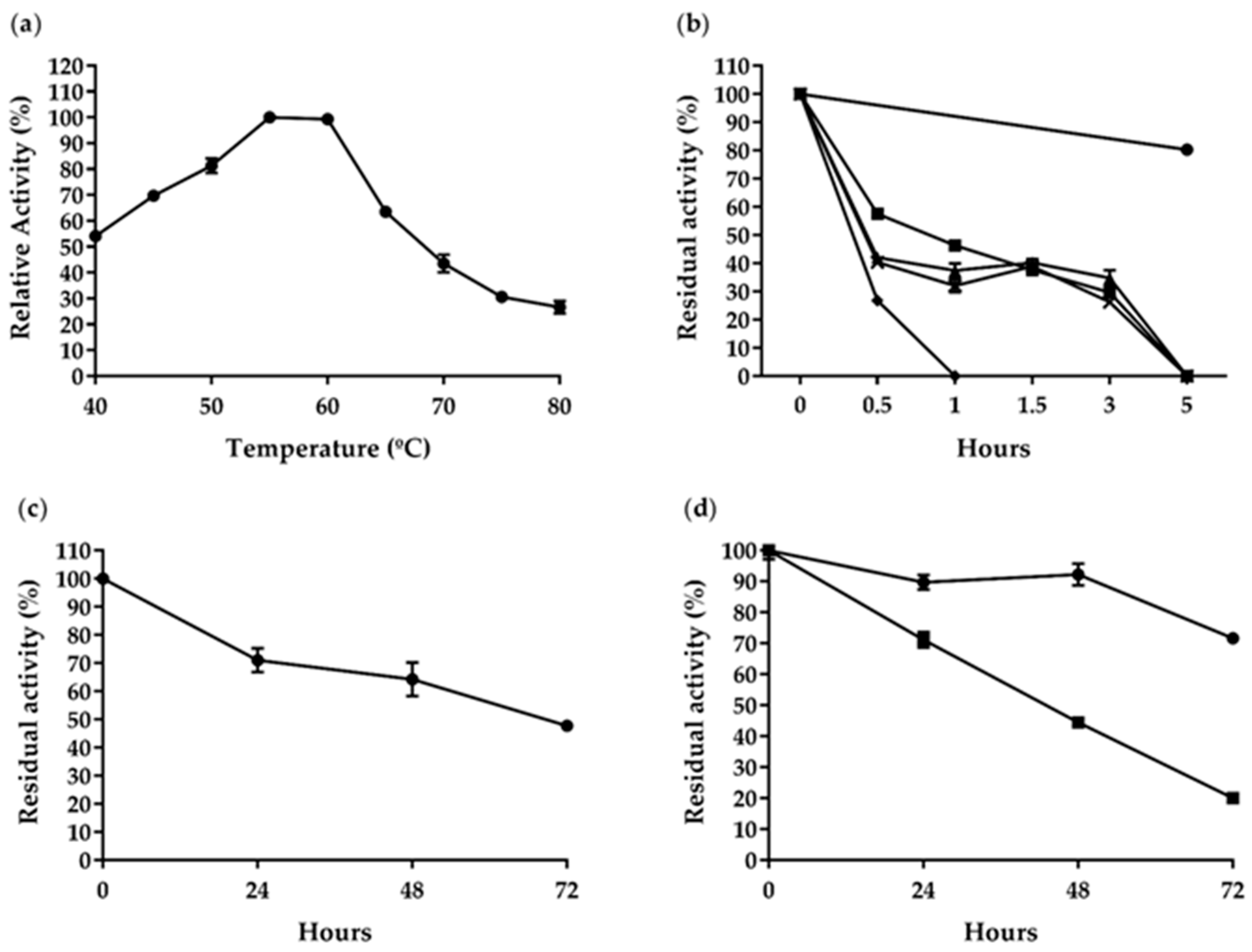
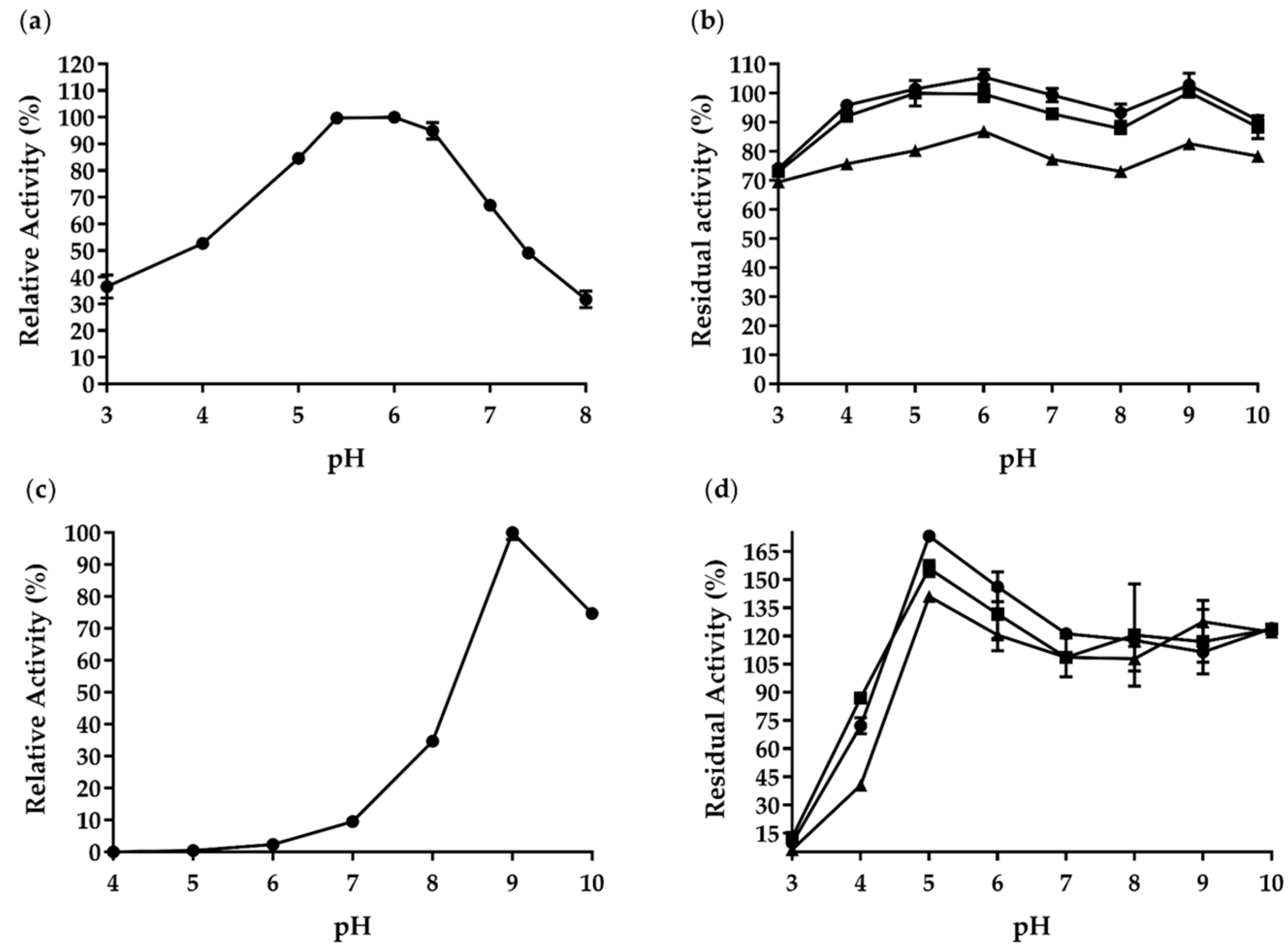
2.3. Enzymatic Properties of AfCel6A and AfAA9_B
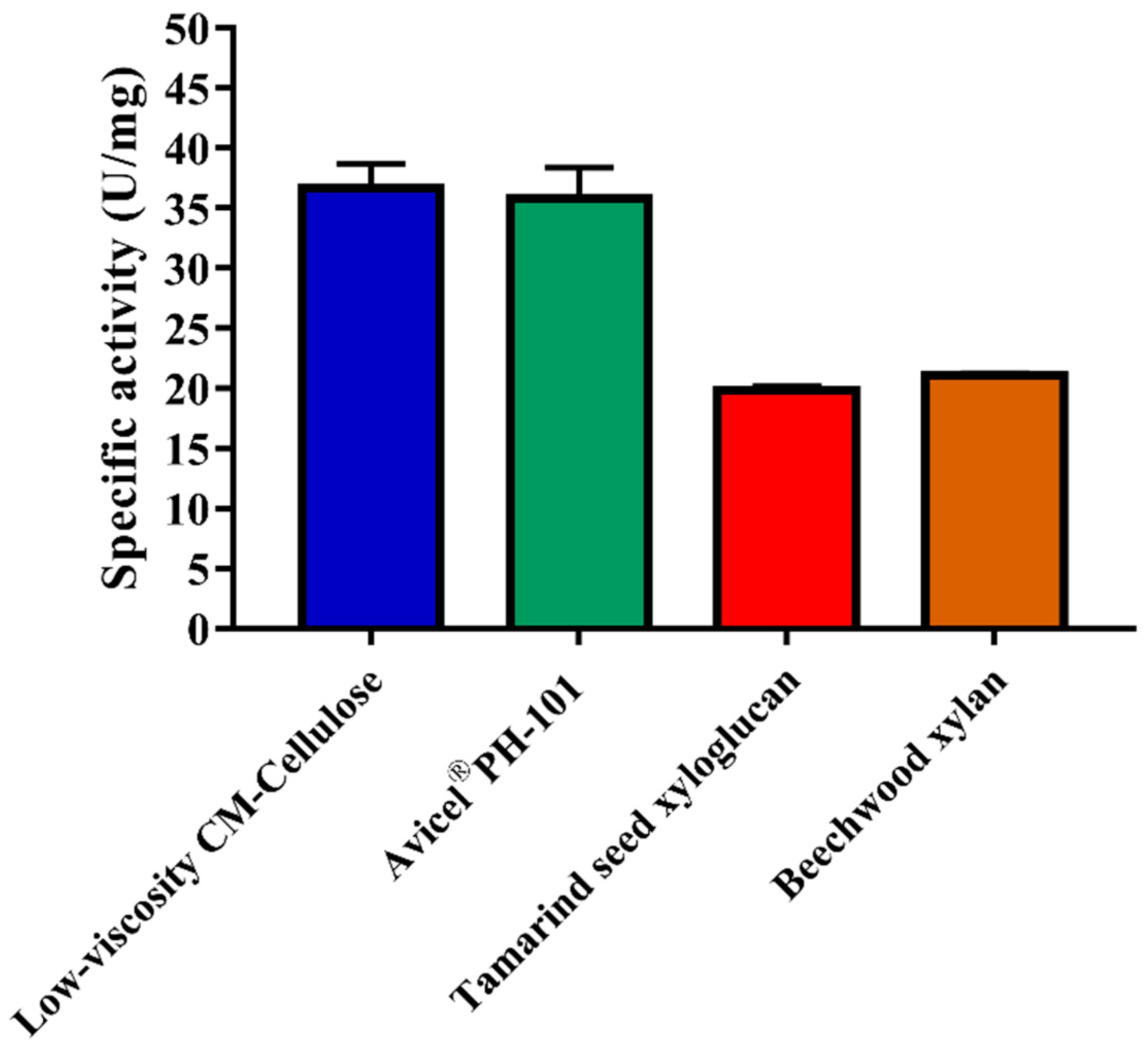
2.4. Substrate Specificity and Kinetic Parameters
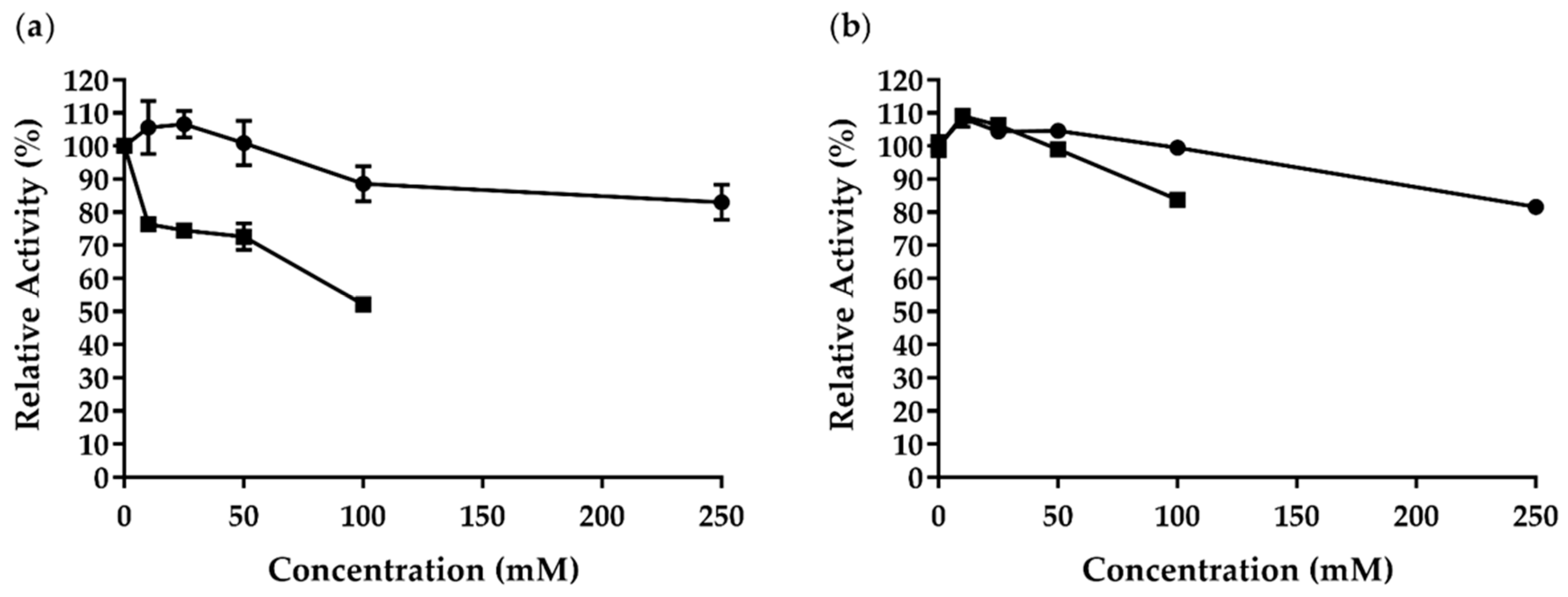
2.5. Effect of Different Metal Ions and Chemicals
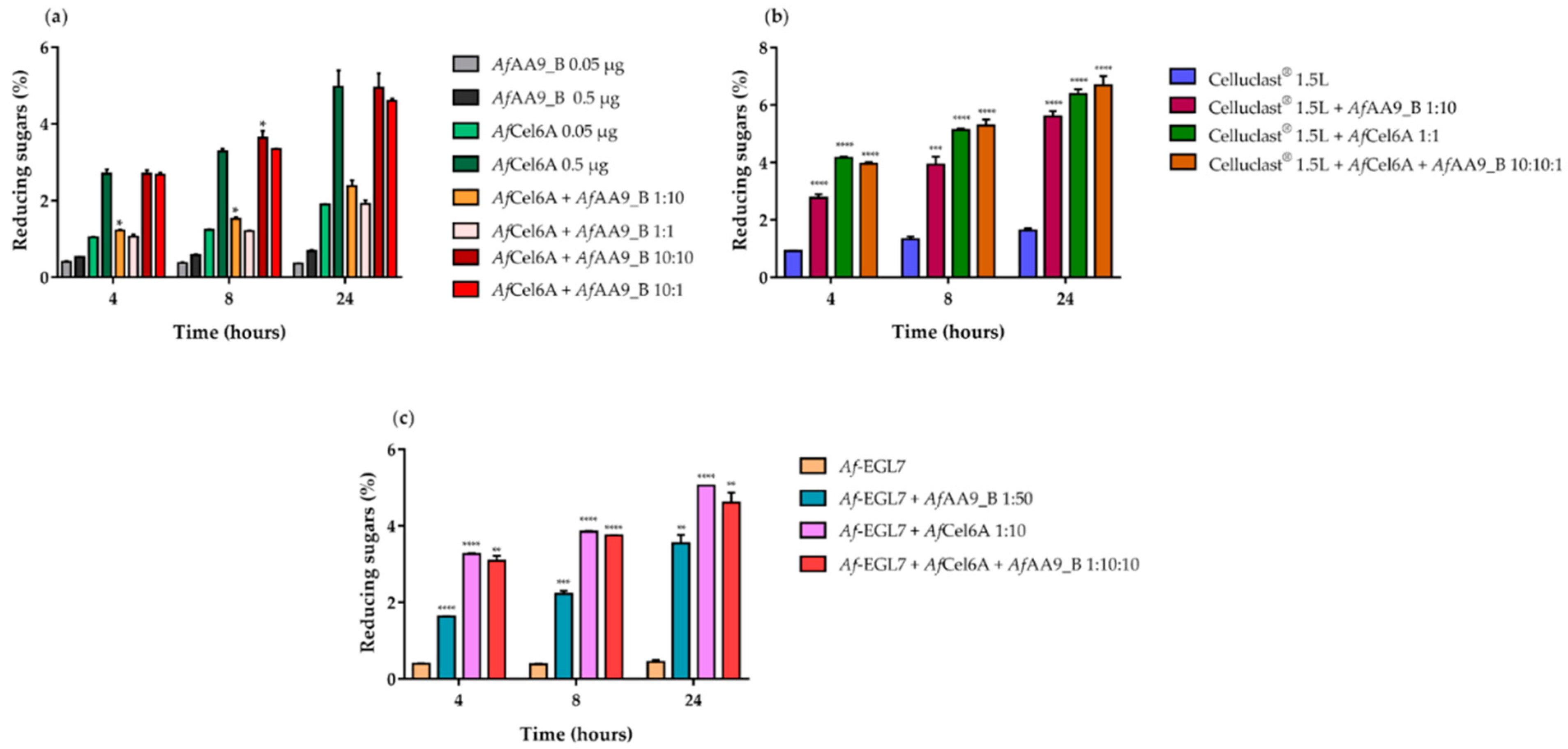
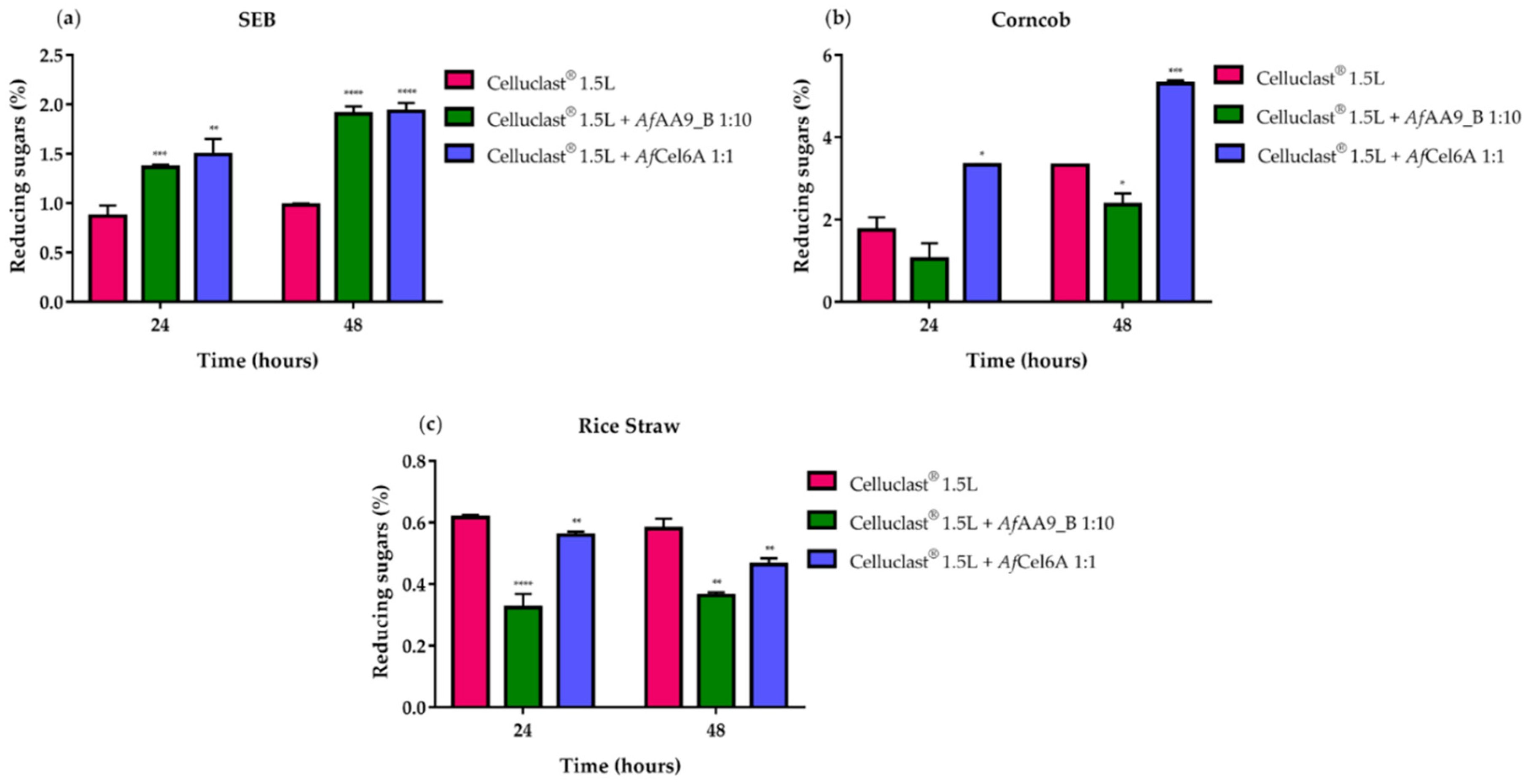
2.6. Synergistic Action on Cellulose Hydrolysis
3. Materials and Methods
3.1. Strains, Culture Conditions, and Vectors
3.2. RNA Extraction, cDNA Synthesis, and Gene Amplification
3.3. Enzyme Production and Purification
3.4. Glycosylation
3.5. Structural Analysis by Circular Dichroism (CD)
3.6. LPMO Activity Assay
3.7. AfCel6A Activity Assay
3.8. Enzymatic Properties of AfAA9_B and AfCel6A
3.9. Effect of Additives
3.10. Glucose and Cellobiose Effects on AfCel6A and AfAA9_B Activity
3.11. Kinetic Assays
3.12. Combined Assays
3.13. Synergistic Activity with Celluclast® 1.5L
3.14. Lignocellulosic Biomass Saccharification
3.15. Reproducibility of the Results
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2G ethanol | Second-generation ethanol |
| LPMO | Lytic Polysaccharide Monooxygenase |
| AA | Auxiliary Activity |
| GH | Glycoside Hydrolase |
| CD | Circular Dichroism |
| CBM | Carbohydrate Binding Module |
| CMC-Na | Sodium Carboxymethyl Cellulose |
| 2,6-DMP | 2,6-Dimethoxyphenol |
| PASC | Phosphoric-acid Swollen Cellulose |
| pNPC | 4(p)-nitrophenyl β-D-cellobioside |
| SEB | Sugarcane Exploded Bagasse |
| DNS | 3,5-Dinitrosalicylic acid |
| FPU | Filter Paper Unit |
References
- El-Tayeb, T.S.; Abdelhafez, A.A.; Ali, S.H.; Ramadan, E.M. Effect of acid hydrolysis and fungal biotreatment on agro-industrial wastes for obtainment of free sugars for bioethanol production. Braz. J. Microbiol. 2012, 43, 1523–1535. [Google Scholar] [PubMed]
- Anwar, M.; Lou, S.; Chen, L.; Li, H.; Hu, Z. Recent advancement and strategy on bio-hydrogen production from photosynthetic microalgae. Bioresour. Technol. 2019, 292, 121972. [Google Scholar] [PubMed]
- Ren, H.Y.; Kong, F.; Zhao, L.; Ren, N.Q.; Ma, J.; Nan, J.; Liu, B.F. Enhanced co-production of biohydrogen and algal lipids from agricultural biomass residues in long-term operation. Bioresour. Technol. 2019, 289, 121774. [Google Scholar]
- Boboescu, I.-Z.; Damay, J.; Chang, J.K.W.; Beigbeder, J.-B.; Duret, X.; Beauchemin, S.; Lalonde, O.; Lavoie, J.-M. Ethanol production from residual lignocellulosic fibers generated through the steam treatment of whole sorghum biomass. Bioresour. Technol. 2019, 292, 121975. [Google Scholar] [PubMed]
- Dhakate, S.R.; Pathak, A.K.; Jain, P.; Singh, M.; Singh, B.P.; Subhedar, K.M.; Sharda, S.S.; Seth, R.K. Rice straw biomass to high energy yield biocoal by torrefaction: Indian perspective. Curr. Sci. 2019, 116, 831–838. [Google Scholar]
- Hill, J.; Polasky, S.; Nelson, E.; Tilman, D.; Huo, H.; Ludwig, L.; Neumann, J.; Zheng, H.; Bonta, D. Climate change and health costs of air emissions from biofuels and gasoline. Proc. Natl. Acad. Sci. USA 2009, 106, 2077–2082. [Google Scholar] [PubMed] [Green Version]
- Mudinoor, A.R.; Goodwin, P.M.; Rao, R.U.; Karuna, N.; Hitomi, A.; Nill, J.; Jeoh, T. Interfacial molecular interactions of cellobiohydrolase Cel7A and its variants on cellulose. Biotechnol. Biofuels 2020, 13, 1–16. [Google Scholar]
- Kucharska, K.; Rybarczyk, P.; Hołowacz, I.; Łukajtis, R.; Glinka, M.; Kamiński, M. Pretreatment of lignocellulosic materials as substrates for fermentation processes. Molecules 2018, 23, 2937. [Google Scholar]
- Ravindran, R.; Jaiswal, A.K. A comprehensive review on pre-treatment strategy for lignocellulosic food industry waste: Challenges and opportunities. Bioresour. Technol. 2016, 199, 92–102. [Google Scholar]
- Nishiyama, Y. Structure and properties of the cellulose microfibril. J. Wood Sci. 2009, 55, 241–249. [Google Scholar]
- Payne, C.M.; Knott, B.C.; Mayes, H.B.; Hansson, H.; Himmel, M.E.; Sandgren, M.; Ståhlberg, J.; Beckham, G.T. Fungal cellulases. Chem. Rev. 2015, 115, 1308–1448. [Google Scholar] [PubMed] [Green Version]
- Medie, F.M.; Davies, G.J.; Drancourt, M.; Henrissat, B. Genome analyses highlight the different biological roles of cellulases. Nat. Rev. Microbiol. 2012, 10, 227–234. [Google Scholar]
- Blumer-Schuette, S.E.; Kataeva, I.; Westpheling, J.; Adams, M.W.; Kelly, R.M. Extremely thermophilic microorganisms for biomass conversion: Status and prospects. Curr. Opin. Biotechnol. 2008, 19, 210–217. [Google Scholar] [PubMed]
- Hu, J.; Chandra, R.; Arantes, V.; Gourlay, K.; van Dyk, J.S.; Saddler, J.N. The addition of accessory enzymes enhances the hydrolytic performance of cellulase enzymes at high solid loadings. Bioresour. Technol. 2015, 186, 149–153. [Google Scholar] [PubMed]
- Hu, J.; Arantes, V.; Pribowo, A.; Saddler, J.N. The synergistic action of accessory enzymes enhances the hydrolytic potential of a “cellulase mixture” but is highly substrate specific. Biotechnol. Biofuels 2013, 6, 112. [Google Scholar] [PubMed] [Green Version]
- Müller, G.; Várnai, A.; Johansen, K.S.; Eijsink, V.G.H.; Horn, S.J. Harnessing the potential of LPMO-containing cellulase cocktails poses new demands on processing conditions. Biotechnol. Biofuels 2015, 8, 1–9. [Google Scholar]
- Garcia-Santamarina, S.; Probst, C.; Festa, R.A.; Ding, C.; Smith, A.D.; Conklin, S.E.; Brander, S.; Kinch, L.N.; Grishin, N.V.; Franz, K.J.; et al. A lytic polysaccharide monooxygenase-like protein functions in fungal copper import and meningitis. Nat. Chem. Biol. 2020, 16, 337–344. [Google Scholar]
- Vaaje-Kolstad, G.; Horn, S.J.; Van Aalten, D.M.F.; Synstad, B.; Eijsink, V.G.H. The non-catalytic chitin-binding protein CBP21 from Serratia marcescens is essential for chitin degradation. J. Biol. Chem. 2005, 280, 28492–28497. [Google Scholar]
- Merino, S.T.; Cherry, J. Progress and challenges in enzyme development for biomass utilization. Adv. Biochem. Eng. Biotechnol. 2007, 108, 95–120. [Google Scholar]
- Vaaje-kolstad, G.; Westereng, B.; Horn, S.J.; Liu, Z.; Zhai, H.; Sørlie, M.; Eijsink, V.G.H. An oxidative enzyme boosting the enzymatic conversion of recalcitrant polysaccharides. Science 2010, 330, 219–222. [Google Scholar]
- Frandsen, K.E.H.; Tovborg, M.; Jørgensen, C.I.; Spodsberg, N.; Rosso, M.N.; Hemsworth, G.R.; Garman, E.F.; Grime, G.W.; Poulsen, J.C.N.; Batth, T.S.; et al. Insights into an unusual Auxiliary Activity 9 family member lacking the histidine brace motif of lytic polysaccharide monooxygenases. J. Biol. Chem. 2019, 294, 17117–17130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertini, L.; Breglia, R.; Lambrughi, M.; Fantucci, P.; De Gioia, L.; Borsari, M.; Sola, M.; Bortolotti, C.A.; Bruschi, M. Catalytic Mechanism of Fungal Lytic Polysaccharide Monooxygenases Investigated by First-Principles Calculations. Inorg. Chem. 2018, 57, 86–97. [Google Scholar] [CrossRef]
- Dimarogona, M.; Topakas, E.; Christakopoulos, P. Cellulose degradation by oxidative enzymes. Comput. Struct. Biotechnol. J. 2012, 2, e201209015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaaje-Kolstad, G.; Forsberg, Z.; Loose, J.S.; Bissaro, B.; Eijsink, V.G. Structural diversity of lytic polysaccharide monooxygenases. Curr. Opin. Struct. Biol. 2017, 44, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, G.R.; Henrissat, B.; Davies, G.J.; Walton, P.H. Discovery and characterization of a new family of lytic polysaccharide monooxygenases. Nat. Chem. Biol. 2014, 10, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, R.J.; Sweeney, M.D.; Leggio, L.L.; Otten, H.; Poulsen, J.-C.N.; Johansen, K.S.; Krogh, K.B.R.M.; Jorgensen, C.I.; Tovborg, M.; Anthonsen, A.; et al. Insights into the oxidative degradation of cellulose by a copper metalloenzyme that exploits biomass components. Proc. Natl. Acad. Sci. USA 2011, 108, 15079–15084. [Google Scholar] [CrossRef] [Green Version]
- Eijsink, V.G.H.; Petrovic, D.; Forsberg, Z.; Mekasha, S.; Røhr, Å.K.; Várnai, A.; Bissaro, B.; Vaaje-Kolstad, G. On the functional characterization of lytic polysaccharide monooxygenases (LPMOs). Biotechnol. Biofuels 2019, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Bissaro, B.; Røhr, Å.K.; Müller, G.; Chylenski, P.; Skaugen, M.; Forsberg, Z.; Horn, S.J.; Vaaje-Kolstad, G.; Eijsink, V.G.H. Oxidative cleavage of polysaccharides by monocopper enzymes depends on H2O2. Nat. Chem. Biol. 2017, 13, 1123–1128. [Google Scholar] [CrossRef]
- Walton, P.H.; Davies, G.J. On the catalytic mechanisms of lytic polysaccharide monooxygenases. Curr. Opin. Chem. Biol. 2016, 31, 195–207. [Google Scholar] [CrossRef]
- Danneels, B.; Tanghe, M.; Desmet, T. Structural Features on the Substrate-Binding Surface of Fungal Lytic Polysaccharide Monooxygenases Determine Their Oxidative Regioselectivity. Biotechnol. J. 2019, 14, e1800211. [Google Scholar] [CrossRef]
- Eibinger, M.; Ganner, T.; Bubner, P.; Rošker, S.; Kracher, D.; Haltrich, D.; Ludwig, R.; Plank, H.; Nidetzky, B. Cellulose surface degradation by a lytic polysaccharide monooxygenase and its effect on cellulase hydrolytic efficiency. J. Biol. Chem. 2014, 289, 35929–35938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardi, A.V.; Yonamine, D.K.; Uyemura, S.A.; Dinamarco, T.M. A Thermostable Aspergillus fumigatus GH7 Endoglucanase Over-Expressed in Pichia pastoris Stimulates Lignocellulosic Biomass Hydrolysis. Int. J. Mol. Sci. 2019, 20, 2261. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Zhang, A.; Ludeña, Y.; Villena, G.K.; Yu, F.; Sherman, D.H.; Gutiérrez-Correa, M. Insights from the genome of a high alkaline cellulase producing Aspergillus fumigatus strain obtained from Peruvian Amazon rainforest. J. Biotechnol. 2017, 251, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Zhang, R.; Yang, X.; Wu, H.; Xu, D.; Tang, Z.; Shen, Q. Thermostable cellulase production of Aspergillus fumigatus Z5 under solid-state fermentation and its application in degradation of agricultural wastes. Int. Biodeterior. Biodegrad. 2011, 65, 717–725. [Google Scholar] [CrossRef]
- Christensen, S.J.; Bertel, K.; Mørkeberg, R.; Spodsberg, N.; Borch, K.; Westh, P. A biochemical comparison of fungal GH6 cellobiohydrolases. Biochem. J. 2019, 6, 2157–2172. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, A.V.; De Gouvêa, P.F.; Gerolamo, L.E.; Yonamine, D.K.; De Lourdes, L.; Balico, D.L.; Uyemura, S.A. Functional characterization of GH7 endo-1,4-β-glucanase from Aspergillus fumigatus and its potential industrial application. Protein Expr. Purif. 2018, 150, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Adsul, M.G.; Gokhale, D.V. The conundrum of making biomass-to-biofuels economic. Biofuels 2012, 3, 383–386. [Google Scholar] [CrossRef]
- Jin, M.; Sousa, L.D.C.; Schwartz, C.; He, Y.; Sarks, C.; Gunawan, C.; Balan, V.; Dale, B.E. Toward lower cost cellulosic biofuel production using ammonia based pretreatment technologies. Green Chem. 2016, 18, 957–966. [Google Scholar] [CrossRef]
- Chandel, A.K.; Chandrasekhar, G.; Silva, M.B.; Silva, S.D.S. The realm of cellulases in biorefinery development. Crit. Rev. Biotechnol. 2012, 32, 187–202. [Google Scholar] [CrossRef]
- Puri, D.J.; Heaven, S.; Banks, C.J. Improving the performance of enzymes in hydrolysis of high solids paper pulp derived from MSW. Biotechnol. Biofuels 2013, 6, 107. [Google Scholar] [CrossRef] [Green Version]
- Adsul, M.; Sandhu, S.K.; Singhania, R.R.; Gupta, R.; Puri, S.K. Enzyme and Microbial Technology Designing a cellulolytic enzyme cocktail for the e ffi cient and economical conversion of lignocellulosic biomass to biofuels. Enzyme Microb. Technol. 2020, 133, 109442. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.; Tchelet, R. Next Generation Enzymes. Biofuels Int. 2014, 12, 49–50. [Google Scholar]
- De Gouvêa, P.F.; Gerolamo, L.E.; Bernardi, A.V.; Matheus, L.; Pereira, S.; Uyemura, S.A.; Dinamarco, T.M. Lytic Polysaccharide Monooxygenase from Aspergillus fumigatus can Improve Enzymatic Cocktail Activity During Sugarcane Bagasse Hydrolysis. Protein Pept. Lett. 2019, 126, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Rubio, M.V.; Zubieta, M.P.; Cairo, J.P.L.F.; Calzado, F.; Leme, A.F.P.; Squina, F.M.; Prade, R.A.; Damásio, A.R.D.L. Mapping N-linked glycosylation of carbohydrate-active enzymes in the secretome of Aspergillus nidulans grown on lignocellulose. Biotechnol. Biofuels 2016, 9, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Bissaro, B.; Kommedal, E.; Røhr, Å.K.; Eijsink, V.G.H. Controlled depolymerization of cellulose by light-driven lytic polysaccharide oxygenases. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Chylenski, P.; Petrović, D.M.; Müller, G.; Dahlström, M.; Bengtsson, O.; Lersch, M.; Siika-Aho, M.; Horn, S.J.; Eijsink, V.G.H. Enzymatic degradation of sulfite-pulped softwoods and the role of LPMOs. Biotechnol. Biofuels 2017, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Courtade, G.; Wimmer, R.; Røhr, Å.K.; Preims, M.; Felice, A.K.G.; Dimarogona, M.; Vaaje-Kolstad, G.; Sørlie, M.; Sandgren, M.; Ludwig, R.; et al. Interactions of a fungal lytic polysaccharide monooxygenase with β-glucan substrates and cellobiose dehydrogenase. Proc. Natl. Acad. Sci. USA 2016, 113, 5922–5927. [Google Scholar] [CrossRef] [Green Version]
- Dodda, S.R.; Sarkar, N.; Aikat, K.; Krishnaraj, N.R.; Bhattacharjee, S.; Bagchi, A.; Mukhopadhyay, S. Insights from the Molecular Dynamics Simulation of Cellobiohydrolase Cel6A Molecular Structural Model from Aspergillus fumigatus NITDGPKA3. Comb. Chem. High Throughput Screen. 2016, 19, 325–333. [Google Scholar] [CrossRef]
- Thompson, A.J.; Heu, T.; Shaghasi, T.; Jones, A.; Friis, E.P.; Wilson, K.S.; Davies, G.J. research papers Structure of the catalytic core module of the Chaetomium thermophilum family GH6 cellobiohydrolase Cel6A. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 875–882. [Google Scholar] [CrossRef]
- Mertz, B.; Kuczenski, R.S.; Larsen, R.T.; Hill, A.D.; Reilly, P.J. Phylogenetic Analysis of Family 6 Glycoside Hydrolases. Biopolymers 2005, 79, 197–206. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.L.; Eddy, S.R.; Durbin, R. Pfam: A comprehensive database of protein domain families based on seed alignments. Proteins Struct. Funct. Genet. 1997, 28, 405–420. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. et Biophys. Acta (BBA) Proteins Proteom. 2005, 1751, 119–139. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Bulyáki, É.; Kun, J.; Moussong, É.; Lee, Y.-H.; Goto, Y.; Réfrégiers, M.; Kardos, J. BeStSel: A web server for accurate protein secondary structure prediction and fold recognition from the circular dichroism spectra. Nucleic Acids Res. 2018, 46, W315–W322. [Google Scholar] [CrossRef] [PubMed]
- Lo Leggio, L.; Weihe, C.D.; Poulsen, J.C.N.; Sweeney, M.; Rasmussen, F.; Lin, J.; De Maria, L.; Wogulis, M. Structure of a lytic polysaccharide monooxygenase from Aspergillus fumigatus and an engineered thermostable variant. Carbohydr. Res. 2018, 469, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, B.A.; Janes, R.W. Modern Techniques for Circular Dichroism and Synchrotron Radiation Circular Dichroism Spectroscopy; IOS Press: Amsterdam, NY, USA, 2009; Volume 1, ISBN 9781607500001. [Google Scholar]
- Kabsch, W.; Sander, C. Dictionary of protein secondary structure: Pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 1983, 22, 2577–2637. [Google Scholar] [CrossRef] [PubMed]
- Teugjas, H.; Väljamäe, P. Product inhibition of cellulases studied with 14C-labeled cellulose substrates. Biotechnol. Biofuels 2013, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Fuelbiol, F.; Bai, R.; Yang, H.; Wang, F.; He, J.; Wang, C.; Tu, M. Enzyme and Microbial Technology Heterologous expression of codon optimized Trichoderma reesei Cel6A in Pichia pastoris. Enzyme Microb. Technol. 2016, 92, 107–116. [Google Scholar]
- Boonvitthya, N.; Bozonnet, S. Comparison of the Heterologous Expression of Trichoderma reesei Endoglucanase II and Cellobiohydrolase II in the Yeasts Pichia pastoris and Yarrowia lipolytica. Mol. Biotechnol. 2013, 54, 158–169. [Google Scholar] [CrossRef]
- Okada, H.; Sekiya, T.; Yokoyama, K.; Tohda, H.; Kumagai, H.; Morikawa, Y. Efficient secretion of Trichoderma reesei cellobiohydrolase II in Schizosaccharomyces pombe and characterization of its products. Appl. Microbiol. Biotechnol. 1998, 49, 301–308. [Google Scholar] [CrossRef]
- Takahashi, M.; Takahashi, H.; Nakano, Y.; Konishi, T.; Terauchi, R.; Takeda, T. Characterization of a Cellobiohydrolase ( MoCel6A) Produced by Magnaporthe oryzae. Appl. Environ. Microbiol. 2010, 76, 6583–6590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riyadh, S.K.; Vipin, I.A.I.K.; Kalia, C.; Lee, J. Characterization of Cellobiohydrolases from Schizophyllum commune KMJ820. Indian J. Microbiol. 2020, 60, 160–166. [Google Scholar]
- Limam, F.; Chaabouni, S.E.; Ghrir, R.; Marzouki, N. Two cellobiohydrolases of Penicillium occitanis mutant Pol 6: Purification and properties. Enzym. Microb. Technol. 1995, 0229, 340–346. [Google Scholar] [CrossRef]
- Tuohy, M.G.; Walsh, D.J.; Murray, P.G.; Claeyssens, M.; Cuffe, M.M.; Savage, A.V.; Coughlan, M.P. Kinetic parameters and mode of action of the cellobiohydrolases produced by Talaromyces emersonii. Biochim. et Biophys. Acta (BBA) Protein Struct. Mol. Enzym. 2002, 1596, 366–380. [Google Scholar] [CrossRef]
- Song, J.; Liu, B.; Liu, Z. Cloning of two cellobiohydrolase genes from Trichoderma viride and heterogenous expression in yeast Saccharomyces cerevisiae. Mol. Biol. Rep. 2010, 37, 2135–2140. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.; Huang, C.; Hseu, R. Cloning and characterization of a thermostable and pH-stable cellobiohydrolase from Neocallimastix patriciarum J11. Protein Expr. Purif. 2013, 90, 153–159. [Google Scholar] [CrossRef]
- Toda, H.; Nagahata, N.; Amano, Y.; Nozaki, K.; Kanda, T.; Okazaki, M.; Shimosaka, M. Gene Cloning of Cellobiohydrolase II from the White Rot Fungus Irpex lacteus MC-2 and Its Expression in Pichia pastoris. Biosci. Biotechnol. Biochem. 2008, 72, 3142–3147. [Google Scholar] [CrossRef]
- Wang, X.; Peng, Y.; Zhang, L. Directed evolution and structural prediction of cellobiohydrolase II from the thermophilic fungus Chaetomium thermophilum. Appl. Microbiol. Biotechnol. 2012, 95, 1469–1478. [Google Scholar] [CrossRef]
- Agrawal, D.; Basotra, N.; Balan, V.; Tsang, A.; Chadha, B.S. Discovery and Expression of Thermostable LPMOs from Thermophilic Fungi for Producing Efficient Lignocellulolytic Enzyme Cocktails. Appl. Biochem. Biotechnol. 2019, 191, 463–481. [Google Scholar] [CrossRef]
- Keller, M.B.; Felby, C.; Labate, C.A.; Pellegrini, V.O.A.; Higasi, P.; Singh, R.K.; Polikarpov, I.; Blossom, B.M. A simple enzymatic assay for the quantification of C1-specific cellulose oxidation by lytic polysaccharide monooxygenases. Biotechnol. Lett. 2020, 42, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Hangasky, J.A.; Iavarone, A.T.; Marletta, M.A. Reactivity of O 2 versus H 2 O 2 with polysaccharide monooxygenases. Proc. Natl. Acad. Sci. USA 2018, 115, 4915–4920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsberg, Z.; Sørlie, M.; Petrović, D.; Courtade, G.; Aachmann, F.L.; Vaaje-Kolstad, G.; Bissaro, B.; Røhr, Å.K.; Eijsink, V.G. Polysaccharide degradation by lytic polysaccharide monooxygenases. Curr. Opin. Struct. Biol. 2019, 59, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Kuusk, S.; Bissaro, B.; Kuusk, P.; Forsberg, Z.; Eijsink, V.G.H.; Sørlie, M.; Valjamae, P. Kinetics of H2O2-driven degradation of chitin by a bacterial lytic polysaccharide monooxygenase. J. Biol. Chem. 2018, 293, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frandsen, K.E.H.; Simmons, T.J.; Dupree, P.; Poulsen, J.C.N.; Hemsworth, G.R.; Ciano, L.; Johnston, E.M.; Tovborg, M.; Johansen, K.S.; Von Freiesleben, P.; et al. The molecular basis of polysaccharide cleavage by lytic polysaccharide monooxygenases. Nat. Chem. Biol. 2016, 12, 298–303. [Google Scholar] [CrossRef] [Green Version]
- Musaddique, H.; Reddy Dodda, S.; Kapoor, B.S.; Aikat, K.; Mukhopadhyay, S.S. Investigation the biomass conversion efficiency, biochemical characterisation and structural insights of the newly isolated AA16 family of Lytic Polysaccharide Monooxygenase (LPMO) from Aspergillus fumigatus. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Frommhagen, M.; Westphal, A.H.; Hilgers, R.; Koetsier, M.J.; Hinz, S.W.A.; Visser, J.; Gruppen, H.; van Berkel, W.J.H.; Kabel, M.A. Quantification of the catalytic performance of C1-cellulose-specific lytic polysaccharide monooxygenases. Appl. Microbiol. Biotechnol. 2018, 102, 1281–1295. [Google Scholar] [CrossRef] [Green Version]
- Kadowaki, M.A.S.; Várnai, A.; Jameson, J.K.; Leite, A.E.T.; Costa-Filho, A.J.; Kumagai, P.S.; Prade, R.A.; Polikarpov, I.; Eijsink, V.G.H. Functional characterization of a lytic polysaccharide monooxygenase from the thermophilic fungus Myceliophthora thermophila. PLoS ONE 2018, 13, e0202148. [Google Scholar] [CrossRef]
- Singh, R.K.; Blossom, B.M.; Russo, D.A.; Van Oort, B.; Croce, R.; Jensen, P.E.; Felby, C.; Bjerrum, M.J. Thermal unfolding and refolding of a lytic polysaccharide monooxygenase from: Thermoascus aurantiacus. RSC Adv. 2019, 9, 29734–29742. [Google Scholar] [CrossRef] [Green Version]
- Breslmayr, E.; Hanžek, M.; Hanrahan, A.; Leitner, C.; Kittl, R.; Šantek, B.; Oostenbrink, C.; Ludwig, R. A fast and sensitive activity assay for lytic polysaccharide monooxygenase. Biotechnol. Biofuels 2018, 11, 79. [Google Scholar] [CrossRef]
- Meleiro, L.P.; Carli, S.; Fonseca-Maldonado, R.; Torricillas, M.d.S.; Zimbardi, A.L.R.L.; Ward, R.J.; Jorge, J.A.; Furriel, R.P.M. Overexpression of a Cellobiose-Glucose-Halotolerant Endoglucanase from Scytalidium thermophilum. Appl. Biochem. Biotechnol. 2018, 185, 316–333. [Google Scholar] [CrossRef]
- Atreya, M.E.; Strobel, K.L.; Clark, D.S. Alleviating product inhibition in cellulase enzyme Cel7A. Biotechnol. Bioeng. 2016, 113, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostylev, M.; Wilson, D. A distinct model of synergism between a processive endocellulase (TfCel9A) and an exocellulase (TfCel9A) from Thermobifida fusca. Appl. Environ. Microbiol. 2014, 80, 339–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, B.; Li, B.; Wang, X.; Shen, W.; Park, S.; Collings, C.; Feng, A.; Smith, S.J.; Walton, J.D.; Ding, S.Y. Real-time imaging reveals that lytic polysaccharide monooxygenase promotes cellulase activity by increasing cellulose accessibility. Biotechnol. Biofuels 2018, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Arantes, V.; Pribowo, A.; Gourlay, K.; Saddler, J.N.; Stephen, J.D.; Mabee, W.E.; Saddler, J.N.; Hu, J.; Arantes, V.; et al. Substrate factors that influence the synergistic interaction of AA9 and cellulases during the enzymatic hydrolysis of biomass. Energy Environ. Sci. 2014, 7, 2308. [Google Scholar] [CrossRef]
- Tokin, R.; Ipsen, J.Ø.; Westh, P.; Johansen, K.S. The synergy between LPMOs and cellulases in enzymatic saccharification of cellulose is both enzyme- and substrate-dependent. Biotechnol. Lett. 2020, 42, 1975–1984. [Google Scholar] [CrossRef]
- Quan, J.; Tian, J. Circular polymerase extension cloning for high-throughput cloning of complex and combinatorial DNA libraries. Nat. Protoc. 2011, 6, 242–251. [Google Scholar] [CrossRef]
- Greenberg, C.S.; Craddock, P.R. Rapid single-step membrane protein assay. Clin. Chem. 1982, 28, 1725–1726. [Google Scholar] [CrossRef]
- Shevchenko, A.; Jensen, O.N.; Podtelejnikov, A.V.; Sagliocco, F.; Wilm, M.; Vorm, O.; Mortensen, P.; Shevchenko, A.; Boucherie, H.; Mann, M. Linking genome and proteome by mass spectrometry: Large-scale identification of yeast proteins from two dimensional gels. Proc. Natl. Acad. Sci. USA 1996, 93, 14440–14445. [Google Scholar] [CrossRef] [Green Version]
- BIOVIA Discovery Studio Visualisation; Accelrys Software Inc.: San Diego, CA, USA, 2017.
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Gene | Peptide Sequence | Precursor MZ | Precursor Charge | Product MZ | Product Charge |
|---|---|---|---|---|---|
| AFUA_4G07850 | ITSIAGLLASASLVAGHGFVSGIVADGK | 871.675226 | 3 | 1049.562586 | 1 |
| AFUA_4G07850 | ITSIAGLLASASLVAGHGFVSGIVADGK | 871.675226 | 3 | 992.541122 | 1 |
| AFUA_4G07850 | ITSIAGLLASASLVAGHGFVSGIVADGK | 871.675226 | 3 | 845.472708 | 1 |
| AFUA_4G07850 | ITSIAGLLASASLVAGHGFVSGIVADGK | 871.675226 | 3 | 746.404294 | 1 |
| AFUA_4G07850 | ITSIAGLLASASLVAGHGFVSGIVADGK | 871.675226 | 3 | 659.372266 | 1 |
| AFUA_4G07850 | ITSIAGLLASASLVAGHGFVSGIVADGK | 871.675226 | 3 | 602.350802 | 1 |
| AFUA_4G07850 | NTDPGIK | 372.912411 | 2 | 630.345717 | 1 |
| AFUA_4G07850 | NTDPGIK | 372.912411 | 2 | 529.298038 | 1 |
| AFUA_4G07850 | NTDPGIK | 372.912411 | 2 | 414.271095 | 1 |
| AFUA_4G07850 | NTDPGIK | 372.912411 | 2 | 317.218332 | 1 |
| Source Organism | Enzyme Name | Expression System | Substrate | Vmax | KM | kcat | kcat/KM | Optimal T | Optimal pH | Thermal Stability | pH Stability | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aspergillus fumigatus Af293 | AfCel6A | Pichia pastoris | CMC-Na | 195.2 ± 4.65 U mg−1 | 7.44 ± 0.51 g/L | 147.9 s−1 | 19.9 mL mg−1 s−1 | 55–60 °C | pH 5.5–6.5 | >70% after 24 h at 50 °C; about 40% after 90 min at 60–80 °C; more than 25% after 30 min at 90 °C | More than 70% at pH 3.0–10.0 after 72 h | This study |
| Aspergillus fumigatus | AfCel6A | Aspergillus oryzae | Avicel PH101 | - | 48.6 ± 14.8 g L−1 | 0.9 ± 0.1 s−1 | - | 70 °C | - | No loss at 60 °C after 1 h | - | [35] |
| Aspergillus terreus | AtCel6A | Aspergillus oryzae | Avicel PH101 | - | - | - | - | 50 °C | - | >90% after 1 h at 50 °C | - | [35] |
| Talaromyces funiculosus | TfCel6A | Aspergillus oryzae | Avicel PH101 | - | 21.6 ± 3.2 g L−1 | 0.5 ± 0.02 s−1 | - | 60 °C | - | No loss at 50 °C after 1 h | - | [35] |
| Colletotrichum graminicola | CgCel6A | Aspergillus oryzae | Avicel PH101 | - | - | - | - | 40 ° C | - | >90% after 1 h at 40 °C | - | [35] |
| CgCel6B | 89.0 ± 13.2 g L−1 | 1.8 ± 0.2 s−1 | 50 °C | >90% after 1 h at 50 °C | ||||||||
| Trichoderma reesei | TrCel6A | Aspergillus oryzae | Avicel PH101 | - | 24.3 ± 4.0 g L−1 | 0.6 ± 0.04 s−1 | - | 70 °C | - | No loss at 50 °C after 1 h | - | [35] |
| Cel6A1 | Pichia pastoris | CMC-Na | 10.7 mmol min−1 mg−1 | 0.31 mg mL−1 | - | - | 60 °C | pH 5.5 | 90% after 30 min at 60 °C | - | [59] | |
| Cel6A2 | Pichia pastoris | PASC | - | - | - | - | 55 °C | pH 5.5–6.0 | 100% at 40 °C and 50% at 60 °C, after 30 min | No loss at pH 5.0–6.0; rapid inactivation at more alkaline pH; and some instability at more acidic pH after 30 min | [60] | |
| CBHII | - | PASC | 10 U mg−1 | 3.8 mg mL−1 | - | - | 60 °C | pH 5.0 | 80% after 30 min at 60 °C | Stable at pH 3.5–6.0 after 30 min | [61] | |
| Magnaporthe oryzae Ina72 | MoCel6A | Magnaporthe oryzae | Cellotetraose | 454.5 µg min−1 mg−1 | 24.3 mM | - | - | 40 °C | pH 9.0 | - | - | [62] |
| Cellopentaose | 63.3 µg min−1 mg−1 | 3.3 mM | ||||||||||
| Schizophyllum commune KMJ820 | CBH II | Escherichia coli | pNPC | 20.8 U mg−1 | 1.4 mM | - | - | 50 °C | pH 5.0 | - | - | [63] |
| Penicillium occitanis Pol 6 | CBH II | - | pNPC | - | 5 mM | - | - | 65 °C | pH 4.0–5.0 | Almost 100% at 30–50 °C; 50% at 60 °C; and complete inactivation at 70 °C, after 30 min | Stable at pH 2.0–9.0 after 24 h | [64] |
| Talaromyces emersonii | CBH II | - | CNPG3 | 9.1 U mg−1 | 4.5 mM | 8.9 s−1 | 1.9 mM−1 s−1 | 68 °C | pH 3.8 | t1/2 = 38 min at 80 °C (pH 5.0) | t1/2 = 38 min at pH 5.0 (80 °C) | [65] |
| Trichoderma viride CICC13038 | CBH II | Saccharomyces cerevisiae | CMC-Na | - | - | - | - | 70 °C | pH 5.0 | - | - | [66] |
| Neocallimastix patriciarum J11 | J11 CelA | Escherichia coli | Barley β-glucan | - | - | - | - | 50 °C | pH 6.0 | More than 70% at up to 50 °C and approximately 50% at 70 °C, after 1 h | More than 80% at pH 5.2–11.3; and approximately 70% at pH 3.0, 4.2, and 12.3, after 1 h | [67] |
| Irpex lacteus MC-2 | Ex-4 | Pichia pastoris | PASC | - | - | - | - | 50 °C | pH 5.0 | More than 80% at 60 °C (pH 3.0–8.0) after 1 h | More than 80% at pH 3.0–8.0 (60 °C) after 1 h | [68] |
| Chaetomium thermophilum HSAUP072651 | CBH II | Pichia pastoris | pNPC | - | - | - | - | 50 °C | pH 4.0 | No loss at 50 °C; approximately 20% at 60 °C; and complete inactivation at 80 °C, after 1 h | - | [69] |
| Source Organism | Protein Name | Expression System | Substrate | Co-Substrate | Vmax | KM | kcat | kcat/KM | Optimal T | Optimal pH | Thermostability | pH Stability | Ref |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aspergillus fumigatus Af293 | AfAA9_B | Pichia pastoris X33 | 2,6-DMP | § a H2O2 | 78.52 ± 3.33 U g−1 | 2.04 ± 0.24 µM | 0.034 s−1 | 0.017 µM−1 s−1 | - | *** 9 | - | - | This study |
| § b H2O2 | 1481 ± 72.19 U g−1 | 0.79 ± 0.12 µM | 0.64 s−1 | 0.81 µM−1 s−1 | - | 60 °C: 50 % after 48 h 50 °C: almost 100% of activity after 48 h | No loss of activity at pH 5.0–10.0 | ||||||
| § 2,6-DMP | a H2O2 | 49.26 ± 4.48 U g−1 | 106.3 ± 27.9 µM | 0.021 s−1 | 1,98 × 10−4 µM−1 s−1 | - | - | - | - | ||||
| b H2O2 | 972.5 ± 28.31 U g−1 | 12.15 ± 1.76 µM | 0.42 s−1 | 0.035 µM−1 s−1 | - | - | - | - | |||||
| Scytalidium thermophilum | PMO9D_SCYTH | Pichia pastoris X33 | Avicel | § H2O2 | 0.36 U mg−1 | 4.54 mg mL−1 | 2.99 × 10−2 min−1 | 6.58 × 10−3 mg−1 mL min−1 | - | - | - | - | [70] |
| CMC | 14.96 U mg−1 | 10.6 mg mL−1 | 1.61 min−1 | 1.52 × 10−1 mg−1 mL min−1 | 60 °C | 7 | 60 °C (t1/2 = 60.58 h, pH 7.0) | Above 90% after 48 h at pH 7.0 | |||||
| 2,6-DMP | 0.13 U mg−1 | 0.51 mM | 1.84 × 10−1 min−1 | 3.57 × 10−1 mM−1 min−1 | - | - | - | ||||||
| Malbranchea cinnamomea | PMO9D_MALCI | Avicel | 0.17 U mg−1 | 5.87 mg mL−1 | 1.05 × 10−2 min−1 | 1.79 × 10−3 mg−1 mL min−1 | - | - | - | ||||
| CMC | 9.59 U mg−1 | 29.27 mg mL−1 | 0.76 min−1 | 2.62 × 10−2 mg−1 mL min−1 | 50 °C | 9 | 50 °C (t1/2 = 144 h, pH 7.0) | Above 80% after 72 h at pH 9.0 | |||||
| 2,6-DMP | 0.12 U mg−1 | 1.17 mM | 1.21 min−1 | 1.03 × 10−1 mM−1 min−1 | - | - | - | ||||||
| Thielavia terrestris | TtLPMO9E | - | PWS | § O2 | - | 49.80 g L−1 | 3.8 min−1 | * 1.85 × 103 M−1 min−1 | - | - | - | - | [71] |
| Myceliophthora thermophila | MtPMO9E | Neurospora crassa | cellohexaose | § O2 | - | 32 µM | 10.1 min−1 | 0.30 µM−1 min−1 | - | - | - | - | [72,73] |
| § cellohexaose | O2 | - | 230 µM | 17 min−1 | 7.4 × 10−2 µM−1 min−1 | - | - | - | - | ||||
| H2O2 | - | 53 µM | # 15.9 s−1 | 3.0 × 105 M−1 s−1 | - | - | - | - | |||||
| Serratia marcescens | CBP21 | Escherichia coli BL21(DE3) Star | CNW | § H2O2 | 1.11 µM s−1 | 0.58 mg mL−1 | 6.7 s−1 | ≅106 M−1 s−1 | - | - | - | - | [74] |
| § CNW | H2O2 | - | 2.8 µM | - | - | - | - | - | - | ||||
| Lentinus similis | Ls(AA9)A | Aspergillus oryzae MT3568 | cellotetraose | § O2 | - | 43 µM | 0.11 s−1 | 2.6 × 103 M−1 s−1 | - | - | - | - | [75] |
| Aspergillus fumigatus NITDGPKA3 | ** CAF32158.1 | Pichia pastoris X33 | 2,6-DMP | § H2O2 | 1.11 µM min−1 | 11.23 µM | 0.642 min−1 | 5.7 × 10−2 µM min−1 | - | - | - | - | [76] |
| Myceliophthora thermophila | MtLPMO9D | Myceliophthora thermophila C1 | - | - | - | - | - | - | - | - | **** Tmapp at pH 7.0 = 68 °C | - | [77] |
| Myceliophthora thermophila | MtLPMO9J | Aspergillus nidulans | - | - | - | - | - | - | - | - | **** Tmapp at pH 6.0 = 58 °C | - | [78] |
| Thermoascus aurantiacus | TaLPMO9A | Aspergillus oryzae | - | - | - | - | - | - | - | - | **** Tmapp at pH 7.0 = 69 °C | - | [79] |
| AfAA9_B | AfCel6A | |
|---|---|---|
| Additive | % Relative Activity | |
| None | 100.0 ± 1.0 | 100.0 ± 0.9 |
| SDS | 107.8 ± 4.8 | 53.72 ± 0.31 |
| Tween 20 | 103.7 ± 9.6 | 93.24 ± 2.11 |
| EDTA | 0 | 91.88 ± 1.56 |
| Ascorbic Acid | - | 121.40 ± 2.55 |
| DMSO | 108.3 ± 1.5 | 101.03 ± 3.58 |
| β-mercaptoethanol | 36.7 ± 0.6 | 134.24 ± 1.02 |
| ZnSO4 | 36.8 ± 6.1 | 83.67 ± 0.65 |
| MnCl2 | 82.2 ± 0.7 | 189.25 ± 2.33 |
| CoCl2 | 0 | 116.75 ± 1.36 |
| CaCl2 | 93.6 ± 3.3 | 101.1 ± 2.4 |
| FeSO4 | 0 | 125.83 ± 3.61 |
| MgSO4 | 113.9 ± 1.9 | 95.92 ± 2.13 |
| CuSO4 | 35.2 ± 4.6 | 85.08 ± 0.65 |
| AgNO3 | 0 | 179.27 ± 20.04 |
| KCl | 107.5 ± 2.1 | 98.87 ± 2.44 |
| (NH4)2SO4 | 88.9 ± 3.2 | 99.9 ± 2.1 |
| DTT | 0 | 150.68 ± 5.29 |
| Triton X-100 | 84.8 ± 2.3 | 91.55 ± 1.19 |
| SLS | 115.3 ± 0.7 | 93.80 ± 1.60 |
| Primer Name | Sequence (5′-3′) |
|---|---|
| AfAA9_B Fw | CAAAAAACAACTAATTATTCGAAACGAGGAATTCCATGACTTTGTCCAAGATCAC |
| AfAA9_B Rv | CAGATCCTCTTCTGAGATGAGTTTTTGTTCTAGAGCGTTGAACAGTGCAGGAC |
| AfCel6A Fw | GAGAAAAGAGAGGCTGAAGCTGAATTCCAGCAGACCGTATGG |
| AfCel6A Rv | ATCCTCTTCTGAGATGAGTTTTTGTTCTAGAAAGGACGGGTTAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardi, A.V.; Gerolamo, L.E.; de Gouvêa, P.F.; Yonamine, D.K.; Pereira, L.M.S.; de Oliveira, A.H.C.; Uyemura, S.A.; Dinamarco, T.M. LPMO AfAA9_B and Cellobiohydrolase AfCel6A from A. fumigatus Boost Enzymatic Saccharification Activity of Cellulase Cocktail. Int. J. Mol. Sci. 2021, 22, 276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010276
Bernardi AV, Gerolamo LE, de Gouvêa PF, Yonamine DK, Pereira LMS, de Oliveira AHC, Uyemura SA, Dinamarco TM. LPMO AfAA9_B and Cellobiohydrolase AfCel6A from A. fumigatus Boost Enzymatic Saccharification Activity of Cellulase Cocktail. International Journal of Molecular Sciences. 2021; 22(1):276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010276
Chicago/Turabian StyleBernardi, Aline Vianna, Luis Eduardo Gerolamo, Paula Fagundes de Gouvêa, Deborah Kimie Yonamine, Lucas Matheus Soares Pereira, Arthur Henrique Cavalcante de Oliveira, Sérgio Akira Uyemura, and Taisa Magnani Dinamarco. 2021. "LPMO AfAA9_B and Cellobiohydrolase AfCel6A from A. fumigatus Boost Enzymatic Saccharification Activity of Cellulase Cocktail" International Journal of Molecular Sciences 22, no. 1: 276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010276