Enzyme Properties of a Laccase Obtained from the Transcriptome of the Marine-Derived Fungus Stemphylium lucomagnoense

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Target Selection, Aspergillus Niger Transformation, and Screening
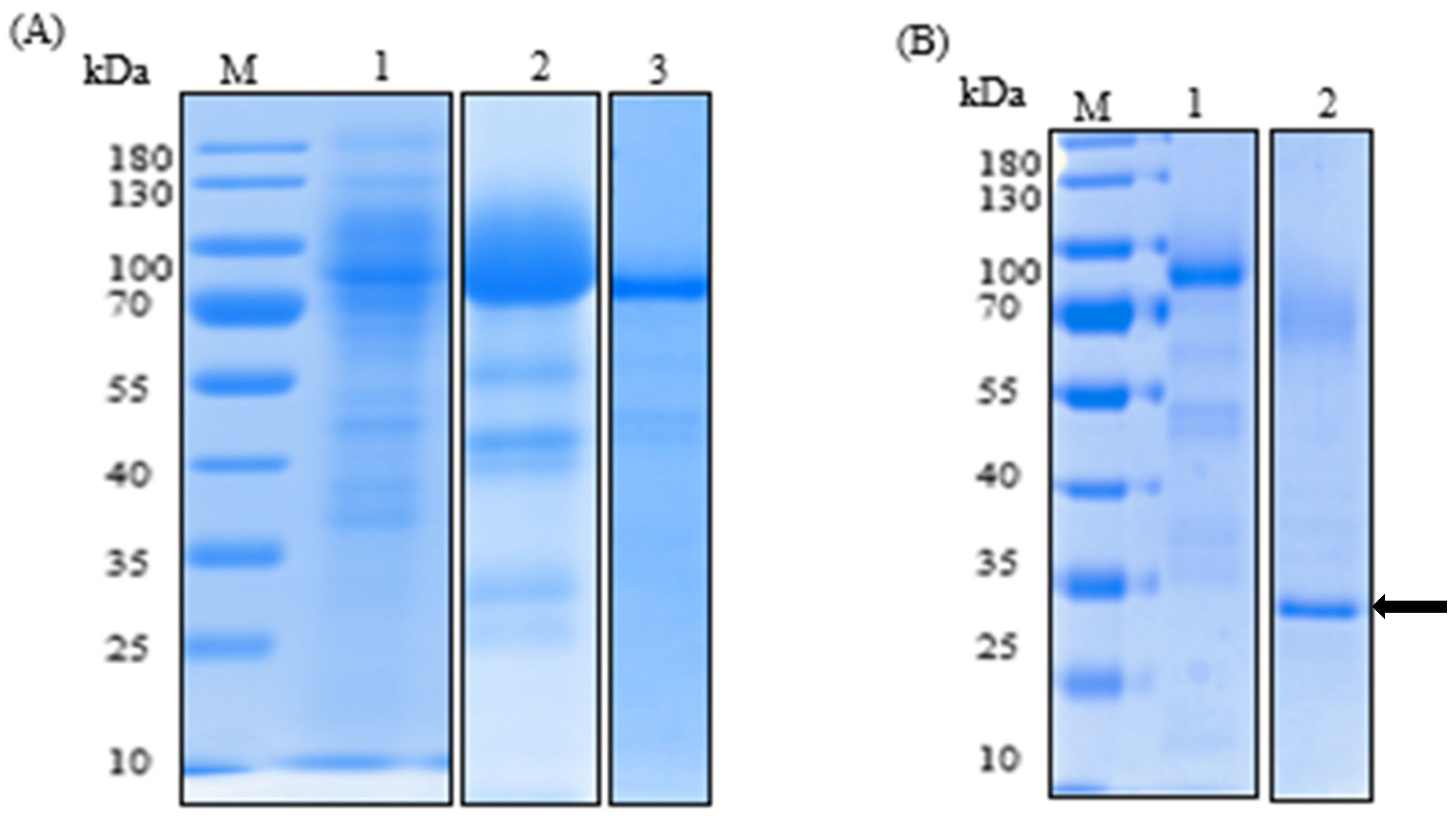
2.2. Purification of the Recombinant Laccase and Study of Physico-Chemical Properties
2.3. Kinetic Parameters
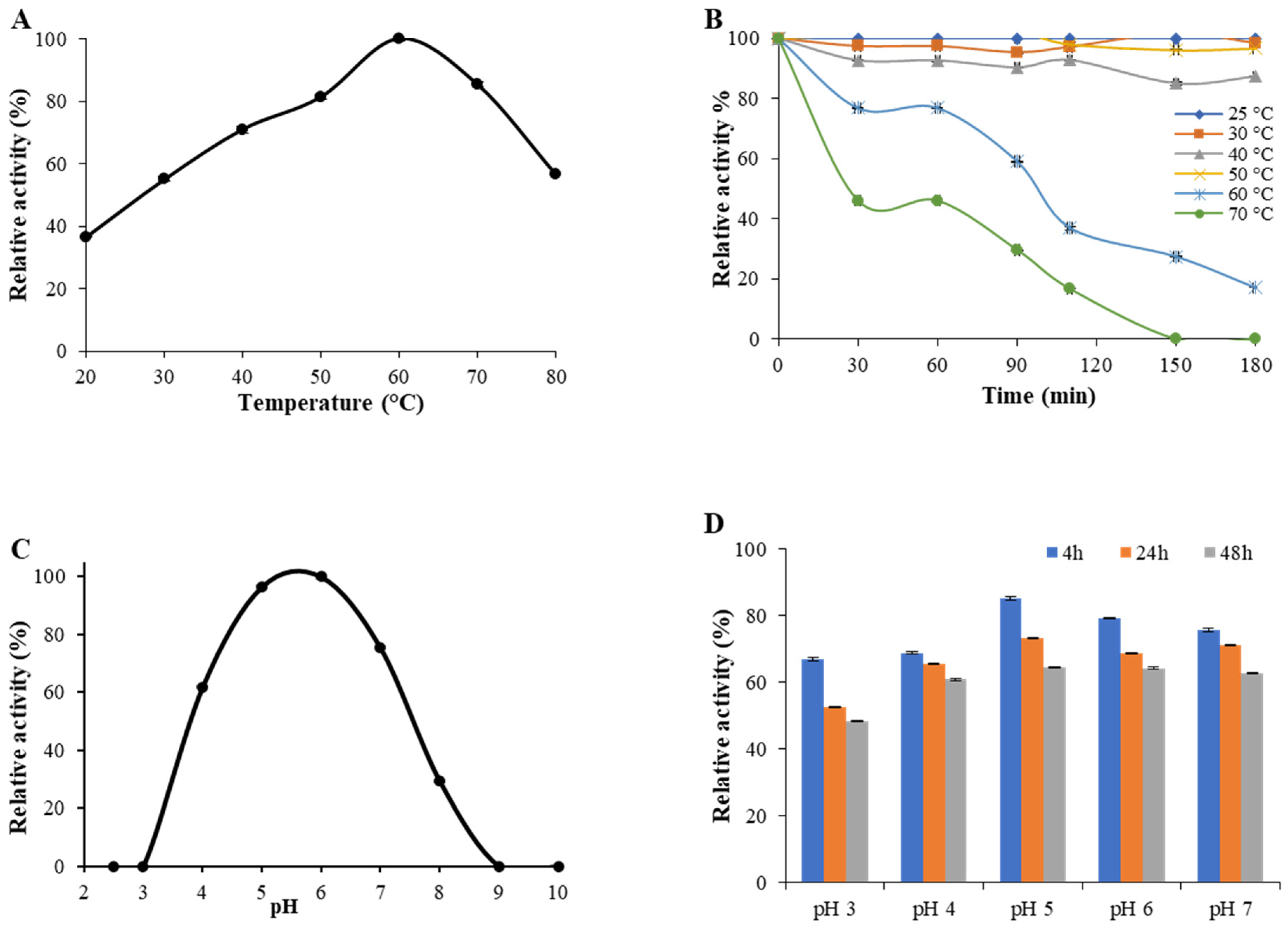
2.4. Enzyme Activity and Stability at Different pH and Temperature Values
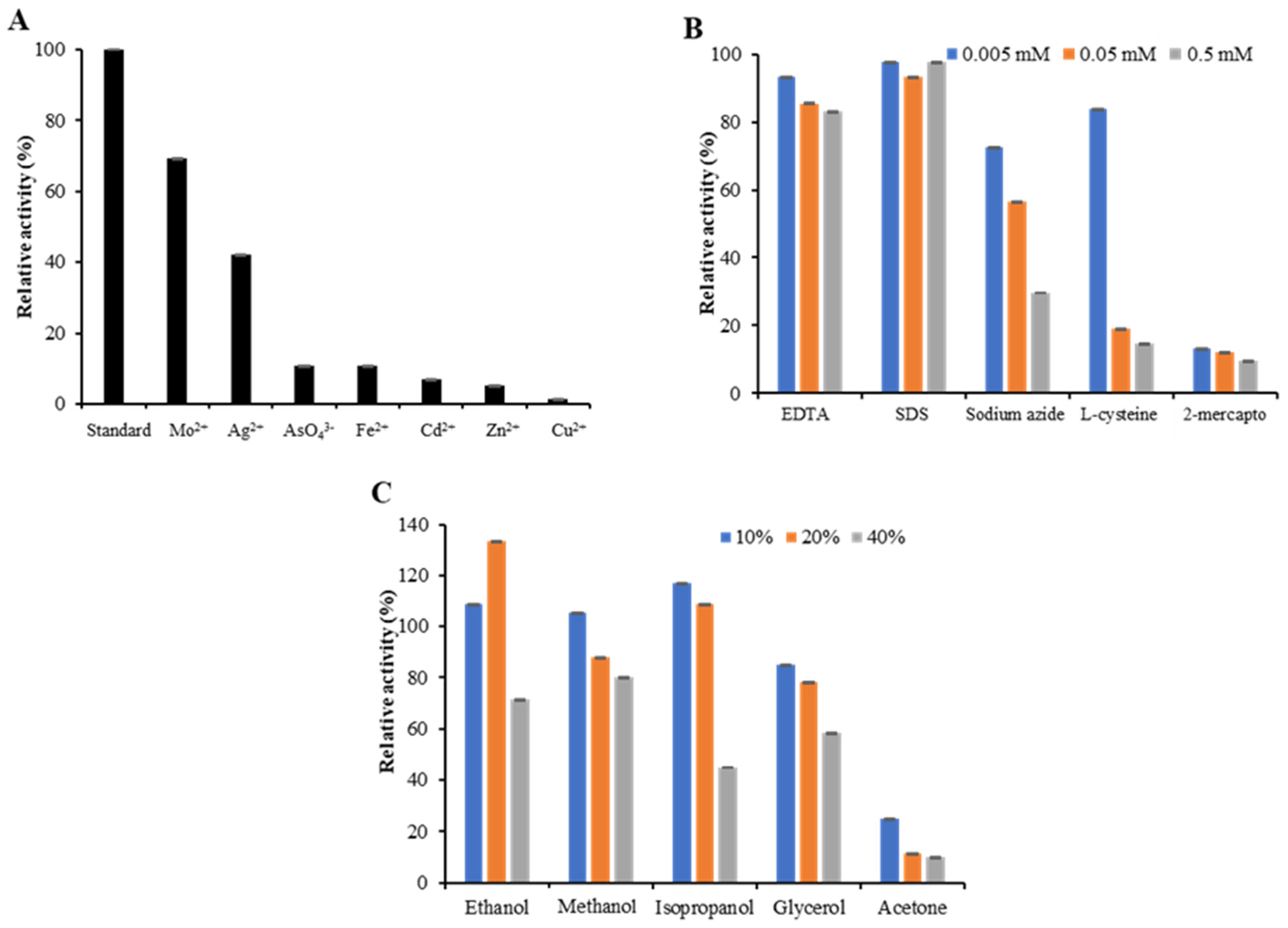
2.5. Effect of Metal Ions, Inhibitors and Solvents on Laccase Activity
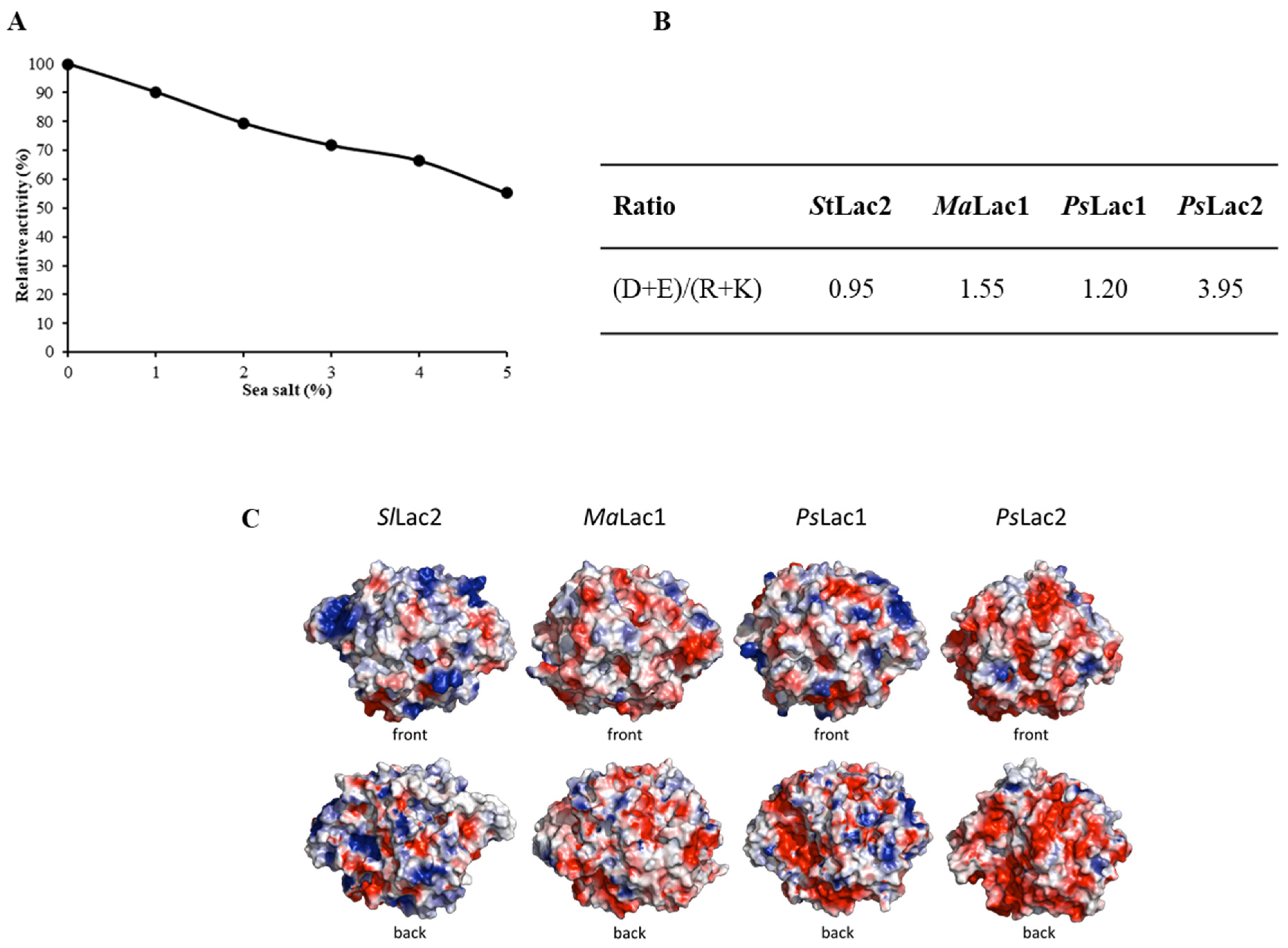
2.6. Effect of Sea Salt on Laccase Activity and Surface Charge of SlLac2
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Cloning and Expression of SlLac2-Encoding Gene
4.3. Screening of Transformants and Laccase Activity Assay
4.4. Production and Purification of Recombinant SlLac2
4.5. Bioinformatic Analysis
4.6. Determination of N-Terminal Amino Acid Sequence and Glycosylation Level
4.7. Substrate Specificity and Kinetics
4.8. Effect of pH and Temperature on the Activity and Stability of the Laccase
4.9. Effect of Metal Ions, Inhibitors, Solvents, and Sea Salt on Laccase Activity.
4.10. Nucleotide Sequence Accession Number
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sivakumar, R.; Rajendran, R.; Balakumar, C.; Tamilvendan, M. Isolation, screening and optimization of production medium for thermostable laccase production from Ganoderma sp. Int. J. Eng. Sci. Technol. 2010, 2, 7133–7141. [Google Scholar]
- Kumar, R.; Kaur, J.; Jain, S.; Kumar, A. Optimization of laccase production from Aspergillus flavus by design of experiment technique: Partial purification and characterization. J. Genet. Eng. Biotechnol. 2016, 14, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giardina, P.; Faroca, V.; Pezzela, P.C.; Piscitelli, A.; Vanhilla, S.; Sannia, G. Laccase: A never-ending story. Cell. Mol. Life Sci. 2010, 62, 369–386. [Google Scholar] [CrossRef] [PubMed]
- Durán, N.; Rosa, M.A.; D’Annibale, A.; Gianfreda, L. Applications of laccases and tyrosinases (phenoloxidases) immobilized on different supports: A review. Enzym. Microb. Technol. 2002, 31, 907–931. [Google Scholar] [CrossRef]
- Arora, D.S.; Sharma, R.K. Ligninolytic fungal laccases and their biotechnological applications. Appl. Biochem. Biotechnol. 2010, 160, 1760–1788. [Google Scholar] [CrossRef]
- Xu, F. Oxidation of phenols, anilines, and benzenethiols by fungal laccases: Correlation between activity and redox potentials as well as halide inhibition. Biochemistry 1996, 35, 7608–7614. [Google Scholar] [CrossRef]
- Minussi, R.C.; Pastore, G.M.; Durán, N. Enzima de interés en enología: Lacasas. Aliment 1999, 304, 145–150. [Google Scholar]
- Christopher, F. The structure and function of fungal laccase. Microbiology 1994, 140, 19. [Google Scholar]
- Baldrian, P. Purification and characterization of laccase from the white-rot fungus Daedalea quercina and decolorization of synthetic dyes by the enzyme. Appl. Microbiol. Biotechnol. 2004, 63, 560–563. [Google Scholar] [CrossRef]
- Li, K.; Xu, F.; Eriksson, K.E. Comparison of fungal laccases and redox mediators in oxidation of a nonphenolic lignin model compound. Appl. Microbiol. Biotechnol. 1999, 65, 2654–2660. [Google Scholar] [CrossRef] [Green Version]
- Munk, L.; Sitarz, A.K.; Kalyani, D.C.; Mikkelsen, J.D.; Meyer, A.S. Can laccases catalyze bond cleavage in lignin? Biotechnol. Adv. 2015, 33, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase Properties, Physiological Functions, and Evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Shin, W.; Brown, S.H.; Wahleithner, J.A.; Sundaram, U.M.; Solomon, E.I. A study of a series of recombinant fungal laccases and bilirubin oxidase that exhibit significant differences in redox potential, substrate specificity, and stability. Biochim. Biophys. Acta Protein Struct. Mol. Enzym. 1996, 1292, 303–311. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates: An expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Call, H.P.; Mücke, I. History, overview and applications of mediated lignolytic systems, especially laccase-mediator-systems (Lignozym®-process). J. Biotechnol. 1997, 53, 163–202. [Google Scholar] [CrossRef]
- Bajpai, P.; Kondo, R. Biotechnology for Environmental Protection in the Pulp and Paper Industry; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 3-642-60136-7. [Google Scholar]
- Wesenberg, D.; Kyriakides, I.; Agathos, S.N. White-rot fungi and their enzymes for the treatment of industrial dye effluents. Biotechnol. Adv. 2003, 22, 161–187. [Google Scholar] [CrossRef]
- Vinodhkumar, T.; Thiripurasundari, N.; Ramanathan, G.; Karthik, G. Screening of dye degrading bacteria from textile effluents. J. Chem. Pharm. Res. 2013, 3, 848–857. [Google Scholar]
- Michaels, G.B.; Lewis, D.L. Sorption and toxicity of azo and triphenylmethane dyes to aquatic microbial populations. Environ. Toxicol. Chem. 1985, 4, 45–50. [Google Scholar] [CrossRef]
- Chung, K.-T.; Stevens, S.E., Jr. Degradation azo dyes by environmental microorganisms and helminths. Environ. Toxicol. Chem. 1993, 12, 2121–2132. [Google Scholar]
- Ghoreishi, S.M.; Haghighi, R. Chemical catalytic reaction and biological oxidation for treatment of non-biodegradable textile effluent. Chem. Eng. J. 2003, 95, 163–169. [Google Scholar] [CrossRef]
- Santhanam, N.; Vivanco, J.M.; Decker, S.R.; Reardon, K.F. Expression of industrially relevant laccases: Prokaryotic style. Trends Biotechnol. 2011, 29, 480–489. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, M.; Zhang, M.; Wang, C.; Liu, Y.; Fan, X.; Li, H. Characterization of a novel, cold-adapted, and thermostable laccase-like enzyme with high tolerance for organic solvents and salt and potent dye decolorization ability, derived from a marine metagenomic library. Front. Microbiol. 2018, 9, 2998. [Google Scholar] [CrossRef] [Green Version]
- Bonugli-Santos, R.C.; Dos Santos Vasconcelos, M.R.; Passarini, M.R.Z.; Vieira, G.A.L.; Lopes, V.C.P.; Mainardi, P.H.; Dos Santos, J.A.; de Azevedo Duarte, L.; Otero, I.V.R.; da Silva Yoshida, A.M.; et al. Marine-derived fungi: Diversity of enzymes and biotechnological applications. Front. Microbiol. 2015, 6, 269. [Google Scholar] [CrossRef] [PubMed]
- Punt, P.J.; van den Hondel, C.A.M.J.J. Transformation of filamentous fungi based on hygromycin b and phleomycin resistance markers. Methods Enzymol. 1992, 216, 447–457. [Google Scholar]
- Ben Ali, W.; Chaduli, D.; Navarro, D.; Lechat, C.; Turbé-Doan, A.; Bertrand, E.; Faulds, C.B.; Sciara, G.; Lesage-Meessen, L.; Record, E.; et al. Screening of five marine-derived fungal strains for their potential to produce oxidases with laccase activities suitable for biotechnological applications. BMC Biotechnol. 2020, 20, 27. [Google Scholar] [CrossRef] [PubMed]
- Ben Ali, W.; Navarro, D.; Kumar, A.; Drula, E.; Turbé-Doan, A.; Correia, L.O.; Baumberger, S.; Bertrand, E.; Faulds, C.B.; Henrissat, B.; et al. Characterization of the CAZy Repertoire from the Marine-Derived Fungus Stemphylium lucomagnoense in Relation to Saline Conditions. Mar. Drugs 2020, 18, 461. [Google Scholar] [CrossRef]
- Wikee, S.; Hatton, J.; Turbé-Doan, A.; Mathieu, Y.; Daou, M.; Lomascolo, A.; Kumar, A.; Lumyong, S.; Sciara, G.; Faulds, C.B.; et al. Characterization and Dye Decolorization Potential of Two Laccases from the Marine-Derived Fungus Pestalotiopsis sp. Int. J. Mol. Sci. 2019, 20, 1864. [Google Scholar] [CrossRef] [Green Version]
- Atalla, M.M.; Zeinab, H.K.; Eman, R.H.; Amani, A.Y.; Abeer, A.A.E. Characterization and kinetic properties of the purified Trematosphaeria mangrovei laccase enzyme. Saudi J. Biol. Sci. 2013, 20, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Mainardi, P.H.; Feitosa, V.A.; de Paiva, L.B.B.; Bonugli-Santos, R.C.; Squina, F.M.; Pessoa, A., Jr.; Sette, L.D. Laccase production in bioreactor scale under saline condition by the marine-derived basidiomycete Peniophora sp. CBMAI 1063. Fungal Biol. 2018, 122, 302–309. [Google Scholar] [CrossRef]
- D’Souza-Ticlo, D.; Sharma, D.; Raghukumar, C. A Thermostable Metal-Tolerant Laccase with Bioremediation Potential from a Marine-Derived Fungus. Mar. Biotechnol. 2009, 11, 725–737. [Google Scholar] [CrossRef]
- Record, E.; Punt, P.J.; Chamkha, M.; Labat, M.; van den Hondel, C.A.; Asther, M. Expression of the Pycnoporus cinnabarinus laccase gene in Aspergillus niger and characterization of the recombinant enzyme. Eur. J. Biochem. 2002, 269, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P. Fungal laccases-occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef] [Green Version]
- Eggert, C.; Temp, U.; Eriksson, K.-E. The ligninolytic system of the white rot fungus Pycnoporus cinnabarinus: Purification and characterization of the laccase. Appl. Environ. Microbiol. 1996, 62, 1151–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, K.; Kang, F.; Waigi, M.G.; Gao, Y.; Huang, Q. Laccase-mediated transformation of triclosan in aqueous solution with metal cations and humic acid. Environ. Pollut. 2017, 220, 105–111. [Google Scholar] [CrossRef]
- Murugesan, K.; Kim, Y.-M.; Jeon, J.-R.; Chang, Y.-S. Effect of metal ions on reactive dye decolorization by laccase from Ganoderma lucidum. J. Hazard. Mater. 2009, 168, 523–529. [Google Scholar] [CrossRef]
- Lu, L.; Zhao, M.; Wang, T.-N.; Zhao, L.-Y.; Du, M.-H.; Li, T.-L.; Li, D.-B. Characterization and dye decolorization ability of an alkaline resistant and organic solvents tolerant laccase from Bacillus licheniformis LS04. Bioresour. Technol. 2012, 115, 35–40. [Google Scholar] [CrossRef]
- Si, J.; Peng, F.; Cui, B. Purification, biochemical characterization and dye decolorization capacity of an alkali-resistant and metal-tolerant laccase from Trametes pubescens. Bioresour. Technol. 2013, 128, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Siroosi, M.; Amoozegar, M.A.; Khajeh, K. Purification and characterization of an alkaline chloride-tolerant laccase from a halotolerant bacterium, Bacillus sp. strain WT. J. Mol. Catal. B Enzym. 2016, 134, 89–97. [Google Scholar] [CrossRef]
- Wang, S.-S.; Ning, Y.-J.; Wang, S.-N.; Zhang, J.; Zhang, G.-Q.; Chen, Q.-J. Purification, characterization, and cloning of an extracellular laccase with potent dye decolorizing ability from white rot fungus Cerrena unicolor GSM. Int. J. Biol. Macromol. 2017, 95, 920–927. [Google Scholar] [CrossRef]
- Yan, J.; Chen, D.; Yang, E.; Niu, J.; Chen, Y.; Chagan, I. Purification and characterization of a thermotolerant laccase isoform in Trametes trogii strain and its potential in dye decolorization. Int. Biodeter. Biodegr. 2014, 93, 186–194. [Google Scholar] [CrossRef]
- Younes, S.B.; Sayadi, S. Purification and characterization of a novel trimeric and thermotolerant laccase produced from the ascomycete Scytalidium thermophilum strain. J. Mol. Catal. B Enzym. 2011, 73, 35–42. [Google Scholar] [CrossRef]
- Xu, X.; Huang, X.; Liu, D.; Lin, J.; Ye, X.; Yang, J. Inhibition of metal ions on Cerrena sp. laccase: Kinetic, decolorization and fluorescence studies. J. Taiwan Inst. Chem. E 2018, 84, 1–10. [Google Scholar] [CrossRef]
- Bollag, J.-M.; Leonowicz, A. Comparative studies of extracellular fungal laccases. Appl. Environ. Microbiol. 1984, 48, 849–854. [Google Scholar] [CrossRef] [Green Version]
- Slomczynski, D.; Nakas, J.P.; Tanenbaum, S.W. Production and Characterization of Laccase from Botrytis cinerea 61. Appl. Environ. Microbiol. 1995, 61, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Johannes, C.; Majcherczyk, A. Laccase activity tests and laccase inhibitors. J. Biotechnol. 2000, 78, 193–199. [Google Scholar] [CrossRef]
- Milstein, O.; Hüttermann, A.; Majcherczyk, A.; Schulze, K.; Fründ, R.; Lüdemann, H.-D. Transformation of lignin-related compounds with laccase in organic solvents. J. Biotechnol. 1993, 30, 37–48. [Google Scholar] [CrossRef]
- Luterek, J.; Gianfreda, L.; Wojtaś-Wasilewska, M.; Cho, N.S.; Rogalski, J.; Jaszek, M.; Malarczyk, E.; Staszczak, M.; Fink-Boots, M.; Leonowicz, A. Activity of free and immobilized extracellular Cerrena unicolor laccase in water miscible organic solvents. Holzforsch. Int. J. Biol. Chem. Phys. Technol. Wood 1998, 52, 589–595. [Google Scholar]
- Robles, A.; Lucas, R.; Martínez-Cañamero, M.; Ben Omar, N.; Pérez, R.; Gálvez, A. Characterization of laccase activity produced by the hyphomycete Chalara (syn. Thielaviopsis) paradoxa CH. Enzym. Microb. Technol. 2002, 31, 516–522. [Google Scholar] [CrossRef]
- Patel, I.; Kracher, D.; Ma, S.; Garajova, S.; Haon, M.; Faulds, C.B.; Berrin, J.-G.; Ludwig, R.; Record, E. Salt-responsive lytic polysaccharide monooxygenases from the mangrove fungus Pestalotiopsis sp. NCi. Biotechnol. Biofuels 2016, 9, 108. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, S.; Xie, Y.; Fang, Z.; Xiao, Y.; Fang, W.; Zhang, X. Mechanism of the salt activation of laccase Lac. Biochem. Biophys. Res. Commun. 2020, 521, 997–1002. [Google Scholar] [CrossRef]
- Kern, M.; McGeehan, J.E.; Streeter, S.D.; Martin, R.N.A.; Besser, K.; Elias, L.; Eborall, W.; Malyon, G.P.; Payne, C.M.; Himmel, M.E.; et al. Structural characterization of a unique marine animal family 7 cellobiohydrolase suggests a mechanism of cellulase salt tolerance. Proc. Natl. Acad. Sci. USA 2013, 110, 10189–10194. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Bag, S.K.; Das, S.; Harvill, E.T.; Dutta, C. Molecular signature of hypersaline adaptation: Insights from genome and proteome composition of halophilic prokaryotes. Genome Biol. 2008, 9, R70. [Google Scholar] [CrossRef] [Green Version]
- Lanyi, J.K. Salt-dependent properties of proteins from extremely halophilic bacteria. Bacteriol. Rev. 1974, 38, 272. [Google Scholar] [CrossRef] [Green Version]
- Gordon, C.L.; Khalaj, V.; Ram, A.F.; Archer, D.B.; Brookman, J.L.; Trinci, A.P.; Jeenes, D.J.; Doonan, J.H.; Wells, B.; Punt, P.J. Glucoamylase: Green fluorescent protein fusions to monitor protein secretion in Aspergillus niger. Microbiology 2000, 146, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Van Hartingsveldt, W.; Mattern, I.E.; van Zeijl, C.M.; Pouwels, P.H.; van den Hondel, C.A. Development of a homologous transformation system for Aspergillus niger based on the pyrG gene. Mol. Gen. Genet. MGG 1987, 206, 71–75. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Chojnacki, S.; Cowley, A.; Lee, J.; Foix, A.; Lopez, R. Programmatic access to bioinformatics tools from EMBL-EBI update: 2017. Nucleic Acids Res. 2017, 45, W550–W553. [Google Scholar] [CrossRef] [Green Version]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.-B.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Signal Peptide (Amino Acids) | Mature Protein (Amino Acids) | Molecular Mass (Da) | PI | Coordination Sites |
|---|---|---|---|---|---|
| Q70KY3 | 22 | 601 | 68958 | 5.18 | IHWHG WYHSH HPMHLH HCHIAWH |
| KY554800 | 21 | 555 | 60904 | 6.09 | IHWHG WYHSH HPMHLH HCHIAWH |
| KY554801 | 18 | 543 | 59813 | 4.10 | IHWHG WYHSH HPMHLH HCHIAWH |
| APMZ2_prot14771 * | 21 | 592 | 64581 | 8.07 | IHWHG WYHSH HPIHLH HCHIAFH |
| APMZ2_prot8345 | 23 | 578 | 63456 | 5.64 | IHWHG WYHSH HPIHLH HCHIAWH |
| APMZ2_prot7177 | 21 | 615 | 68221 | 6.03 | IHFHG WYHSH HPIHKH HCHINNH |
| APMZ2_prot10330 | No | 754 | 83453 | 5.40 | IHFHG WYHAH HPFHLH HCHNMWH |
| APMZ2_prot15523 | No | 729 | 80156 | 5.37 | LHAHG WYHSH HPMHLH HCHLAWH |
| APMZ2_prot2393 | No | 554 | 60438 | 4.66 | LHFHG WYHSH HPFHLH HCHIEWH |
| APMZ2_prot9198 | 19 | 576 | 64532 | 5.08 | MHWHG FYHSH HPFHLH HCHVLQH |
| Purification Steps | Volume (mL) | Total Activity (nKat) | Protein (mg) | Specific Activity (nkat mg−1) | Activity Yield (%) | Purification Factor (fold) |
|---|---|---|---|---|---|---|
| Culture medium | 930 | 78,027 | 176 | 443 | 100 | 1 |
| Ultrafiltration | 500 | 99,000 | 156 | 635 | 126.9 | 1.4 |
| CM-Sepharose | 325 | 20,775 | 15.4 | 1349 | 26.6 | 3.0 |
| Sephacryl S-200HR | 40 | 18,782 | 7.2 | 2609 | 24.1 | 5.9 |
| Substrate | KM (mM) | Kcat (s−1) | Kcat/KM (s−1 mM−1) | |
|---|---|---|---|---|
| ABTS |  | 0.0206 +/− 0.0039 | 0.153 +/− 0.0026 | 7.42 |
| DMP |  | 0.0240 +/− 0.0039 | 0.0059 +/− 0.0003 | 0.24 |
| Syringaldazine |  | 0.0035 +/− 0.00057 | 0.0074 +/− 0.0002 | 2.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, W.B.; Ayed, A.B.; Turbé-Doan, A.; Bertrand, E.; Mathieu, Y.; Faulds, C.B.; Lomascolo, A.; Sciara, G.; Record, E.; Mechichi, T. Enzyme Properties of a Laccase Obtained from the Transcriptome of the Marine-Derived Fungus Stemphylium lucomagnoense. Int. J. Mol. Sci. 2020, 21, 8402. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218402
Ali WB, Ayed AB, Turbé-Doan A, Bertrand E, Mathieu Y, Faulds CB, Lomascolo A, Sciara G, Record E, Mechichi T. Enzyme Properties of a Laccase Obtained from the Transcriptome of the Marine-Derived Fungus Stemphylium lucomagnoense. International Journal of Molecular Sciences. 2020; 21(21):8402. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218402
Chicago/Turabian StyleAli, Wissal Ben, Amal Ben Ayed, Annick Turbé-Doan, Emmanuel Bertrand, Yann Mathieu, Craig B. Faulds, Anne Lomascolo, Giuliano Sciara, Eric Record, and Tahar Mechichi. 2020. "Enzyme Properties of a Laccase Obtained from the Transcriptome of the Marine-Derived Fungus Stemphylium lucomagnoense" International Journal of Molecular Sciences 21, no. 21: 8402. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218402